
Abstract
To analyze newborn cerebrovascular autoregulation, middle cerebral arteries from 3-4-d-old piglets were cannulated, and diameter changes after transmural pressure variation were measured. After an equilibration period at 30 mm Hg, pressure was modified from 10 to 70 mm Hg in 20-mm Hg steps. Segments with endothelium showed vasodilation during pressure decrease and vasoconstriction during pressure increase. In each case the maximum response was about 5% that of the resting diameter. Segments without endothelium responded passively to pressure change. Vasodilation during pressure decrease was reduced by the preferential calcium-activated potassium (KCa) channel blocker, tetraethylammonium (1 mM), and was absent with the nitric oxide (NO) synthase inhibitor NG-nitro-L-arginine methylester (L-NAME, 10 µM). The NO synthase substrate, L-arginine (10µM), counteracted the dilation blockade caused by L-NAME. The cyclooxygenase inhibitor indomethacin (10 µM) and the endothelin A receptor antagonist BQ-123 (10 µM) eliminated the pressure increase-induced vasoconstriction. The ATP-sensitive potassium channel blocker, glibenclamide (1 µM), and the endothelin B receptor antagonist, BQ-788 (10 nM), did not modify the autoregulatory response. None of these drugs modified the passive changes produced by pressure variations in segments without endothelium. These results suggest that: 1) piglet middle cerebral artery autoregulation is endothelium-dependent;2) NO and KCa channels are involved in vasodilation during transmural pressure decrease, and 3) endothelin-1, through endothelin A receptors, and prostanoids mediate vasoconstriction during pressure increase.
Similar content being viewed by others

Main
Cerebral arterial autoregulation is of particular importance after birth; the impairment of this homeostatic mechanism has a chief role in the pathophysiology of the ischemic and hemorrhagic insults in newborn brains(1). Previous studies characterizing the response of newborn cerebral vessels to intraluminal pressure changes have been performed mainly on pial arterioles using the close cranial window(2) but not on large cerebral arteries. However, large cerebral arteries are more important than arterioles for cerebral autoregulation at physiologic perfusion pressure levels(3). Results from studies on cerebral arterioles cannot be extrapolated to large cerebral arteries because their differences in diameter imply a different magnitude in their myogenic response(4). In addition, some endothelial factors play a more important role in cerebral arteries than in arterioles(5). There is a recent report analyzing, in vitro, the autoregulation of newborn piglet large cerebral arteries in response to flow changes(6). However, in newborn animals the absolute level of cerebral blood flow as well as the setting of autoregulatory limits seem to be determined mainly by blood pressure(7).
In vitro studies concerning the response of large cerebral arteries to intraluminal pressure change have usually been performed in adult animals. In such studies, the importance of endothelium in the autoregulatory responses is controversial. In resistance arteries from different organs, changes in vessel diameter during pressure variations are independent of endothelium(8), whereas the removal of endothelium in cat cerebral arteries is followed by a loss of autoregulation(9). Although there are no studies analyzing the effect of endothelium removal from newborn cerebral arteries, two factors suggest that the role of endothelium in the response to pressure changes might be relevant just after birth. First, large cerebral arteries from newborn lambs have less stiffness and less contractile force than those from adult sheep(10), suggesting an inferior myogenic ability in newborn cerebral arteries. Second, there is a greater response to endothelium-related vasodilators in 3-than in 10-d-old piglet cerebral arteries(11), which corresponds to the inverse relationship between the ability to relax and maturation reported in newborn lamb cerebral arteries(12).
Previous studies in newborn piglets support the relevance of cyclooxygenase products in the autoregulatory response(2,13,14). The role of ET-1(15) and potassium channels(16) in cerebrovascular responses is significant in adult animals but has not been evaluated in newborn animals. Despite the growing importance paid to NO in cerebrovascular regulation(17), whether or not it mediates the response to pressure change in either adult or newborn cerebral arteries is a matter of controversy(14,18–22).
The aim of the present study was to investigate the role that endothelium plays in the autoregulatory response of newborn large cerebral arteries, and to study the mechanisms involved in such responses. To do this, we assessed the effect of intraluminal pressure changes in segments of MCAs isolated from newborn piglets. We used a pressurized in vitro technique that allows the experiments to be performed in a controlled milieu and avoids the influence of metabolic variations(23).
