
Abstract
Lysosomal storage diseases are a class of inborn errors of metabolism that lead to widespread disease in multiple tissues. The murine model of mucopolysaccharidosis type VII (MPS VII) closely parallels the human syndrome and has been extensively used to investigate the natural history and therapeutic strategies for lysosomal storage diseases in general. Here we demonstrate a previously undescribed immune defect in the MPS VII mouse. Although the normal populations of cells are present in lymph nodes of these mice, MPS VII mice show a blunted T cell proliferative response and decreased antibody production after immunization with antigens. One mechanism of this defect is ineffective processing of protein antigens, as responses to peptide antigens are normal. This phenotype is presumably caused by the lysosomal disorder, as the defect can be corrected in vivo by direct enzyme replacement therapy. These findings have implications for the use of this animal model, and may have clinical significance for other, more-common lysosomal storage diseases.
Similar content being viewed by others

Main
MPS VII is a lysosomal storage disease caused by the absence of the lysosomal enzyme GUSB (1). This defect results in a progressive accumulation of undegraded GAGs, resulting in widespread lysosomal distention. Virtually all organ systems are affected, leading to a constellation of symptoms, which include abnormal skeletal development, visceral lesions, mental retardation, hearing defects, and early mortality (2–4). Like most of the lysosomal storage diseases, MPS VII is inherited as an autosomal recessive trait, and several different human mutations have been described (5). The murine model of MPS VII closely reproduces many of the characteristics of the human disease and has provided much information on the pathogenesis of the mucopolysaccharidoses. In this model, a single base pair deletion in exon 10 of the GUSB gene leads to a frameshift mutation and premature stop codon, resulting in the absence of GUSB protein and mRNA in all tissues. Although MPS VII is rare in humans, the similarity of symptoms for many of the lysosomal storage diseases, coupled with the limited number of small animal models, have made the MPS VII mouse an important tool for examining lysosomal storage diseases in general. The murine MPS VII model has been widely used to examine the therapeutic effects of treatments such as ERT (6–8), bone marrow transplantation (2, 9, 10), and gene therapy (11–15).
The lysosomal compartment plays an important role in the assembly of MHC class II molecules with processed protein antigens. After production in the endoplasmic reticulum, empty MHC class II molecules associated with invariant chain move into the endosomal pathway. The removal of invariant chain from nascent MHC class II molecules and subsequent binding of antigenic fragments occurs mainly in a dense set of organelles with lysosomal properties (16). Disruption of the lysosomal pathway using agents such as chloroquine inhibits the ability of macrophages to present antigen via MHC class II (17–19).
In the MPS VII mouse, cells of the macrophage lineage are among the most severely affected cell types in the body (20). Grossly distended lysosomes and distortion of cellular architecture are found in Kupffer cells of the liver, peritoneal macrophages, and reticuloendothelial cells of the spleen. Although this histopathologic phenotype has long been observed, little is known about the effects of lysosomal storage disease on immune function in vivo. For some of the lysosomal storage diseases, recurrent infections have been described as a cause of morbidity (21, 22). In addition, a minor defect in lymphocyte function has been described in patients with Gaucher's disease (23).
Here we describe the first detailed report of an immune defect in an animal model of lysosomal storage disease. MPS VII mice have a deficient response to antigen priming by multiple routes. The mechanism of this defect appears to be caused by an inability to process protein antigens efficiently, as the defect can be bypassed by using peptide antigens to prime the animal. Reconstitution of the missing lysosomal enzyme restores immune function to normal. These results may have implications for the management of patients with lysosomal storage diseases and are important for the interpretation of data generated in this mouse model.
METHODS
Mice.
Syngeneic B6.C-H-2bm1 (B6) and MPS VII (mps/mps) mice were obtained from a pedigreed colony of B6.C-H-2bm1/ByBir-gusmps/+ mice maintained by strict brother-sister matings by M.S.S. at Washington University. Mice were housed in a specific pathogen-free facility in microisolation cages under a strictly controlled light cycle and given a high-fat autoclaved chow diet ad libitum. All mice were maintained in accordance with the guidelines of the Animal Studies Committee of Washington University.
Flow cytometry.
