
Abstract
Kasabach-Merritt Syndrome (KMS) is seen in children with large vascular tumors. KMS is characterized by very low platelet counts and a consumption of coagulation factors causing life-threatening complications. It has been proposed that thrombopenia in these patients is caused by intratumoral trapping of platelets. The truncated form of the cMpl-receptor ligand thrombopoietin, pegylated human megakaryocyte growth and development factor (Peg-rHuMGDF), is an agent that stimulates platelet production. We hypothesized that stimulation of the platelet production would prevent the life-threatening complications of patients with KMS owing to low platelet counts. In a mouse model of KMS, with tumors derived from a hemangioendothelioma cell line, we studied the effect of Peg-rHuMGDF. Treatment with Peg-rHuMGDF (10 μg/kg/day intraperitoneally) increased platelet counts by 7–8-fold compared with control tumor-bearing mice after 11 d of treatment (p< 0.001, n= 8). Survival was significantly increased, with 50% of treated animals alive at 1 mo versus 0% in untreated controls. Interestingly, we also observed an inhibition of tumor growth by 75% (p< 0.001, n= 8). Hematoxylin and eosin staining showed fresh fibrin clots in the treated tumors, suggesting that higher platelet counts caused intravascular thrombosis of tumor vessels. We conclude that increased platelet production in this model of KMS resulted in an antivascular tumor effect via platelet trapping. Further, we propose that thrombopoietin may be of critical value in preventing life-threatening complications from KMS.
Similar content being viewed by others


Main
In 1940, Kasabach and Merritt (1) described for the first time an infant with a vascular anomaly accompanied by thrombocytopenic purpura. This disease, named Kasabach-Merritt syndrome (KMS), is associated with thrombocytopenia, petechiae, spontaneous bleeding, and enlargement of the hemangioma. Based on reviewing the world literature in 1988, the mortality rate is estimated to be 12% or higher as a result of severe bleeding, sepsis, or invasion in vital organs (2, 3). In comparison with normal platelet counts of ±250,000–500,000/μL in children (0–5 yr), children with KMS typically have counts of 25,000/μL (range 3,000–60,000) with a consumptive coagulopathy-like state (3). The vascular tumors associated with KMS are large, red to purple, warm, and edematous. These tumors can be characterized by an infiltrative growth pattern, spindling of endothelium, microthrombi, and hemosiderin (3). It has also been demonstrated that platelets trap in these vascular anomalies (4, 5). Thus far, no universally successful treatment has been found, although pharmacologic therapy (including steroids, antiplatelet-aggregating agents, and antifibrinolytic agents), radiotherapy, and surgery have been tried (reviewed in Refs. 3 and 6). Platelet transfusions have been given in life-threatening situations, although exacerbations of s.c. bleeding and swelling of the tumors have been reported presumably due to intratumoral trapping and activation of platelets (6, 7). If possible, such transfusions should be avoided.
In 1994, thrombopoietin (Tpo), a growth factor for megakaryocytes, was discovered to be a ligand for the receptor encoded by the c-mpl proto-oncogene (c-Mpl) (8). The c-Mpl receptor is a member of the hematopoietin receptor super family and is involved in megakaryocytopoiesis (9). In addition to Tpo, a truncated form of Tpo that contains the Mpl receptor binding domain was cloned as the megakaryocyte and development factor (MGDF) (10). In clinical trials, both compounds have been shown to elevate platelet counts and hasten platelet recovery in patients (11, 12). For patients with the Kasabach-Merritt syndrome, we hypothesized that stimulation of platelet production may be helpful to prevent life-threatening situations. Stimulation of endogenous platelet production may avoid the adverse side effects of platelet transfusions because there is no partial activation due to storage (13). To test this hypothesis, we stimulated platelet production by Peg-rHuMGDF in a mouse model of KMS (14). In this murine model, mice die from hematologic changes similar to those found in the KMS in human beings including platelet aggregates, areas of hemorrhage in the tumors, and a reduced red cell survival (14).
