
Abstract
In this work we have undertaken a comparative study of human umbilical vein endothelial cells (HUVECs) and human saphenous vein endothelial cells (HSVECs) with respect to functional and antigenic tissue factor (TF), tissue factor pathway inhibitor (TFPI), and TF mRNA. Monolayers of each cell type (passage 2, except where specified) were grown to confluence and then activated for 4 h with either 50 U/mL IL-1-α or 10 μg/mL tumor necrosis factor-α. Activated factor X appearing in supernatant was measured using a chromogenic assay, and both Northern blots and quantitative RT-PCR were performed to assess concentrations of TF mRNA accompanying activation. The role of TFPI was separately determined by ELISA for supernatant TFPI antigen, and by measurements of production of activated factor X in the presence of 0, 5, 15, or 50 μg/mL of an antibody directed against TFPI. To address a non-TF pathway endothelial cell function, antigenic concentrations of tissue plasminogen activator for both cell types was also determined by ELISA. HUVECs were found to produce 2.4- to 3.5-fold more functional TF. No significant HUVEC-HSVEC differences were detected in TF antigen, supernatant TFPI, anti-TFPI affinity for endothelial cell-associated TFPI, TF mRNA or its amplification products, and tissue plasminogen activator. Immunostaining for TF antigen, however, may have failed to detect a modest HUVEC-HSVEC difference. Our finding with respect to functional TF indicates that HUVECs and HSVECs are not equivalent in terms of models for endothelial cell function in small children versus adults.
Similar content being viewed by others
Main
Most studies of the function of cultured ECs are still performed with infrequent regard to the tissue source (e.g. conceptus versus adult, or arteries versus microvasculature versus veins). In particular, most studies use cultured HUVECs (1), which may be appropriate for the investigation of disorders of large veins in newborns, but do not assure relevance to the older child or adult. For example, it has long been known that the pulmonary endothelium in newborn rabbits has a greater permeability to water than that for adult animals (2). Further, newborn plasma contains unusually large von Willebrand factor multimers (released from endothelium); these multimers disappear from the plasma of infants by 8 wk of age (3). Finally, because heparin sulfate and chondroitin sulfate are significantly increased in the inferior vena cava of newborn versus adult rabbits (4), antithrombin III activity may be enhanced in the newborn. For studies having relevance to the adult population, one available EC source has been saphenous vein segments left over from coronary artery bypass grafting, from which have been cultured HSVECs (5).
In this work we have undertaken a comparative study of HUVECs and HSVECs with respect to functional and antigenic TF, antigen for TFPI, TF mRNA, and, as a control for a non-TF pathway function, antigen for t-PA. TF is of interest insofar as it is now recognized to be 1) the initiator of coagulation in basic hemostasis, and 2) a principal participant in coagulation disorders associated with endotoxemia, malignancy, immune dysfunction, and atherosclerosis (6). Although TF is not normally expressed by ECs, it can be induced by a number of stimulatory agents, including IL-1, TNF, and endotoxin (7, 8). In contrast, both TFPI (9) and t-PA (10) are constitutively expressed, with relatively minor changes in concentrations after cytokine activation.
METHODS
EC monolayers.
Monolayers of each EC type were grown to confluence on 12-well (4.2 cm2/well) tissue culture dishes (Falcon; Becton Dickinson, Franklin Lakes, NJ) that had been precoated with fibronectin (adsorbed from a 10 μg/mL solution for 1 h at room temperature; Becton Dickinson). Although preliminary experiments were performed with monolayers of passages 1 through 4, the data and figures of the present work, with two exceptions, were from passage 2 cells of either type. The first exception is that passage 3 and 4 ECs were used for the Northern blots (see below). The second exception is that the t-PA determinations used passage 1 cultures for one experiment and passage 2 cultures for a second. All monolayers generally reached confluence at 7 to 10 d after passage, as described elsewhere for HUVECs (1) and HSVECs (5). Media consisted of M199 with 25 mM HEPES, to which was added 20% FCS, 150 μg/mL heparin, 50 μg/mL EC growth supplement, 2 mM L-glutamine, penicillin/streptomycin, and supplemental sodium bicarbonate. No difference in morphology was observed at confluence between HUVECs and HSVECs. Monolayers were activated with IL-1-α (50 U/mL for 4 h; Genzyme, Inc, Cambridge, MA) or, in some cases, with TNF-α, 10 μg/mL for 4 h (Genzyme). For mRNA studies, for which a larger number of ECs were required per experiment, ECs were grown on fibronectin-coated T75 culture flasks (Falcon; Becton Dickinson).