Single-cell suspensions were prepared by mechanical disruption in Hanks' balanced salt solution (HBSS; BioWhittaker, Inc., Walkersville, MD, U.S.A.) supplemented with 1% newborn calf serum (HyClone, Logan, UT, U.S.A.), 1 mM HEPES, 50 U/mL penicillin, and 50 μg/mL streptomycin (BioWhittaker). The cell suspension was passed through a 40-μm Nytex filter to remove debris. All cells were resuspended in FACS buffer [HBSS with 1% BSA and 1 mg/mL human IgG (Sandoz Pharmaceuticals, East Hanover, NJ, U.S.A.)] at 2 × 107 cells/mL. Primary antibody was added to 50 μL of each cell suspension (1 μg/106 cells) and allowed to incubate for 30 min on ice. The following antibodies were purchased from Pharmingen (San Diego, CA, U.S.A.): hamster anti-mouse CD3-ε (no. 01081D), rat anti-mouse CD4 (no. 01064A), rat anti-mouse CD45 (no. 01114A), hamster anti-mouse TCRαβ (no. 01302A), and goat anti-mouse Igs (no. 12062D). Rat anti-mouse CD8 (no. RM2215) was purchased from Caltag (Burlingame, CA, U.S.A.). After one 4-mL wash with FACS buffer, the secondary antibodies were added in 100 μL of FACS buffer and allowed to incubate in the dark for 30 min on ice. After a final wash, the cells were resuspended in 400 μL of FACS buffer and immediately analyzed, or fixed with 1% paraformaldehyde for analysis at a later date. Flow cytometric analysis was done on a triple-laser flow cytometer (FACSCAN, Becton-Dickinson, Mountain View, CA, U.S.A.) and analysis was performed on a Macintosh Quadra 650 using the Cell Quest program (Becton-Dickinson). Dead cells were excluded on the basis of forward and side light scatter, and the cells present in the lymphocyte gate as defined by light scatter and CD45+ staining were analyzed.
CD3-ε stimulation assay.
Splenocytes from 9-wk-old +/+ or MPS VII mice were incubated on either immobilized hamster anti-mouse CD3-ε or control antibody (hamster anti-TNP, Clone A19–3, Pharmingen) in a 96-well tissue culture plate (Costar 3799, Cambridge, MA, U.S.A.) in RPMI media (BioWhittaker) containing 0.5% murine serum, 2 mM Glutamax I (Life Technologies, Gaithersburg, MD, U.S.A.), 50 U/mL penicillin, 50 μg/mL streptomycin, and 2 × 10−5 M β-mercaptoethanol (2-ME, Fisher Scientific, Pittsburgh, PA, U.S.A.). The assay plate was incubated for 72 h at 37°C, 5% CO2, and then pulsed with 1 μCi [3H]thymidine (New England Nuclear, Boston, MA, U.S.A.) for 18–24 h. Assays were harvested on a Micro 96 harvester (Skatron Instruments, Tranby, Norway), and incorporation of [3H]thymidine was determined using a Betaplate liquid scintillation counter (Wallac, Inc., Gaithersburg, MD, U.S.A.).
LN proliferation assays.
Age-matched groups of 3–5 +/+ and mps/mps mice, ranging from 6 to 12 wk of age, received footpad injections of antigen emulsified in CFA as previously described (24). Briefly, mice were primed in each of the rear footpads with 20 μg (for protein antigens) or 20 μmol (for peptide antigens) emulsified in CFA (H37Ra, Adjuvant Complete, Difco Laboratories, Inc., Detroit, MI, U.S.A.). Twelve to 14 d later, the popliteal LNs were removed and pooled, and a single-cell suspension was prepared. The cells were cultured in 96-well microtiter plates at a final concentration of 5 × 105 cells per 0.2 mL. The culture medium consisted of RPMI 1640 supplemented with 2 mM Glutamax I, 1 mM HEPES, 1 mM sodium pyruvate (BioWhittaker), 50 U/mL penicillin, 50 μg/mL streptomycin, 2 × 10−5 M β-mercaptoethanol, and 0.5% (vol/vol) of freshly obtained autologous normal mouse serum. To avoid uptake of corrective GUSB from the serum, mps/mps serum was used for both +/+ and mps/mps LNs. The cultures were incubated at 37°C, 5% CO2, for 72 h, with each well being pulsed with 1.0 μCi of [3H]thymidine for the last 18 h of culture. Each group contained a positive control that consisted of wells containing purified protein derivative, which was generated as the soluble portion of a sonicated preparation of H37Ra organisms. All data are shown as a change in counts per minute, with subtraction of the background proliferation from cultures containing media alone. HEL and OVA were obtained from Sigma Chemical Co. (St. Louis, MO, U.S.A.) and used as intact proteins. The immune response to these proteins has been well characterized in the B6 mouse background (25). HEL peptide 74–88 was synthesized by Genosys Biotechnologies (The Woodlands, TX, U.S.A.).