METHODS
Peg-rHuMGDF was kindly provided by Amgen Inc. (Thousand Oaks, CA) after the institutional board obtained approval for this study. 10 μg/kg of Peg-rHuMGDF, diluted in 0.1% homologous mouse serum diluted in saline, was administered daily by intraperitoneal injection. Treatment was started when mean tumor volume was at least 150 mm3. Controls were treated with daily intraperitoneal injections of saline.
Cell culture.
The murine hemangioendothelioma cell line EOMA and bovine capillary endothelial cells (BCEs) were maintained in Dulbecco's modified Eagle's medium supplemented with 10% bovine calf serum and 1% glutamine-penicillin-streptomycin in 37°C in a 10% CO2 incubator. BCEs grew on precoated plates with 1% gelatin.
In vivo model of the Kasabach-Merritt syndrome.
Six-wk-old male 129J mice, the favorable mice for growing EOMA tumors, were obtained from Jackson Laboratory (Bar Harbor, ME). They were housed in the animal research facilities of Children's Hospital. After trypsinizing and one PBS wash, 1 × 107 Eoma cells were diluted in 1 mL of PBS. Subsequently, 1 × 106 cells (=100 μL) were injected s.c. under sterile conditions in the proximal midline of the back of a mouse. Length and width of tumors were measured to calculate tumor volume with the formula length × (width)2 × 0.52. Mice were weighed once a week. Animal care was reviewed by the animal care committee and maintained in accordance with constitutional guidelines.
Platelet counts.
By tail bleeding, 20-μL samples of blood were sampled in unopette capillary pipettes and diluted in reservoirs of unopette microcollection systems (Becton Dickinson, Franklin Lakes, NJ). Within 1 h, platelets were counted under phase contrast microscopy in a microchamber.
Immunohistochemistry.
Tumor tissues were fixed in Carnoy's fixative and embedded in paraffin according to standard histologic procedures. Tissue sections (4–6 μM) were pretreated with 2 μg/mL proteinase K (Boehringer Mannheim, Mannheim, Germany) at 37°C for 15 min. Staining was performed with a rabbit polyclonal antibody against human von Willebrand factor (DAKO, Carpinteria, CA). Subsequently, a secondary antibody against rabbit conjugated to horseradish peroxidase (DAKO) was used, and finally diaminobenzidine tetrahydrochloride (DAKO) was used as a chromagen. After counter staining with methyl green (Schmid & Co., Stuttgart, Germany), they were mounted in Permount (Fisher Scientific). H&E staining was performed according to standard procedures.
Proliferation assay.
Eoma or BCE cells (2 × 103) in 100 μL were plated in 96-well plates. The next day, Peg-rHuMGDF in a concentration range from 0 to 40 ng/mL in triplicate was added in each well. The effects of Peg-rHuMGDF were studied on basic fibroblast growth factor (bFGF, 1 ng/mL) stimulated or unstimulated BCEs. bFGF is an angiogenic growth factor that stimulates proliferation of endothelial cells. After 72 h, the cells were washed with PBS and fixed with ethanol. Subsequently, the cells were stained with methylene blue in borate buffer for 10 min. The plates were then thoroughly washed and dried. After the addition of 100 μL of 0.1 M HCl per well and a 20-min incubation at 37°C, plates were read at 600 nm. Colorization of the HCl represents the cell number, which is expressed by the plate reader in numbers (15).
Corneal neovascularization assay.
In the stroma of the mouse cornea adjacent to the limbus, pellets were implanted with bFGF as described previously (16). In brief, after anesthetizing the mice, 0.4 × 0.4-mm pockets were made in the cornea. Subsequently, 80 ng bFGF pellets were implanted 1.0–1.2 mm from the limbal vessels. Then, erythromycin was topically applied (E. Fougera, Melville, NY). The vascular response to the bFGF pellets was measured 5 d after implantation by maximal vessel length and number of clock hours of neovascularization. The area of corneal neovascularization was calculated by using a modified formula of a half ellipse, which best approximates the area of neovascularization: Area (mm2) = [πp× clock hours × length (mm) × 0.2 mm].