METHODS
General preparation. The study protocol was approved by the Committee of Investigation of the Hospital "Virgen de la Luz." Newborn piglets (3-4 d old, n = 55), harvested from a farm, were anesthetized with thiopental i.p. (20 mg/kg) and killed by bleeding. Brains were removed and placed in Petri dishes containing oxygenated Krebs-Henseleit solution at room temperature. The MCAs were carefully removed from the brain, cleaned from surrounding tissues, and transferred to an organ chamber filled with Krebs-Henseleit solution, where two to four segments of around 5 mm in length were selected and isolated.
Following the method described by Halpern and Kelley(24), we cannulated each end of the segment with a 120-µm tapered glass micropipette, securely tying each end with 10-0 suture. Any small branches of the artery were tied off, and the segment was adjusted to its previous in vivo length. The cannulated segment was placed in an organ chamber filled with Krebs-Henseleit solution, which was maintained at 37°C, bubbled with a 95% O2-5% CO2 mixture, and continuously renewed using a peristaltic pump. TP was regulated with a servocontrolled syringe pump filled with Krebs-Henseleit solution and connected by a tube to a pressure transducer in line with the proximal micropipette. Arterial segments were pressurized to detect leaks, and leaky vessels were excluded. Then, segments were equilibrated at 30 mm Hg (resting pressure), which was equivalent to two-thirds of the mean arterial blood pressure recommended for this kind of in vitro experiment(24,25). During equilibration segments developed a spontaneous tone, usually within 60-90 min.
Measurement of segment diameter. The segment was transilluminated using a heat-filtered, 100-W tungsten light source. The resulting image was amplified using a microscope (Wild Heerbrugg, Switzerland) coupled to a videocamera (Sony CCDIris, Sony Corp., Japan) and displayed on a videomonitor. The internal diameter of the segment was measured with a videoscaler. Reference markers were used to ensure that measurements were repeated at the same point in the segment. Both internal diameter and TP were continuously recorded on a computer (Macintosh LCIII, Apple Computer Inc., CA) using the MacLab software (Chart version 3.3.8, AD Instruments Pty Ltd., Australia).
Protocol. After equilibration all segment were contracted with KCl (50 mM), and once the response plateaued, bradykinin (1 µM) was added. The integrity of the endothelium was verified when bradykinin induced a vasodilation equivalent to 75% of the previous KCl contraction. Endothelium-denuded segments were prepared by infusing intact segments with distilled water at 1 mL/min for 1 min, as described by Pearce et al.(26). Although this procedure produces a minimal trauma of segments during denudation, it may affect their autoregulatory properties(27). Therefore, six paired experiments were performed to compare the autoregulatory responses of segments whose endothelium was removed by infusing distilled water with those whose endothelium was removed by rubbing with a cotton-coated wire, and no significant differences were observed. After endothelium removal, the segment was allowed to develop spontaneous tone. Deendothelized segments were selected with resting diameters similar to those of intact segments to avoid attributing any differential response to pressure change to differences in initial segment diameter(4). The absence of endothelium was confirmed by the incapacity of bradykinin to induce vasodilation after precontraction with KCl. After bradykinin testing, all segments were further equilibrated for 30 min at resting pressure.
The following procedure was used to obtain the pressure curves. TP was decreased to 10 mm Hg, then increased to 70 mm Hg, and finally decreased to 30 mm Hg. Each pressure change maneuver was carried out in 20-mm Hg increments and lasted for 2-4 s, followed by a resting period to allow the vessel to become stabilized; this usually occurred within 30 s. The upper limit of 70 mm Hg was chosen because at higher pressures most segments dilated dramatically and did not recover their resting diameter at 30 mm Hg, suggesting that the segments were damaged. The lower limit of 10 mm Hg was chosen because at lower pressure segment collapse was frequent.
To study the role played by different endothelial factors on the segment response to pressure change, the pressure curve was repeated after incubating the segment with different drugs. To block cyclooxygenase, some segments were incubated with indomethacin at a concentration (10 µM) that has been shown to block the prostaglandin production in piglet cerebral arteries(6). To determine whether NO played a role in the pressure-induced response, other segments were treated with L-NAME at a concentration (10 µM) that has been shown to inhibit NOS in piglet cerebral arteries(11); in the L-NAME-treated segments, the pressure curve was performed in the presence of L-NAME alone and again after adding L-arginine (10 µM) to restore NO production. Other segments were incubated with 1 µM glibenclamide to block KATP channels; the receptor selectivity of this antagonist was assessed by obtaining a response to the KATP agonist cromakalim (1 µM) before and after gliben-clamide. Other segments were incubated with the KCa channel blocker TEA, at a concentration (1 mM) that has been shown to block KCa channels in adult cerebral arteries(16). Other segments were incubated with the ETA receptor blocker BQ-123 or with the ETB receptor blocker BQ-788, at concentrations (10 µM for BQ-123 and 10 nM for BQ-788), which counteracted the effect of ET-1 (10 nM). Some intact segments were contracted with 1 µM NE to analyze the effect of the increase in vascular tone on the response to pressure change. Drug solutions were administered extraluminally to the arterial segment, by infusing them into the organ chamber using a peristaltic pump. In selected segments, three successive control pressure curves were performed to demonstrate the reproductibility of the curves, ruling out the possibility that the effects observed in the presence of drugs were due to changes in the sensitivity of segments.