Intraperitoneal antibody stimulation and ELISA assay.
On day 0, mice were primed intraperitoneally with 20 μg of protein antigen emulsified in CFA. On day 7, mice received an i.p. boost using 20 μg of protein antigen emulsified in IFA. Blood samples were obtained before the first injection and at day 7 by retroorbital eye bleed. On day 14, mice were killed, and blood was collected by cardiac bleed. Detection of HEL-specific antibodies in serum was accomplished by a standard ELISA, using 0.5 μg of HEL protein coated on Immulon 4 microtiter plates (Dynex Technologies, Chantilly, VA, U.S.A.). Briefly, HEL diluted in PBS was coated onto the microplates by an overnight incubation at 4°C. Excess antigen was removed by five washes in PBS with 0.05% Tween-20 (Sigma Chemical Co.) (wash buffer), and unbound sites were blocked with 5% BSA in wash buffer. Serum samples were diluted in 1% BSA in wash buffer (diluent buffer), added to each well and incubated at room temperature for 2 h or overnight at 4°C. After five washes, the appropriate alkaline phosphatase-conjugated secondary antibodies at a 1:2000 dilution (goat anti-mouse IgG, no. 1030–04, goat anti-mouse IgM, no. 1020–04, Southern Biotechnology Associates, Birmingham, AL, U.S.A.) were added to each well, and samples were incubated 2 h at room temperature. After a final set of washes, each well was incubated in p-nitrophenyl phosphate substrate (Sigmafast tablets, Sigma Chemical Co.). The reaction was stopped with 3 N sodium hydroxide, and the OD was read at 405 nm. All results shown are those obtained at a 1:100 dilution of sera.
ERT.
Recombinant human GUSB (26) was kindly provided by W. S. Sly. Adult mice received injections of 29,000 U of purified recombinant human GUSB via tail vein injection on the following schedule: the first injection of enzyme was given 8 d before footpad priming, with a second injection of enzyme given 1 d before footpad priming. A third injection of enzyme was given 7 d after footpad treatment. Control mice received injections of PBS on the same schedule. Mice were killed 14 d after footpad priming, and lymphocyte proliferation assays were performed as described above.
Statistical analysis.
A standard t test was performed using Graph Pad Prism (San Diego, CA, U.S.A.) for data analysis.
Results
MPS VII mice have a defective proliferative response to footpadinoculation.
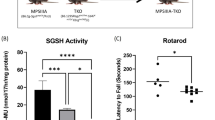
Nine-week-old MPS VII mice received a single injection in the rear footpads with a mixture of CFA and 20 μg of OVA. Age-matched normal mice and mice heterozygous for the mps mutation were injected as controls. Animals were killed 14 d after injection, and popliteal LNs were harvested. Lymphocytes from homozygous normal and heterozygous mice showed dose-dependent proliferative responses when exposed to OVA in vitro (Fig. 1). In contrast, MPS VII mice showed a severely blunted response to antigen, with a >3-log difference seen in proliferative responses to protein antigens. Similar results were seen when HEL was used as the antigen (data not shown). Because MPS VII is a progressive disease, we performed additional experiments using mice primed by footpad injection at either 6 or 12 wk of age. Regardless of the age at which the mice were primed, MPS VII mice consistently showed a severely reduced response (data not shown).

Proliferative response after footpad inoculation with protein antigen. Nine-week-old B6 (+/+), MPS VII (mps/mps), and heterozygous (+/mps) mice were primed in the rear footpads with 20 μg of OVA emulsified in CFA, and killed 14 d later. Popliteal LNs were harvested and assayed for OVA-specific proliferative activity. Lymphocytes from both normal and heterozygous mice showed a strong proliferative response to antigen in vitro. In contrast, lymphocytes from MPS VII mice showed a greatly reduced proliferative response. Assays were performed in triplicate. Bars represent ±1 SD. Each point represents the average of three mice.