Statistical analysis.
Results are expressed as mean ± SEM. Instat (Mac) program was used for statistical analysis. Statistically significant differences were calculated by unpaired t test.
RESULTS
Platelet counts.
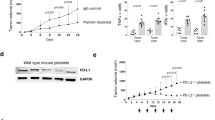
Once the tumor reached a size of 150–200 mm3 (approximately 4 d after implantation), platelet counts were already depressed from normal levels of 800,000 ± 71,000/μL to 600,000 ± 59,000/μL (p< 0.01). After an additional 11 d, platelet counts in untreated animals had fallen to 300,000 ± 102,240/μL. However, mice treated with Peg-rHuMGDF had significantly elevated platelet counts of 2.4 million ± 0.56/μL (p< 0,001, as shown in Fig. 1). This elevation was sustained at d 18 in treated animals (platelet counts 1.46 million ± 0.18/μL), when most of the untreated control animals had died. Platelet counts of untreated mice that were still alive at d 18 were 151,000 ± 22,000/μL (mean of multiple experiments). The cause of death of these mice was assumed to be the result of severe thrombocytopenia and subsequent anemia, as described by others (14).

Platelet counts in eoma-bearing mice. Platelet counts at d 11 of treatment with 10 μg/kg of Peg-rHu-MGDF vs PBS as control.
Survival.
The mice were treated for a 30-d period. At the conclusion of treatment, 50% of treated mice were still alive versus none of the untreated animals. This can also be expressed as a minimum of 9 d of prolongation of survival in treated animals versus control (p< 0.001, n= 8). Figure 2 shows a representative experiment.

Survival of eoma-bearing mice as expressed in percentage alive (treatment with 10 μg/kg Peg-rHu-MGDF, n= 4, controls n= 5).
Tumor growth.
Treatment with Peg-rHuMGDF inhibited tumor growth by 75% (p< 0.001, n= 8). Figure 3 shows a representative experiment. In a nonplatelet-trapping mouse tumor model (Lewis lung carcinoma), Peg-rHuMGDF did not significantly effect tumor growth after 14 d of treatment, despite the elevation of platelet counts by 2–3-fold (data not shown).

Tumor growth during treatment with Peg-rHu-MGDF (treated vs controls = 75% inhibition).
Immunohistochemistry.
H&E staining of embedded tumor tissue showed prominent endothelial cells and many mitoses of the Eoma cells. The treated tumors were more solid than the untreated, which had more intratumoral spaces filled with blood cells. In addition, intraluminal fresh clots and organized clots were observed in the treatment group, indicative for infarction of the tumors. We were unable to count microvessel density of the tumors as a result of a diffuse staining by von Willebrand factor antibody of both endothelial and Eoma cells.
Cell culture experiments.
Peg-rHuMGDF (20 ng/mL) had no effect on the proliferation of Eoma cells in vitro. In addition, it did not influence the proliferation of BCEs, which were cultured either with or without stimulation of bFGF.
Corneal neovascularization assay.
Peg-rHuMGDF treatment (10 μg/kg) for 5 d did not produce a significant inhibition on bFGF-induced corneal neovascularization (p= 0.08, area of corneal neovascularization = 2.0 mm2 ± 0.2, n= 8 versus 2.2 mm2 ± 0.2, n= 8 for the treatment group and the controls, respectively).
DISCUSSION
Kasabach-Merritt syndrome is a life threatening complication in children with vascular anomalies growing as kaposiform hemangioendothelioma (3, 6). The mortality rate is 12% or higher (2). Patients die of bleeding, sepsis, and/or invasion of vital structures. Platelet transfusions have been given in these children to prevent them from bleeding to death but can worsen the clinical condition (4). It is unknown why transfusions can cause these unwanted effects, but it is assumed that the platelets are immediately trapped by the tumor, become activated, and release their growth factors. These growth factors may cause the adverse effects that have been observed. Storage of platelets, before transfusion, is known to activate platelets to some extent (13).