To verify the integrity of the segment at the end of the experiment, 120 mM KCl and then 100 µM papaverine were administered to strongly contract and dilate the vessel. If the contraction induced by 120 mM KCl was not greater than that caused by 50 mM KCl or the papaverine-induced vasodilation was not greater than 20% of resting diameter, the results obtained from the segment were excluded from analysis.
Solutions and drugs. The composition of Krebs-Henseleit solution (mM) was: NaCl 115, CaCl2 2.5, KCl 4.6, NaH2PO4 1.2, MgSO4·7H2O 1.2, NaHCO3 25, glucose 11.1, and Na2EDTA 0.03. A KCl (120 mM) solution was identical to Krebs-Henseleit solution except that NaCl was replaced by KCl on an equimolar basis. L-Arginine, bradykinin, NE, L-NAME, and TEA (Sigma Chemical Co., St. Louis, MO), and BQ-123 (Neosystems Laboratories, Montreal, Quebec, Canada), were dissolved in distilled water. Indomethacin (Sigma Chemical Co.) was dissolved in distilled water containing sodium carbonate (1.5 mM). Glibenclamide and BQ-788 (Sigma Chemical Co.) were dissolved in DMSO. Further drug dilutions were carried out in Krebs-Henseleit solution.
Data analysis. Segment resting diameter was measured just before the initial pressure curve. The change in diameter induced by drug incubation is expressed as a percentage of the resting segment diameter. Diameter variations during the subsequent pressure curves are expressed as a percentage of the segment diameter after drug incubation. All results are expressed as mean ± SEM. Statistical differences of drug-associated responses from control responses were studied by analysis of variance, with Scheffé's multiple range test, from a statistics package (Statview SE+ Graphics, Abacus Concepts, Berkeley, CA). Differences between two particular values were studied using the t test, with the Bonferroni's correction. A value of p less than 0.05 was considered significant.
RESULTS
Eighty-five MCA segments obtained from 55 piglets were used. The resting diameter was similar in segments with (240 ± 10 µm, n= 48) and without (234 ± 8 µm, n = 37) endothelium.
A representative model of response to pressure change in arterial segments with and without endothelium is illustrated in Figure 1. During TP decrease to 10 mm Hg intact segments vasodilated, but in 30 out of 48 arterial segments, the response was biphasic, starting with a small decrease in segment diameter followed by a long lasting vasodilation. The final diameter during TP decrease to 10 mm Hg was even greater than the resting diameter. During TP increase, intact segments vasoconstricted, the magnitude being pressure-dependent. Endothelium-denuded segments responded passively to pressure change. Occasionally, segments without endothelium showed a transient vasodilation(4-6 s) during pressure decrease to 10 mm Hg.

Representative model showing diameter variations of MCA segments (with, E+, or without, E-, endothelium) from the same piglet (4-d-old) during TP changes. Diameter line declining represents vasoconstriction, and line ascending represents vasodilation. The E+ segment was autoregulated over all pressure ranges, but not the E- segment.
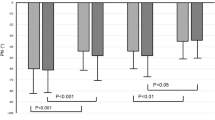
During the pressure curve, diameter variations of segments with endothelium were inverse to those of segments without endothelium(Fig. 2). However, the maximum dilator or constrictor response was close to 5% resting diameter in both types of segments. Segment resting diameter was recovered at the end of the pressure curve both in segments with and without endothelium.

Changes in diameter of MCA segments with(E+, n = 48) and without endothelium(E-, n = 37) from 3-4-d-old piglets during TP changes. Variations are expressed in percentage of diameter of segments after the equilibrium period at 30 mm Hg. Each symbol represents the mean ± SEM of 10 to 15 experiments. *p <0.05 by ANOVA.