MPS VII mice show no quantitative or qualitative differences inlymphocyte populations.
To determine whether the defective response was because of the lack of a given lymphocyte subset in MPS VII mice, FACS analysis was performed on lymphocytes obtained from multiple organs in normal and MPS VII mice. Although some minor shifts in lymphocyte populations were seen, no populations were notably absent in the thymus, spleen, or LNs of MPS VII mice when compared with age-matched normal controls (Table 1). In addition, the cellular composition of popliteal LNs 12 d after footpad stimulation with OVA protein was similar in MPS VII mice and normal controls, even though the proliferative responses are significantly different (Table 1), indicating that lymphocyte trafficking to sites of inflammation is grossly intact.
To determine whether the lack of an in vivo response was because of an intrinsic proliferative defect in MPS VII lymphocytes, we examined the response of splenocytes to α-CD3 antibody stimulation in vitro. Both normal and mutant mice showed a dose-dependent response to α-CD3 antibody stimulation in vitro, suggesting that the defect seen in MPS VII mice is not primarily because of a T cell receptor-mediated functional defect (Fig. 2). In addition, responses to concanavalin A-mediated stimulation were similar for both mutants and normals (data not shown).

α-CD3 stimulation of naive splenocytes. Splenocytes were isolated from naive 9-wk-old B6 (+/+) and MPS VII (mps/mps) mice and assayed for in vitro response to αCD3 antibody. Both groups of mice showed equivalent, dose-dependent proliferative responses over a range of antibody dilutions. Each point represents the average of three mice. Bars indicate ±1 SD.
MPS VII mice respond normally to peptide, but not protein,antigens.
Because the most severely affected cells in the MPS VII mouse are those of the macrophage lineage, we hypothesized that the defect might be caused by impaired antigen processing by dendritic cells or macrophages after s.c. immunization. To test this, LN stimulation was performed as previously described using either intact HEL protein or HEL peptide 74–88. This peptide has previously been shown to be the immunodominant peptide in the C57BL/6 strain (25, 27). Once again, MPS VII mice had a very poor in vitro response to either protein or peptide when primed with intact HEL protein in vivo (Fig. 3A). However, when mice were initially primed with HEL peptide 74–88, popliteal LN lymphocytes from MPS VII mice showed an in vitro response to peptide antigen that approached that seen in normal mice (Fig. 3B). Neither group of mice responded well to protein antigen in vitro after peptide priming. This phenomenon of poor priming to intact protein after immunization with a single peptide has been previously described (28).

Proliferative response after footpad inoculation with peptide antigen. Mice were primed with either intact HEL protein or HEL peptide 74–88. Although lymphocytes from protein-primed B6 mice (+/+) responded to both HEL protein and peptide in vitro, lymphocytes from protein-primed MPS VII mice (mps/mps) showed no response (A). In contrast, MPS VII mice primed with HEL peptide 74–88 showed a much improved response to peptide in vitro, which was comparable to that seen in age-matched normals (B). Neither group of peptide-primed mice responded well to protein antigen. Each antigen concentration was tested in triplicate. Each point represents the average of three mice. Bars equal ±1 SD.
The immune defect can be corrected using ERT.
To determine whether the immune defect in the MPS VII mouse could be attributed to the lysosomal storage disease, adult mice were treated with ERT before footpad injection. Six-week-old MPS VII mice were treated with 29,000 U of human enzyme via tail vein injection. Similar amounts of enzyme have been shown in previous studies to transiently reduce lysosomal distention in the liver and spleen of MPS VII mice (6). MPS VII mice received ERT on day −8 and −1 before footpad injection. Control groups included normal and MPS VII mice who received i.v. injections of PBS on the same schedule. After footpad injection with HEL protein in CFA on day 0, mice received an additional treatment with ERT or PBS on day 7. Animals were killed on day 14, and in vitro proliferation assays were performed. ERT-treated mice showed a response to in vitro stimulation with HEL protein that was significantly better than that seen in untreated MPS VII mice (Fig. 4).