In a mouse model of the KMS in which mice die from bleeding complications, including anemia (14), treatment with Peg-rHuMGDF increased platelet counts by 7–8-fold, preventing bleeding and lethargy, resulting in significantly prolonged survival. The elevation of platelet counts to levels several times normal is not unusual and is also seen when Peg-rHuMGDF is given to patients with advanced cancer before chemotherapy (11). Interestingly, the ability of Peg-rHuMGDF to enhance platelet production in a mouse model of KMS refutes the assumption that platelet production is already maximally stimulated by endogenous thrombopoietin. Indeed, there is considerable evidence that platelets are self-regulating the activity of endogenously produced thrombopoietin. They express the Mpl-receptor expressed on their cell surface and scavenge free-circulating thrombopoietin (17). It has been found that patients with thrombopenia owing to a decreased production of platelets (bone marrow disease) have very high plasma thrombopoietin levels. In contrast, patients with thrombopenia owing to an increased platelet turnover from autoimmune thrombocytopenia have plasma thrombopoietin concentrations within normal range, indicating that platelets are still able to bind free thrombopoietin even when circulating for only a short time (18). This last possibility may explain why treatment with Peg-rHu-MGDF can increase platelet counts in KMS mice. Based on clinical studies (4), it is expected that the Eoma-bearing mice suffer from an increased platelet turnover rather than a bone marrow failure.
The mechanism of how increased platelet production inhibited tumor growth by 75% remains unknown, but there are several hypotheses. 1) Trapped platelets could inhibit tumor growth by secretion of angiogenesis inhibitors, like platelet factor 4 and thrombospondin, and this secretion may be in excess to the released angiogenesis stimulators by platelets (e.g. vascular endothelial growth factor) (19). Although possible, it seems unlikely that the secretion products of platelets have an inhibitory effect on angiogenesis, as serum (containing the secretion products of platelets) is generally stimulatory for cell lines and primary cultures of endothelial cells (20). 2) Peg-rHuMGDF may inhibit angiogenesis directly or has a direct inhibitory action on the EOMA cells in vivo. We were unable to find an inhibitory activity of Peg-rHuMGDF on cultured EOMA or endothelial cells. As well, Peg-rHuMGDF was not antiangiogenic in an in vivo assay of corneal neovascularization. Further, using a murine tumor (Lewis lung carcinoma), no antiangiogenic inhibition of tumor growth was detected, despite an elevation of platelet counts to over 3 million with Peg-rHuMGDF. 3) The combination of a persistent increase in platelet counts and a platelet-trapping tumor may cause thrombosis of tumor vessels, similar to antivascular therapy (21). The latter possibility was supported by H&E staining of the tumor specimens. Fresh fibrin clots were observed in the treated specimens, whereas clots could not be found in the untreated group. Similar to our finding, it has been described in a case report of a patient with KMS that generating intratumoral thrombosis by aminocaproic acid and cryoprecipitate can be effective (22). Additionally, the observation of a lack of effect in the Lewis lung carcinoma also supports the idea that this antivascular effect is seen only in tumors that trap platelets to such an extent. These data may explain why isolated transfusions of platelets have not shown a benefit in the clinic due to the lack of persistent elevation of platelets necessary for the antivascular effect.
In conclusion, these data indicate in a preclinical model that stimulation of the platelet production is effective against KMS. Two antiangiogenic drugs, angiostatin (23) and TNP-470 (24), have been reported to potently inhibit tumor growth in this mouse model of KMS. However, TNP-470 is currently being tested in the phase-2 clinical trial, and angiostatin is not yet available for patients. Although the stimulator of the platelet production Peg-rHuMGDF used in this study is not available for clinical use anymore, the recombinant form of the natural circulating Tpo is currently being tested in clinical trials. In addition, another stimulator of platelet production, IL-11, is also under clinical investigation in adults. It is expected that these agents will be effective in children (newborns) as well, because Tpo was capable of inducing a dose-dependent proliferation of megakaryocyte precursors of preterm and term babies (25). Based on our findings, we expect that Tpo and IL-11 may be of benefit in life-threatening complications for children suffering from KMS, for whom no effective treatment is known.