During three successive control pressure curves, segment resting diameter remained unchanged (Fig. 3). Incubation with L-arginine(10 µM), glibenclamide (1 µM), BQ-123 (10 µM), BQ-788 (10 nM), or indomethacin (10 µM) did not modify the segment diameter. In contrast, L-NAME (10 µM) and TEA (1 mM) induced a contraction of about 20% of resting diameter. The magnitude of the vasocontraction induced by L-NAME and TEA was similar to that caused by NE (1 µM) (-18.7 ± 2.6, -20.7 ± 4.2, and -23.1 ± 3.6 µm, for L-NAME, TEA and NE, respectively). No drug used modified the resting diameter in endothelium-denuded segments (data not shown).

Effect of different drugs on resting diameter of intact MCA segments from 3-4-d-old piglets (n = 48). Control bar represents the resting diameter variation after three successive pressure curves in the absence of drugs. Results are expressed in percentage of resting diameter (mean = 240 ± 10 µm) and are mean ± SEM of 10 to 15 experiments. *p < 0.05 for differences with control by ANOVA. L-ARG, L-arginine; GLIB, glibenclamide;INDO, indomethacin.
The vasodilation during TP decrease to 10 mm HG in segments with endothelium was not modified by L-arginine, indomethacin, or BQ-788, but it was reduced by TEA (Fig. 4). Although glibenclamide was able to counteract the vasodilation evoked by the KATP channel activator cromakalim (34.3 ± 5.9 versus 4.8 ± 1.7% resting diameter for 1 µM cromakalim and cromakalim + glibenclamide, respectively, p < 0.05), this KATP blocker did not modify the pressure decrease-induced vasodilation. The presence of L-NAME prevented the arterial segment from recovering resting diameter during TP decrease to 10 mm Hg. As incubation with both TEA and L-NAME reduced the resting diameter, the segment response to pressure decrease to 10 mm Hg was studied in eight intact segments contracted with 1 µM NE. These segments showed a stronger dilation than did control segments (4.5 ± 0.9 versus 24.1 ± 7.8% resting diameter, in control and NE-contracted segments, respectively, p < 0.005).

Effect of different drugs on changes in diameter of intact MCA segments (n = 48) from 3-4-d-old piglets during TP decrease to 10 mm Hg. Variations are expressed in percentage of diameter of segments after the equilibrium period at 30 mm Hg. Each bar represents the mean ± SEM of 10-15 experiments. *p < 0.05 and **p < 0.01, by ANOVA. L-ARG, L-arginine;INDO, indomethacin; GLIB, glibenclamide.
To demonstrate that the vasodilation blockade by L-NAME was due to inhibition of NOS activity, the ability of L-arginine to revert the effect of L-NAME was investigated (Fig. 5). In intact segments, L-arginine partially reversed the L-NAME-induced decrease in resting diameter(-25.6 ± 5.9 versus -13.5 ± 5.2% resting diameter for L-NAME and L-NAME + L-arginine, respectively, p < 0.05), whereas in endothelium-denuded segments it did not (-11.6 ± 7.9versus -9.1 ± 5.1% resting diameter for L-NAME and L-NAME + L-arginine, respectively). In intact segments, L-arginine recovered the pressure decrease-induced vasodilation previously blocked by L-NAME. However, in endothelium-denuded segments neither L-NAME nor L-arginine plus L-NAME exerted any effect on segment response to pressure change.

Effect of L-NAME and L-arginine + L-NAME on changes in diameter of MCA segments, with (E+) or without (E-) endothelium, from 3-4-d-old piglets during TP decrease to 10 mm Hg. Experiments were paired, and drugs were sequentially added to each artery. Variations are expressed in percentage of diameter of segments after the equilibrium period at 30 mm Hg. Each bar represents the mean ± SEM of six experiments. *p < 0.05 by ANOVA for differences with control. †p < 0.05 by ANOVA for differences with E+. L-ARG, L-arginine.
The vasoconstriction during TP increase to 70 mm Hg in segments with endothelium was reversed by indomethacin and BQ-123 (Fig. 6). The magnitude of the response with BQ-123 was similar to that observed in segments without endothelium (-4.3 ± 0.6versus -6.6 ± 1.8% resting diameter for control endothelium-denuded segments and intact segments with BQ-123, respectively). Although BQ-788 inhibited the 10 nM ET-1-induced contraction as successfully as BQ-123 (-28.7 ± 6, -9,2 ± 3, and 4.1 ± 2.1% resting diameter for ET-1, ET-1 + BQ-123, and ET-1 + BQ-788, respectively, p < 0.05), the ETB-receptor antagonist did not modify the vasoconstrictor response during pressure increase.