Proliferative response after ERT. MPS VII mice (mps/mps-ERT) received ERT with GUSB as described in Methods. Control groups of age-matched B6 mice (+/+) and MPS VII mice (mps/mps) received PBS injections on the same schedule. All mice were primed with 20 μg of HEL emulsified in CFA in the rear footpads at 9 wk of age and killed 14 d later. Lymphocytes from MPS VII mice treated with GUSB showed an improved proliferative response to HEL protein in vitro when compared with untreated mutants. This response approached that seen in normal mice. Each antigen concentration was tested in triplicate. Each point represents the average of three mice. Bars represent ±1 SD. Data were analyzed by t test. * indicates a significant difference (p < 0.05) when compared with untreated MPS VII mice.
MPS VII mice also have defective response to intraperitonealstimulation.
To ensure that the observed immunologic defect was not caused by an anatomic anomaly specific to the footpads of MPS VII mice (such as poor lymphatic drainage secondary to bone and joint abnormalities), we assayed the ability of MPS VII mice to produce antibody after intraperitoneal stimulation. Nine-week-old mice received i.p. injections of HEL protein in CFA, and were subsequently boosted intraperitoneally with HEL protein in IFA 12 d after the initial treatment. Serum was collected at day 0, 12, and 19 and assayed for the presence of HEL-specific IgM and IgG. Before treatment, amounts of antigen-specific immunoglobulin were low for all animals. However, by 12 d after treatment, a clear difference could be seen in amounts of HEL-specific IgM between normal mice and their MPS VII siblings (Fig. 5). This difference was also seen in the secondary response, where IgG concentrations were much higher in normal mice compared with MPS VII mice. Given the normal B cell population observed by flow cytometry in these mice (Table 1) and the defective T cell proliferation described in this paper, we believe that the defective antibody response is because of a lack of T cell help.

Antibody production after intraperitoneal injection. Nine-week-old mice were primed i.p. with 20 μg HEL protein in CFA, and boosted 7 d later with 20 μg HEL protein in IFA. Blood was taken before the first injection (prebleed), before the second injection on day 12 (primary), and at time of sacrifice on day 19 (secondary). Serum was assayed for HEL-specific antibodies by ELISA. All results are shown at 1:100 dilution of serum to present results in the linear range of the assay. Each bar represents serum from an individual mouse. Although preimmunization amounts of antibody are similar in normals (solid bars) and mutants (hatched bars), MPS VII mice show a decreased IgM response at both primary and secondary time points (A). This effect is more pronounced with the secondary IgG response, in which concentrations in MPS VII mice at the secondary time point are virtually absent (B).
Discussion
MPS VII is a systemic, multiorgan disease caused by the absence of GUSB. Here we demonstrate that mice with this disease have a blunted immune response to stimulation by both footpad and intraperitoneal inoculation of antigen. This dysfunction correlates well with the histopathologic pattern of disease seen in these animals. Although lysosomal distension is seen in many tissues, accumulation of undegraded GAGs is most pronounced in cells of the reticuloendothelial system, such as Kupffer cells, peritoneal macrophages, and splenic sinus lining cells (20). The importance of this cell lineage in antigen processing suggests a potential cause for the immunologic defect observed in MPS VII mice.
The observation that MPS VII mice respond to peptide antigens more strongly than protein antigens indicates that the mechanism of this defect may lie in antigen processing. Although we did not directly assay for MHC class II on the surface of mutant cells by flow cytometry, an in vitro proliferative response to peptide antigens was demonstrated, indicating the presence of functional class II molecules. Peptide antigens have the capacity to bind the MHC molecule directly, bypassing the need for lysosomal processing required by intact protein antigens. On the basis of knowledge of the role of the lysosome in MHC assembly and antigen presentation, some possible causes for this antigen-processing defect can be suggested. First, the accumulation of undegraded, negatively charged GAGs may alter the lysosomal environment in a manner that broadly inhibits lysosomal function. It has been shown that the addition of GAGs such as chondroitin sulfate to in vitro enzyme reactions inhibits the hydrolytic activity of multiple lysosomal enzymes (29, 30). This generalized inhibition of lysosomal function has also been seen in vivo in human patients with lysosomal storage disease, in which the absence of a single lysosomal enzyme results in the accumulation of substrates from multiple different enzymes (29, 31). Alternatively, the accumulation of undegraded GAGs may interfere with MHC peptide assembly strictly on a physical basis by reducing the probability that peptides and MHC molecules will come into contact.