Chromogenic assay for factor Xa.
As a measure of functional TF, we determined concentrations of activated clotting factor X appearing in the supernatant of monolayers using an amidolytic technique (11). For this purpose, monolayers were incubated at 37°C with a “reaction complex” (400 μL/well) consisting of 0.01 M HEPES, 0.14 M NaCl, 10 nM factor VII, 100 nM factor X, 5.0 mM CaCl2, and 1 mg/mL BSA. In addition, parallel monolayer wells contained 15 μg/mL of a murine MAb directed against Kunitz domain 1 of TFPI (American Diagnostica, Greenwich, CT). Aliquots of postincubation reaction complex were mixed with an equal volume of 75 mM EDTA to inhibit further activation of factor X and then incubated with Spectrozyme Xa (0.5 mM final concentration; American Diagnostica, New York, NY) at 37°C for 30 min. To block further action of factor Xa on the chromogenic substrate, we then added 30% acetic acid (one part to five parts reaction complex). Samples were read in a microplate reader at 405 nm for increase in absorbance of free chromophore, a calibration curve having been constructed using known concentrations of purified factor Xa (courtesy of Dr. A. Guha, Mt. Sinai Medical Center, New York, NY). Determinations of factor Xa production in femtomoles per square centimeter per 100 000 cells, were corrected for a very small “baseline” factor Xa activity obtained in the absence of cytokine activation and in the absence of anti-TFPI (see “Results”). Cell number densities were determined by first exposing the cells in parallel culture wells to a solution of trypsin-EDTA (Sigma Chemical Co., St. Louis, MO) for less than 5 min at room temperature and then collecting and quenching the detached cells in an equal volume of complete culture medium. Cells were resuspended in PBS, and microliter aliquots were counted in a hemocytometer.
Control experiments were performed in which cultures were incubated for 30 min with a murine MAb to human TF (product no. 4509, American Diagnostica, Greenwich, CT). A concentration of 30 nM was used to inhibit TF activity on intact endothelium (11).
Anti-TFPI assay.
To characterize the reported equilibrium (12, 13) between supernatant “free” TFPI and that which is EC-associated, some monolayers were washed with HEPES buffer with 0.1% BSA and incubated with 0, 1, 5, 15, or 50 μg/mL of anti-TFPI for 30 min at 37°C. The range of the chosen antibody bracketed that found by others (15 μg/mL; (12) to substantially block the effect of TFPI. After a single wash with the same HEPES buffer, the monolayers were assayed for factor Xa production as described above. Control experiments were performed in which TF activity was assessed in the absence of cytokine activation (“baseline” activity), without and with anti-TFPI.
Immunohistochemistry for TF antigen.
TF antigen was identified using an avidin-biotin complex (Vector Laboratories, Burlingame, CA). Primary antibody was a polyclonal TF antibody raised in rabbits to the extracellular domain of recombinant soluble TF, residues 1 to 218 (courtesy of Dr. Y. Nemerson, Mt. Sinai Medical Center, New York, NY).
Isolation of TF RNA and Northern blotting.