Effect of different drugs on changes in diameter of intact MCA segments (n = 48) from 3-4-d-old piglets during TP increase to 70 mm Hg. Variations are expressed in percentage of diameter of segments after the equilibrium period at 30 mm Hg. Each bar represents the mean ± SEM of 10-15 experiments. *p < 0.05 and **p < 0.01, by ANOVA for differences from control. INDO, indomethacin.
Incubation with L-NAME and TEA produced as strong a reduction in the segmental resting diameter as that produced by 1 µM NE. In these vasoconstricted segments, TP increase to 70 mm Hg were unable to induce further contraction and a vasodilation was observed (2.4 ± 0.5, 9.4± 1.2, and 19.2 ± 9.4% resting diameter for L-NAME, TEA, and NE, respectively). No drug solution affected the response to pressure change in endothelium-denuded segments (data not shown).
DISCUSSION
Role of endothelium in piglet cerebral arterial autoregulation. The present study shows that the autoregulatory response to change in TP in newborn piglet MCA segments was endothelium-dependent. Similar results have been described in cat MCA(9). However, in piglet pial arterioles, the pressure decrease-induced vasodilation is dependent on endothelium, whereas pressure increase-induced vasoconstriction may only be myogenic(28). An inverse relationship between the strength of the myogenic reflex and vessel diameter exists(4). Thus, piglet pial arterioles (mean diameter = 173 ± 11 µm) dilate about 20% diameter during blood pressure decrease of 40%(29). In contrast, piglet MCA segment (mean diameter = 240 ± 10 µm) dilated about 5% diameter during intraluminal pressure decrease of 60%. This result is similar to that described in perfused cat MCA segments(23).
Pressure decrease-induced vasodilation. Pressure decrease to values near segment collapse vasodilated intact MCA segments to diameters similar, or greater, than resting diameter. In vivo experiments in piglets reveal that during hemorrhagic hypotension cerebrovascular resistances fall to obtain a cerebral blood flow higher than basal(2). This suggests a stronger tolerance to intravascular pressure decrease in newborn cerebral circulation than in adults, as described for piglets(30) and humans(1).
The role of NO in pressure decrease-induced vasodilation of cerebral vessels is controversial. Administration of NOS inhibitors induces an increase in cerebral vessel tone as much in vivo(22) as in vitro(5,31), suggesting that NO has a tonic vasorelaxant influence on such vessels. We recently demonstrated the involvement of NO in the endothelium-dependent vasodilation of piglet cerebral arteries(31). In our present study, incubation with L-NAME not only reduced the MCA resting diameter by 20%, but it also eliminated the pressure decrease-induced vasodilation. The effect of L-NAME was counteracted by the NOS substrate, L-arginine, suggesting that NO mediated such a vasodilation in piglet MCA segments. Previous in vivo experiments performed in rats(18,19) and piglets(14) failed to demonstrate that NOS inhibitors impair cerebrovascular autoregulation during hypotension. However, in these studies, the concentration of(18,19) or the incubation time with(14) the NOS inhibitors may have been less than recommended(32). Indeed, when these variables are adjusted, NOS inhibitors successfully impair cerebral autoregulation during severe hemorrhagic hypotension in piglets(20), rats(21), and cats(22). Incubation with L-arginine had no effect on either resting diameter or pressure decrease-induced vasodilation, suggesting that the availability of L-arginine was not a limiting factor for NO production in cerebral arteries in our experiment. Similar results have been found in adult animals(5).
In our experiment, incubation with the KCa channel blocker TEA reduced the resting diameter of piglet MCA segments by 20%. In rabbit cerebral arteries, KCa channel blockers reduced resting diameter by 40%(33). These results suggest that Kca channels participate in the control of cerebral arterial tone. During TP decrease to 10 mm Hg, TEA reduced the vasodilation, suggesting a role for Kca channels in this response. Activation of Kca channels leads to vasodilation in rabbit cerebral arteries by increasing the membrane potential(33), a mechanism of particular importance in cerebral arteries(34). There is evidence that the activation of KCa channels mediate the vasodilation caused by NO in systemic(35) and cerebral(36) adult arteries. We observed that the effect of L-NAME on the TP decrease-induced vasodilation of intact segments was similar to that of TEA, and that TEA did not show any effect on segments without endothelium. This suggests that Kca channels might mediate the NO-induced vasodilation during pressure decrease in intact piglet MCA segments.