Although the MPS VII mouse has not traditionally been associated with an immune deficiency, there have been some suggestions of an altered immune response in this animal model. Wolfe et al. (32) reported an increase in the toxicity of both wild-type herpes simplex virus and recombinant herpes simplex virus–based vectors after ocular administration to MPS VII mice. Another observation is that the generation of antibodies against human GUSB commonly used in ERT studies has been surprisingly limited (6, 7). Finally, although generation of B cell hybridomas from untreated MPS VII mice is very difficult, pretreatment with human GUSB before immunization greatly enhances the number of hybridomas generated. (W. S. Sly, St. Louis University School of Medicine, 1998, personal communication). The presence of an immune defect in this widely used animal model mandates caution when evaluating responses against the therapeutic enzyme, as well as when analyzing the innate immunogenicity of viral vectors used for gene transfer. A lack of an inflammatory response to a vector used in the MPS VII mouse may actually be a characteristic of the MPS VII immune defect, rather than a function of the vector used.
Because of the relative scarcity of patients with MPS VII, it is unclear whether a similar defect is present in humans. However, recurrent upper respiratory infections, pneumonia, bronchitis, and middle ear infections have all been described in human MPS VII patients with severe disease (21, 33). Unfortunately, no detailed studies of the immune systems in MPS VII patients have been published, and large-scale studies of morbidity and mortality in this disease are not available. Therefore, it is unclear whether the recurrent infections are caused by the generally poor health of severely affected patients, as opposed to a specific immune defect. Some interesting data have been reported regarding the immune system in patients with Gaucher's disease, a more common lysosomal storage disorder that shares many clinical characteristics with MPS VII. Abnormalities in bacterial killing and superoxide generation were detected in circulating monocytes and monocyte-derived macrophages from human patients with Gaucher's disease (23). These functions improved somewhat after ERT. Although no mechanistic data were provided, this study suggests that there may be a human correlate for the defect described in this paper.
These studies demonstrate the presence of a previously undescribed immune defect in the MPS VII mouse, and provide information on the underlying mechanism. This defect involves antigen processing and can be corrected by in vivo ERT. Further work will be required to determine whether this same defect exists in human patients, and whether these results can be extrapolated more widely to lysosomal storage diseases in general. If a defect of this sort is responsible for symptoms such as the recurrent respiratory infections, it is possible that some form of ERT directed specifically to the lung could improve the quality of life for afflicted patients. In addition, conclusions about the pathogenicity and immunogenicity of various treatments in the MPS VII mouse will have to take this defect into account.
Abbreviations
- ERT:
-
enzyme replacement therapy
- GAG:
-
glycosaminoglycan
- GUSB:
-
β-glucuronidase
- HEL:
-
hen's egg lysozyme
- LN:
-
lymph node
- MPS VII:
-
mucopolysaccharidosis type VII
- OVA:
-
ovalbumin
- MHC:
-
major histocompatibility
- FACS:
-
fluorescence-activated cell sorter
- CFA:
-
complete Freund's adjuvant
- IFA:
-
incomplete Freund's adjuvant
References
Sly WS, Quinton BA, McAlister WH, Rimoin DL 1973 Beta glucuronidase deficiency: report of clinical, radiologic, and biochemical features of a new mucopolysaccharidosis. J Pediatr 82: 249–257
Sands MS, Erway LC, Vogler C, Sly WS, Birkenmeier EH 1995 Syngeneic bone marrow transplantation reduces the hearing loss associated with murine mucopolysaccharidosis type VII. Blood 86: 2033–2040