EC monolayers were exposed in situ to a solution of trypsin-EDTA (Sigma) for less than 5 min at room temperature, and the detached cells were collected and quenched in an equal volume of complete culture medium. Cells were resuspended in PBS and counted as above. Cells were subsequently pelleted and, when necessary, stored at −80°C until use. Total cellular RNA was isolated by means of the guanidinium isothiocyanate method (14), including centrifugation through CsCl2. Isolated RNA pellets were dried with the aid of a Speedvac vacuum system (Savant Instruments, Hicksville, NY), and resuspended in buffer containing diethyl pyrocarbonate (DEPC)-treated water, deionized formamide, 37% formaldehyde, and 10× 3-(N-morpholino)propanesulfonic acid (MOPS). The RNA samples were then heated to 65°C for 15 min to facilitate denaturation and chilled on ice. Agarose gel electrophoresis of the samples was next performed. After electrophoresis, the gel was washed twice with DEPC-treated water, after which the RNA was transferred to nylon membranes (Magna Charge, Micron Separations Inc., Westboro, MA) via transblotting with 20× DEPC-SSC, and the blot was subjected to UV cross-linking. The human TF DNA probe was prepared from a 4228-bp plasmid (provided through the courtesy of Dr. Y. Nemerson, Mt. Sinai Medical Center, New York, NY) using Sma I and Eco RI to yield a 706-bp fragment, and labeled with phosphorus 32 by means of the Prime-It II kit (Stratagene, La Jolla, CA). After prehybridization for 15 min, the probe was hybridized to the blot membrane at 42°C overnight. The filter was then washed twice for 20 min in 2× SSC and 0.1% SDS at room temperature and then in 0.2× SSC/0.1% SDS, autoradiographed, and quantified using the α Imager 2000 image analysis system (α Innotech, Inc., San Leandro, CA). In particular, this analysis was applied to two Northern blots for passage 2 HSVECs versus HUVECS and for one blot each for passage 3 and passage 4 of these cells.
Quantitative RT-PCR amplification products of TF mRNA.
Total RNA from guanidine isothiocyanate extracts of cell cultures (isolated as indicated for Northern analysis) was used for RT-PCR. First-strand cDNA synthesis (RT reaction) used 4 mg total RNA and 0.2 A260 U of random hexamer primers (Boehringer-Mannheim, Indianapolis, IN) and was performed for 1 h at 37°C using 400 U of Maloney murine leukemia virus (M-MLV) reverse transcriptase (Promega, Madison, WI) in Promega reaction buffer and 40 U ribonuclease inhibitor (Boehringer-Mannheim). PCR was performed using a Mini Cycler (MJ Research, Watertown, MA). In each reaction we used 5% of the RT reaction (2 μL of a 40 μL RT total volume), 2 pmol primer oligonucleotides for TF, 1 pmol primer nucleotide for GAPDH, and 2 U Taq polymerase (Fisher Scientific, Pittsburgh, PA) in PCR reaction buffer B (Fisher Scientific), supplemented with 4 μL of 25 mM MgCl2 to a final concentration of 4 mM in a final reaction volume of 25 μL. Oligonucleotide primers for TF were synthesized from published primer sequences (15) and yielded a product of 204 bp. Those for GAPDH were constructed complementary to the known sequence for GAPDH. The primer sequences for GAPDH were AAG GTG AAG GTC GGA GTC AA for the 5′ end and TGA GTC CTT CCA CGA TAC CA for the 3′ end, resulting in a 511-bp product. Thermal cycling settings were initial denaturing at 95°C for 1 min followed by 94°C for 30 s, annealing at 50°C for 30 s, and extension at 72°C for 2 min per cycle with final extension at 72°C for 5 min. Linear amplification with respect to cycle number was established for TF and GAPDH primer pairs to be in the range of 20 to 22 cycles.
Assay for TFPI.
Antigenic concentrations of TFPI in cell culture supernatants were determined using an ELISA for total (full-length plus truncated) TFPI (product 849, American Diagnostica, Greenwich, CT). TFPI was measured from samples collected 4 h from the time media was replaced, either with fresh media containing IL-1-α or fresh media in the absence of IL-1-α. Thus, TFPI values represent TFPI produced by ECs during a 4-h period. (Preliminary experiments had shown that TFPI concentrations in complete media were less than 0.1 ng/mL.)
Assay for t-PA.
Antigenic concentrations of t-PA were measured using an ELISA for t-PA (Asserachrom t-PA, product 0248, American Bioproducts, Parsippany, NJ).