Abbreviations
- KMS:
-
Kasabach-Merritt syndrome
- Tpo:
-
thrombopoietin
- c-Mpl:
-
c-mpl proto-oncogene
- Peg-rHu-MGDF:
-
pegylated recombinant human megakaryocyte growth and development factor
- H&E:
-
hematoxylin and eosin
- BCE:
-
bovine capillary endothelial cells
- bFGF:
-
basic fibroblast growth factor
References
Kasabach HH, Merritt KK 1940 Capillary hemangioma with extensive purpura: report of a case. Am J Dis Child 59: 1063–1070
El-Dessouky M, Azmy AF, Raine PAM, Young DG 1988 Kasabach-Merritt syndrome. J Pediatr Surg 23: 109–111
Sarkar M, Mulliken JB, Kozakewich HPW, Robertson RL, Burrows PE 1997 Thrombocytopenic coagulopathy (Kasabach Merritt Phenomenon) is associated with Kaposiform hemangioendothelioma and not with common infantile hemangioma. Plast Reconstr Surg 100: 1377–1386
Konras SB, Green OC, King L, Duran RJ 1963 Giant hemangioma with thrombocytopenia. Am J Dis Child 105: 188–195
Warrell RP Jr, Kempin SJ, Benua RS, Reiman RE, Young CW 1983 Intratumoral consumption of indium-111 labeled platelets in a patient with hemangiomatosis and intravascular coagulation (Kasabach-Merritt syndrome). Cancer 52: 2256–2260
Enjolras O, Wassef M, Mazoyer E, Frieden IJ, Rieu PN, Drouet L, Taieb A, Stalder JF, Escande JP 1997 Infants with Kasabach-Merritt syndrome do not have “true” hemangiomas. J Pediatr 130: 631–640
Phillips WG, Marsden JR 1993 Kasabach-Merritt syndrome exacerbated by platelet transfusion. J R Soc Med 86: 231–232
Kaushansky K, Lok S, Holly RD, Broudy VC, Lin N, Bailey MC, Forstrom JW, Buddle MM, Oort PJ, Hagen FS, Roth GJ, Papayannopoulou T, Foster DC 1994 Promotion of megakaryocyte progenitor expansion and differentiation by the c-Mpl ligand thrombopoietin. Nature 369: 568–574
Sauvage FJ, Hass PE, Spencer SD, Malloy BE, Gurey AL, Spencer SA, Darbonne WC, Henzel WJ, Wong SC, Kuang WJ, Oles KJ, Hultgren B, Solberg LA, Goeddel DV, Eaton DL 1994 Stimulation of megakaryocytopoiesis and thrombopoiesis by the c-Mpl ligand. Nature 369: 533–538
Bartley TD, Bogenberger J, Hunt P, Li YS, Lu HS, Martin F, Chang MS, Samal B, Nichol JL, Swift S, Johnson MJ, Su RY, Parker VP, Suggs S, Skrine JD, Merewether LA, Clogston C, Hsu E, Hokom MM, Hornkohl A, Choi E, Pangelinan M, Sun Y, Mar V, McNinch J, Simonet L, Jacobsen F, Xie C, Schutter JE, Basu R, Selander L, Trollinger D, Sieu L, Padilla D, Trail G, Elliott G, Izumi R, Covey T, Crouse J, Garcia A, Xu W, Del Castillo J, Biron J, Cole S, Hu MCT, Pacifici R, Ponting I, Saris C, Wen D, Yung YP, Lin H, Bosselman RA 1994 Identification and cloning of a megakaryocyte growth and development factor that is a ligand for the cytokine receptor Mpl. Cell 77: 1117–1124
Basser RL, Rasko JE, Clarke K, Cebon J, Green MD, Hussein S, Alt C, Menchaca D, Tomita D, Marty J, Fox RM, Begley CG 1996 Thrombopoietic effects of pegylated recombinant human megakaryocyte growth and development factor in patients with advanced cancer. Lancet 348: 1279–1281
Nimer SD 1997 Platelet stimulating agents: off the launching pad. Nature Med 3: 154–155
Metcalfe P, Williamson LM, Reutelingsperger CP, Swann I, Ouwehand WH, Goodall AH 1997 Activation during preparation of therapeutic platelets affects deterioration during storage: a comparative flow cytometric study of different production methods. Br J Haematol 98: 86–95