Bastedo L, Sands MS, Lambert DT, Pisa MA, Birkenmeier E, Chang PL 1994 Behavioral consequences of bone marrow transplantation in the treatment of murine mucopolysaccharidosis type VII. J Clin Invest 94: 1180–1186
Birkenmeier EH, Davisson MT, Beamer WG, Ganschow RE, Vogler CA, Gwynn B, Lyford KA, Maltais LM, Wawrzyniak CJ 1989 Murine mucopolysaccharidosis type VII: characterization of a mouse with beta-glucuronidase deficiency. J Clin Invest 83: 1258–1256
Tomatsu S, Fukuda S, Sukegawa K, Ikedo Y, Yamada S, Yamada Y, Sasaki T, Okamoto H, Kuwahara T, Yamaguchi S, Kiman T, Shintaku H, Isshiki G, Orii T 1991 Mucopolysaccharidosis type VII: characterization of mutations and molecular heterogeneity. Am J Hum Genet 48: 89–96
Sands MS, Vogler C, Kyle JW, Grubb JH, Levy B, Galvin N, Sly WS, Birkenmeier EH 1994 Enzyme replacement therapy for murine mucopolysaccharidosis type VII. J Clin Invest 93: 2324–2331
Sands MS, Vogler C, Torrey A, Levy B, Gwynn B, Grubb J, Sly WS, Birkenmeier EH 1997 Murine mucopolysaccharidosis type VII: long term therapeutic effects of enzyme replacement and enzyme replacement followed by bone marrow transplantation. J Clin Invest 99: 1596–1605
O'Connor LH, Erway LC, Vogler CA, Sly WS, Nicholes A, Grubb J, Holmberg SW, Levy B, Sands MS 1998 Enzyme replacement therapy for murine mucopolysaccharidosis type VII leads to improvements in behavior and auditory function. J Clin Invest 101: 1394–1400
Birkenmeier EH, Barker JE, Vogler CA, Kyle JW, Sly WS, Gwynn B, Levy B, Pegors C 1991 Increased life span and correction of metabolic defects in murine mucopolysaccharidosis type VII after syngeneic bone marrow transplantation. Blood 78: 3081–3092
Sands MS, Barker JE, Vogler C, Levy B, Gwynn B, Galvin N, Sly WS, Birkenmeier E 1993 Treatment of murine mucopolysaccharidosis type VII by syngeneic bone marrow transplantation in neonates. Lab Invest 68: 676–686
Watson GL, Sayles JN, Chen C, Elliger SS, Elliger CA, Raju NR, Kurtzman GJ, Podsakoff GM 1998 Treatment of a lysosomal storage disease in MPS VII mice using a recombinant adeno-associated virus. Gene Ther 5: 1642–1649
Daly TM, Okuyama T, Vogler C, Haskins ME, Muzyczka N, Sands MS 1999 Neonatal intramuscular injection with recombinant adeno-associated virus results in prolonged β-glucuronidase expression in situ and correction of liver pathology in mucopolysaccharidosis type VII mice. Human Gene Ther 10: 85–94
Wolfe JH, Sands MS, Barker JE, Gwynn B, Rowe LB, Vogler CA, Birkenmeier EH 1992 Reversal of pathology in murine mucopolysaccharidosis type VII by somatic cell gene transfer. Nature 360: 749–753
Moullier P, Bohl D, Heard JM, Danos O 1993 Correction of lysosomal storage in the liver and spleen of MPS VII mice by implantation of genetically modified skin fibroblasts. Nat Genet 4: 154–159
Daly TM, Vogler C, Levy B, Haskins ME, Sands MS 1999 Neonatal gene transfer leads to widespread correction of pathology in a murine model of lysosomal storage disease. Proc Natl Acad Sci USA 96: 2296–2300
Germain RN, Castellino F, Han R, Sousa CRE, Romagnoli P, Sadegh-Nasseri S, Zhong G-M 1996 Processing and presentation of endocytically acquired protein antigens by MHC class II and class I molecules. Immunol Rev 151: 5–30
Lorenz RG, Tyler AN, Allen PM 1988 T cell recognition of bovine ribonuclease: self/non-self discrimination at the level of binding to the I-Ak molecule. J Immunol 141: 4124–4128
Ziegler HK, Unanue ER 1982 Decrease in macrophage antigen catabolism caused by ammonia and chloroquine is associated with inhibition of antigen presentation to T cells. Proc Natl Acad Sci USA 79: 175–178
Allen PM, Unanue ER 1984 Differential requirements for antigen processing by macrophages for lysozyme-specific T cell hybridomas. J Immunol 132: 1077–1079
Vogler C, Birkenmeier EH, Sly WS, Levy B, Pegors C, Kyle JW, Beamer WG 1990 A murine model of mucopolysaccharidosis VII: gross and microscopic findings in beta-glucuronidase-deficient mice. Am J Pathol 136: 207–217