Statistics and curve fitting.
Values for factor Xa production and for concentrations of TFPI were compared using the t test. The observed data points for factor Xa production versus concentration of anti-TFPI were fitted using a least-squares SAS regression of an exponential relation of the form MATH where F is factor Xa production in femtomoles per minute per 100 000 cells, T is concentration of anti-TFPI in micrograms per milliliter, Fo is the production rate of factor Xa in the absence of anti-TFPI, Fmax is the maximum production rate of factor Xa, and k is the apparent association constant of the antibody for cell-associated TFPI in micrograms per milliliter. The regression yielded estimates of the means and standard errors for Fo, Fmax and k.

RESULTS
Functional TF activity: factor Xa production.
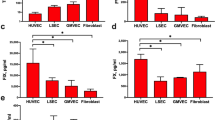
HUVECs activated with IL-1-α produced significantly more factor Xa per minute per 100 000 ECs than did HSVECs when anti-TFPI was present (Table 1). The increase (2 p= 0.045) was 2.4-fold when anti-TFPI was present at 15 μg/mL, and 3.5-fold (2 p< 0.05) based on the estimates for Fmax from the data of Figure 1 (see below). In the absence of anti-TFPI (Table 1), there still was a trend toward an increase. In two experiments, HUVECs also generated more factor Xa than HSVECs in response to TNF-α, the increases being 3.5- and 3.3-fold in the presence and absence of anti-TFPI, respectively (Table 2).

Factor Xa production vs concentration of anti-TFPI for HUVECs (A) vs HSVECs (B).
Control experiments with anti-TF in the presence of anti-TFPI showed a comparable degree of inhibition of factor Xa production with each cell type: 53.8 ± 4.6% (n= 4) for HUVECs, and 59.6 ± 5.5% for HSVECs.
Anti-TFPI studies.
The principal experiments with anti-TFPI were performed to characterize the known equilibrium between supernatant “free” TFPI and that which is EC-associated. The results are depicted in Figure 1, in which factor Xa production is plotted versus concentration of this antibody and the observed data points were fitted using the exponential relation described above. The resulting estimates (mean ± SEM) for F0 are 2.0 ± 0.6 and 0.4 ± 0.2, for Fmax are 7.4 ± 2.0 and 2.1 ± 0.3, and for k are 29.1 ± 25.3 and 12.4 ± 5.9 for HUVECs, and HSVECs, respectively. The values for F0 differ by 5.0-fold, while those for Fmax differ by 3.5-fold. The estimates for k, on the other hand, do not differ significantly.
The estimates for the apparent association constant k indicate that the affinity of anti-TFPI in solution for HSVEC-associated TFPI is at least as great as that for HUVEC-associated TFPI. This in turn suggests that the cell-associated TFPIs are in comparable equilibria (12, 13) with free TFPI in solution. Consequently, any differences in concentrations of cell-associated TFPI are likely to be reflected in differences in supernatant TFPI.
In the absence of cytokine activation, baseline production (mean ± SEM) of factor Xa, in femtomoles per minute per 100 000 cells, for four experiments was 0.10 ± 0.025 and 0.077 ± 0.031 for HUVECs and HSVECs, respectively, in the absence of anti-TFPI. In the presence of 15 μg/mL anti-TFPI, this same baseline activity was 0.14 ± 0.020 and 0.11 ± 0.037 fmol/min per 100 000 cells for HUVECs and HSVECs, respectively. Values for baseline production did not differ significantly with respect to either cell type or the presence versus absence of anti-TFPI.
Immunohistochemistry for TF antigen.
Immunostaining for TF antigen was positive for both activated HSVECs and HUVECs, but only weakly so for unactivated ECs (data not shown). However, variability in staining was such that we were unable to determine whether a difference existed in staining intensity for the two cell types.
Northern blotting.
The Northern blot of Figure 2 shows a similar TF mRNA response of the two cell types to IL-1-α. Control concentrations of GAPDH mRNA were similar in all samples. We measured OD of mRNA bands from passage 2 through 4 HSVECs and HUVECs. When normalized to OD for GAPDH, the ratios differ by less than 2-fold in each case. No TF mRNA was detected in unactivated ECs.