Hoak JC, Warner ED, Cheng HF, Fry GL, Hankenson RR 1971 Hemangioma with thrombocytopenia and microangiopathic anemia (Kasabach-Merritt syndrome): an animal model. J Lab Clin Med 77: 941–950
Goldman R, Bar-Shavitz Z 1979 Dual effect of normal and stimulated macrophages and their conditioned media on target cell proliferation. J Natl Cancer Inst 63: 1009–1016
Kenyon BM, Voest EE, Chen C, Flynn E, Folkman J, D'Amato RJ 1996 A model of angiogenesis in the mouse cornea. Invest Opthalmol Vis Sci 37: 1625–1632
Moliterno AR, Hankins WD, Spivak JL 1998 Impaired expression of the thrombopoietin receptor by platelets from patients with polycythemia vera. N Engl J Med 338: 572–580
Porcelijn L, Folman CC, Bossers B, Huiskes E, Overbeeke MAM, van der Schoot CE, de Haas M, von dem Borne AE 1998 The diagnostic value of thrombopoietin level measurement in thrombocytopenia. Thromb Haemost 79: 1101–1115
Verheul HMW, Hoekman K, Luykx-de Bakker S, Eekman J, Broxterman HJ, Pinedo HM 1997 Platelet: transporter of vascular endothelial growth factor. Clin Cancer Res 3: 2187–2190
Pinedo HM, Verheul HMW, D'Amato RJ, Folkman J 1998 Involvement of platelets in angiogenesis?. Lancet 352: 1775–1777
Huang X, Molema G, King S, Watkins L, Edington TS, Thorpe PE 1997 Tumor infarction in mice by antibody directed targeting of tissue factor to tumor vasculature. Science 275: 547–550
Warrel RP Jr, Kempin SJ 1985 Treatment of severe coagulopathy in the Kasabach-Merritt syndrome with aminocaproic acid and cryoprecipitate. N Engl J Med 313: 309–312
Lanutti BJ, Gately ST, Quevedo ME, Soff GA, Paller S 1997 Human angiostatin inhibits murine hemangioendothelioma tumor growth in vivo. Cancer Res 57: 5277–5280
O'Reilly MS, Brem H, Folkman J 1995 Treatment of murine hemangioendotheliomas with the angiogenesis inhibitor AGM-1470. J Pediatr Surg 30: 325–330
Murray NA, Waits TL, Roberts IA 1998 Endogenous thrombopoietin levels and effect of recombinant human thrombopoietin on megakaryocyte precursors in term and preterm babies. Pediatr Res 43: 148–151
Acknowledgements
The authors thank Dr. Antonio Perez-Atayde, pathologist at Children's Hospital, for his interpretation of the immunohistochemical stainings.
Author information
Authors and Affiliations
Additional information
This study was supported by a grant to the Children's Hospital from Entremed (Rockville, MD). H.M.W.V. is the recipient of the Margot Mattheijssen-van der Voort fellowship and a grant from the Nijbakker Morra Society.
Rights and permissions
About this article
Cite this article
Verheul, H., Panigrahy, D., Flynn, E. et al. Treatment of the Kasabach-Merritt Syndrome with Pegylated Recombinant Human Megakaryocyte Growth and Development Factor in Mice: Elevated Platelet Counts, Prolonged Survival, and Tumor Growth Inhibition. Pediatr Res 46, 562 (1999). https://doi.org/10.1203/00006450-199911000-00012
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-199911000-00012
This article is cited by
-
Integrin β3/Akt signaling contributes to platelet-induced hemangioendothelioma growth
Scientific Reports (2017)