Yamada Y, Kato K, Sukegawa K, Tomatsu S, Fukuda S, Emura S, Kojima S, Masuyama T, Sly WS, Kondo N, Orii T 1998 Treatment of MPS VII (Sly disease) by allogeneic BMT in a female with homozygous A619V mutation. Bone Marrow Transplant 21: 629–634
Sewell AC, Poets CF, Degen I, Stoss H, Pontz BF 1996 The spectrum of free neuraminic acid storage disease in childhood: clinical, morphological and biochemical observations in three non-Finnish patients. Am J Med Genet 63: 203–208
Marodi L, Kaposzta R, Toth J, Laszlo A 1995 Impaired microbicidal capacity of mononuclear phagocytes from patients with type I Gaucher disease: partial correction by enzyme replacement therapy. Blood 86: 4645–4649
Chen JS, Lorenz RG, Goldberg J, Allen PM 1991 Identification and characterization of a T-cell-inducing epitope of bovine ribonuclease that can be restricted by multiple class II molecules. J Immunol 147: 3672–3678
Kobori JA, Hood L, Shastri N 1992 Structure-function relationship among T-cell receptors specific for lysozyme peptides bound to Ab or Abm-12 molecules. Proc Natl Acad Sci USA 89: 2920–2924
Grubb JH, Kyle JW, Cody LB, Sly WS 1993 Large-scale purification of phosphorylated recombinant human β-glucuronidase from over-expressing mouse L cells. FASEB J 7: 1255Aabstr
Gammon G, Shastri N, Cogswell J, Wilbur S, Sadegh-Nasseri S, Krzych U, Miller A, Sercarz E 1987 The choice of T-cell epitopes utilized on a protein antigen depends on multiple factors distant from, as well as at the determinant site. Immunol Rev 98: 53–73
Viner NJ, Nelson CA, Deck B, Unanue ER 1996 Complexes generated by the binding of free peptides to class II MHC molecules are antigenically diverse compared with those generated by intracellular processing. J Immunol 156: 2365–2368
Kint JA, Dacremont G, Carton D, Orye E, Hooft C 1973 Mucopolysaccharidosis: secondarily induced abnormal distribution of lysosomal isoenzymes. Science 181: 352–354
Avila JL, Convit J 1975 Inhibition of leukocytic lysosomal enzymes by glycosaminoglycans in vitro. J 152: 57–64
Greenwood RS, Hillman RE, Alcala H, Sly WS 1978 Sanfilippo A syndrome in the fetus. Clin Genet 13: 241–250
Wolfe JH, Martin CE, Deshmane SL, Reilly JJ, Kesari S, Fraser NW 1996 Increased susceptibility to the pathogenic effects of wild-type and recombinant herpesviruses in MPS VII mice compared to normal siblings. J Neurovirol 2: 417–422
Vervoot R, Gitzelmann R, Bosshard N, Maire I, Liebaers I, Lissens W 1998 Low beta-glucuronidase enzyme activity and mutations in the human beta-glucuronidase gene in mild mucopolysaccharidosis type VII, pseudodeficiency and a heterozygote. Human Genet 102: 69–78
Acknowledgements
The authors thank W. S. Sly for supplying recombinant human GUSB.
Author information
Authors and Affiliations
Additional information
Supported, in part, by National Institutes of Health grants HD-35671 and DK-53920 (M.S.S.), the Charles E. Culpeper Foundation (R.G.L.), and the Raymond A. Bryan IV fellowship (T.M.D.).
Rights and permissions
About this article
Cite this article
Daly, T., Lorenz, R. & Sands, M. Abnormal Immune Function In Vivo in a Murine Model of Lysosomal Storage Disease. Pediatr Res 47, 757–762 (2000). https://doi.org/10.1203/00006450-200006000-00012
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-200006000-00012
This article is cited by
-
A Hitchhiker's Guide to the Blood–brain Barrier: In Trans Delivery of a Therapeutic Enzyme
Molecular Therapy (2014)
-
Mucopolysaccharide diseases: A complex interplay between neuroinflammation, microglial activation and adaptive immunity
Journal of Inherited Metabolic Disease (2014)
-
Widespread biochemical correction of murine mucopolysaccharidosis type VII pathology by liver hydrodynamic plasmid delivery
Gene Therapy (2009)
-
Immune system irregularities in lysosomal storage disorders
Acta Neuropathologica (2008)
-
Prevention of systemic clinical disease in MPS VII mice following AAV-mediated neonatal gene transfer
Gene Therapy (2001)