TF and GAPDH mRNAs for HUVECs vs HSVECs after IL-1-α activation.
Quantitative RT-PCR.
The RT-PCR amplification products of TF mRNA, shown in Figure 3, demonstrate comparable intensities of the bands corresponding to TF cDNA for IL-1-α-activated HSVECs and HUVECs after 22 and 24 cycles, respectively. When normalized to OD for GAPDH, the ratios again differ by less then 2-fold in each case. PCR amplification products for TF were not detected in unactivated ECs.

RT-PCR amplification products for TF mRNA after IL-1α activation.
TFPI.
In terms of free or supernatant TFPI, concentrations for the two EC types were similar with or without IL-1-α activation (Table 3).
t-PA.
Supernatant concentrations for the two EC types in the absence of anti-TFPI were similar with or without IL-1-α activation (Table 4).
DISCUSSION
In response to cytokine activation, HUVECs produce more functional TF than HSVECs, whereas both cell types generate comparable steady-state concentrations of TF mRNA and RT-PCR amplification products of TF mRNA, supernatant (free) TFPI, and t-PA. Although immunostaining for TF antigen allowed identification of this antigen on activated HUVECs and HSVECs, the sensitivity of this approach for cultured ECs was insufficient to resolve a difference between the two cell types. The lack of significant changes in TFPI and t-PA concentrations after cytokine activation is consistent with the previous observations of Ameri et al. (9) and Schleef et al. (10), respectively. The former group reported increases in TFPI only of the order of 25%, whereas the latter observed a 50% decrease in t-PA, but only after 24 h. The decrease in t-PA at 6 h was of the order of 25% or less, as indicated by Schleef et al.[Figure 2 of (10)]. We note, however, that the HUVEC-HSVEC difference in functional TF achieved statistical significance only in the presence of anti-TFPI. We believe this was so because the use of anti-TFPI not only enhanced measurable values for factor Xa production, but also reduced scatter in these values insofar as TFPI is noncovalently (and loosely) cell-associated and readily, if variably, removed by washing steps. Results obtained in the absence of anti-TFPI, although not achieving statistical significance, further suggest an increase in functional TF activity in HUVECs. In fact, such an increase was observed for all five paired (same day) comparisons. The lack of complete inhibition of TF activity with the anti-TF antibody may have occurred because the specific antibody used recognizes an epitope not identical with the functional domain of the TF molecule and/or cannot sterically access all TF copies on an EC monolayer. An alternative explanation is that the factor Xa assay may in part reflect as yet undiscovered, non-TF factor X proconversion activities of activated ECs. Unactivated HUVECs and HSVECs produced negligible concentrations of factor Xa and undetectable TF mRNA by both Northern blotting and quantitative RT-PCR.
The precise mechanism for the posttransciptional difference in functional TF activity remains unknown. Nonetheless, differences in EC function from one vascular bed to another are not unknown, there being a number of reports of heterogeneities in cell responsiveness to cytokines and antigen presentation. These reports concern heterogeneity of dermal microvascular ECs with respect to the expression of adhesion molecules such as CD36 and VCAM-1 compared with expression seen with HUVECs (16), differing expression of monocyte-related antigens by arteriolar, capillary, and venular ECs (17), and differing expression of molecules of the MHC and of the von Willebrand factor-related antigen (18). HUVECs themselves are heterogeneous with respect to cytokine-induced adhesion molecules when considered with respect to genetically distinct cell lines (19). The mechanism for the HSVEC-HUVEC TF difference may involve 1) differing TF structural sequences, 2) posttranslational processing of TF (e.g. leading to a difference in TF interaction with factor VII), and 3) the proportion of TFPI consisting of truncated (relatively inactive) forms.
In any event, an apparent dominance of the TF/factor VII pathway in early and later fetal life is also suggested by the work of Hassan et al. (20). These investigators compared fetal and adult liver with respect to mRNAs and protein concentrations for factors VII, IX, and X using dot and Northern blot hybridization and Western blot analysis. In contrast to low concentrations for factor IX, they found in fetal liver somewhat higher concentrations of factors VII and X, the clotting factors most directly involved in the TF pathway. Further, Solberg et al. (21) reported that HUVECs produced more functional TF in response to thrombin or endotoxin than intact adult saphenous veins. However, cell cultures (or intact veins) of the two EC types were not compared directly, and the method used by these investigators may have been insensitive to the low concentrations of functional TF generated by adult saphenous ECs. Recently, Solovey et al. (22) reported expression of functional TF by circulating ECs in patients with sickle cell anemia. Neither the role of TFPI nor the anatomic origin of the circulating ECs was addressed by these workers. However, their data indicate an abnormally high expression of TF in sickle cell patients in both steady state and during vaso-occlusive crises.
Our findings with respect to the TF/factor VII pathway indicate that HUVECs and HSVECs are not equivalent in terms of models for EC function in small children versus adults. This result may also help explain why newborns have a shorter bleeding time than adults (23), although at the same time most studies of newborn platelet function show hypofunction (24, 25) and none demonstrate hyperfunction. Newborns, on the other hand, have lower concentrations of TFPI than older children and adults, as inferred from cord blood samples (26). In view of the present TFPI findings, such concentrations suggest that TFPI in vivo may have a shorter half-life in the newborn. Lower in vivo concentrations of TFPI also indicate an even greater role for the TF/factor VII pathway in newborns.
Abbreviations
- HUVECs:
-
human umbilical vein endothelial cells
- HSVECs:
-
human saphenous vein endothelial cells
- ECs:
-
endothelial cells
- TF:
-
tissue factor
- TFPI:
-
tissue factor pathway inhibitor
- TNF-α:
-
tumor necrosis factor-α
- RT-PCR:
-
reverse transcriptase-PCR
- t-PA:
-
tissue-plasminogen activator
- factor Xa:
-
activated clotting factor X
References
Jaffe EA, Nachman RL, Becker CG, Minick CR 1973 Culture of human endothelial cells derived from umbilical veins. J Clin Invest 52: 2745–2756
Milligan RG, Wangensteen OD 1978 Pulmonary microvascular filtration and reflection coefficients in two- and four-week old rabbits. Microvasc Res 15: 331–338
Katz JA, Moake JL, McPherson PD, Weinstein MJ, Moise KJ, Carpenter RJ, Sala DJ 1989 Relationship between human development and disappearance of unusually large von Willebrand factor multimers from plasma. Blood 73: 1851–1858
Nitschmann E, Berry L, Bridge S, Dereske M, Richardson M, Monagle P, Chan A, Andrew M 1998 Morphologic and biochemical features affecting the antithrombotic properties of the inferior vena cava of rabbit pups and adult rabbits. Pediatr Res 43: 62–67
Hasson JE, Wiebe DH, Sharefkin JB, Abbott WM 1986 Migration of adult human vascular endothelial cells: effect of extracellular matrix proteins. Surgery 100: 384–391
Camerer E, Kolsto AB, Prydz H 1996 Cell biology of tissue factor, the principal initiator of blood coagulation. Thromb Res 81: 1–41
Bevilacqua MP, Pober JS, Majeau JS, Cotran RS, Gimbrone MA 1984 Interleukin-1 (IL-1) induces biosynthesis and cell surface expression of procoagulant activity in human vascular endothelial cells. J Exp Med 160: 618–623
Brox JH, Osterud B, Bjorklid E, Fenton JW II 1984 Production and availability of thromboplastin in endothelial cells. Br J Haematol 57: 239–246
Ameri A, Kuppuswamy MN, Basu S, Bajaj SP 1992 Expression of tissue factor pathway inhibitor by cultured endothelial cells in response to inflammatory mediators. Blood 79: 3219–3236
Schleef RR, Bevilacqua MP, Sawdey M, Gimbrone MA, Loskutoff DJ 1988 Cytokine activation of vascular endothelium: effects on tissue-type plasminogen activator and type 1 plasminogen activator inhibitor. J Biol Chem 263: 5797–5803
Grabowski EF, Zuckerman DB, Nemerson Y 1993 The functional expression of tissue factor by fibroblasts and endothelial cells under flow conditions. Blood 50: 3265–3270
Hansen JB, Sandset PM, Huseby KR, Huseby NE, Nordoy A 1996 Depletion of intravascular pools of tissue factor pathway inhibitor (TFPI) during repeated or continuous intravenous infusion of heparin in man. Thromb Haemost 76: 703–709
Kokawa T, Enjyoji KI, Kumeda K, Kamikubo YI, Harada-Shiba M, Koh H, Tsushima M, Yamamoto A, Kato H 1996 Measurement of the free form of TFPI antigen in hyperlipidemia. Arterioscler Thromb Vasc Biol 16: 802–808
Chomczynski P, Sacchi N 1987 Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 162: 156–159
Matsumoto Y, Kawai Y, Watanabe K, Sakai K, Murata M, Handa M, Nakamura S, Ikeda Y 1998 Fluid shear stress attenuates tumor necrosis factor-alpha-induced tissue factor expression in cultured human endothelial cells. Blood 91: 4164–4172
Page C, Rose M, Yacoub M, Pigott R 1992 Antigenic heterogeneity of vascular endothelium. Am J Pathol 141: 673–683
Turner RR, Beckstead JH, Warnike RA, Wood GS 1997 Endothelial cell phenotypic diversity. Am J Clin Pathol 87: 569–575
Petzelbauer P, Bender JR, Wilson J, Pober JS 1993 Heterogeneity of dermal microvascular endothelial cell antigen expression and cytokine responsiveness in situ and in cell culture. J Immunol 151: 5062–5072
Bender JR, Sadeghi MM, Watson C, Pfau S, Pardi R 1994 Heterogeneous activation thresholds to cytokines in genetically distinct endothelial cells: evidence for diverse transcriptional responses. Proc Natl Acad Sci USA 91: 3994–3998
Hassan HJ, Leonardi A, Chelucci C, Macioce G, Guerriero R, Russo G, Mannucco PM, Peschle C 1990 Blood coagulation factors in human embryonic-fetal development: preferential expression of the FVII/tissue factor pathway. Blood 76: 1158–1164
Solberg S, Osterud B, Larsen T, Sorlie D 1990 Lack of ability to synthesize tissue factor by endothelial cells in intact human saphenous veins. Blood Coagul Fibrinolysis 1: 595–600
Solovey A, Gui L, Key NS, Hebbel RP 1988 Tissue factor expression by endothelial cells in sickle cell anemia. J Clin Invest 101: 1899–1904
Andrew M, Paes B, Bowker J, Vegh P 1990 Evaluation of an automated bleeding time device in the newborn. Am J Hematol 35: 275–277
Tandini S, Kurekci AE, Koseoglu V, Kurt M, Ozcan O 1995 The normalization of platelet aggregation in newborns. Thromb Res 80: 57–62
Rajasekjar D, Kestin AS, Bednarek FJ, Ellis PA, Barnard MR, Michelson AD 1994 Neonatal platelets are less reactive than adult platelets to physiological agonists in whole blood. Thromb Haemost 72: 957–963
Reverdiau-Moalic C, Delahousse B, Body G, Bardos P, Leroy J, Gruel Y 1996 Evolution of blood coagulation activators and inhibitors in the healthy human fetus. Blood 88: 900–906
Author information
Authors and Affiliations
Additional information
Supported by grants HL 33095 and HL52183 from the National Heart, Lung, and Blood Institute.
Rights and permissions
About this article
Cite this article
Grabowski, E., Carter, C., Ingelfinger, J. et al. Comparison of Tissue Factor Pathway in Human Umbilical Vein and Adult Saphenous Vein Endothelial Cells: Implications for Newborn Hemostasis and for Laboratory Models of Endothelial Cell Function. Pediatr Res 46, 742 (1999). https://doi.org/10.1203/00006450-199912000-00001
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-199912000-00001