When intact piglet MCA segments were contracted with NE the vasodilator response at 10 mm Hg of TP was enhanced, suggesting that the level of precontraction of segments positively influences the sensitivity to pressure change. The magnitude of the contraction caused by NE was similar to that induced by the incubation with L-NAME and TEA. Therefore, the effect of L-NAME and TEA on the segment vasodilation during TP decrease could not be attributed to the vascular tone increase caused by the these drugs.
KATP channels, present in piglet pial arterioles, contribute to cGMP-induced vasodilation(37). The KATP channel activator cromakalim exerted a vasodilator effect in piglet MCA segments, which was abolished by glibenclamide, suggesting that KATP channels are also present in such arteries. KATP channel blockade with glibenclamide impairs autoregulation during hypotension in rats(38). Nevertheless, in our experiments, glibenclamide did not affect pressure decrease-induced vasodilation, suggesting that KATP channels are not involved in piglet MCA autoregulation.
Prostanoids are important in the control of cerebral blood flow in newborn piglets(2,13). However, the cyclooxygenase inhibitor indomethacin did not increase vascular tone in intact piglet MCA segments. This vasoconstrictor effect is observed in piglet pial arterioles(2) and in perfused cat MCA segments(23). Indomethacin did not modify the TP decrease-induced vasodilation, suggesting that prostanoids were not involved in this vasodilator response. It is known that in piglets, prostanoid concentration in sagittal sinus blood increases after an hypotensive insult(13) and that indomethacin reverses the pial arteriolar dilation shown during hypotension(39). This suggests that although prostanoids contribute to the maintenance of cerebral blood flow during hypotension in piglets, their effect is manifested in other cerebral vessels but not in MCA.
The vasoconstriction induced by ETs in cerebral arteries is mediated by ETA receptors, whereas vasodilation is mediated by ETB receptors(15). In our case, BQ-123 and BQ-788, ETA and ETB receptor antagonists, respectively(40), did not affect resting diameter nor did they reduce the vasodilation during TP fall to 10 mm Hg. This suggests that ETs did not participate in the pressure decrease-induced vasodilation in piglet MCA.
Pressure increase-induced vasoconstriction. Indomethacin abolished the vasoconstriction during pressure increase to 70 mm Hg. A similar effect, thought to be due, in part, to an increase in basal tone caused by indomethacin, has been found in perfused cat MCA(23). In our experiment, however, incubation with indomethacin did not affect the resting diameter, suggesting that the inhibition of the pressure increase-induced vasoconstriction by indomethacin was only due to the prostanoid production blockade. In vivo experiments on newborn piglets show that after indomethacin pretreatment, cerebrovascular resistance increases as blood pressure rises(13). This difference between in vivo and in vitro results may occur because in vivo, indomethacin acts on cerebral vessels by mechanisms other than cyclooxygenase inhibition(23).
After incubation with the ETA receptor blocker BQ-123, piglet MCA segments did not contract but did dilate during pressure increase to 70 mm Hg. In fact, the effect of BQ-123 was similar in character and magnitude to endothelium removal, suggesting that ETs played a chief role in the pressure increase-induced vasoconstriction in our in vitro model. ET-1 contracts cerebral vessels in adult(15) and newborn animals(41), this contraction being mediated by ETA receptors(15). In piglet MCA segments the contraction caused by ET-1 was reversed by both BQ-123 and BQ-788, suggesting that this contraction was mediated by ETA and ETB receptors. However, pressure increase-induced vasoconstriction was inhibited by BQ-123 but not by BQ-788, suggesting that only ETA receptors mediated the ET-induced vasoconstriction during pressure increase. It has been reported that prostanoids may be involved in the vasoconstriction induced by ET-1 in piglet pial arterioles(41). Our results agree with this suggestion because indomethacin, which did not modify the pressure decrease-induced vasodilation, exerted an effect similar to BQ-123 on pressure increase-induced vasoconstriction.
L-NAME and TEA abolished the constrictor response to pressure increase. This was attributable to their effect on vascular tone. When a similar level of vasoconstriction was achieved with NE, intact segments were also unable to contract during pressure increase. It has been reported that potassium and phenylephrine eliminate the constrictor response during pressure increase in hamster cheek arterioles(4), suggesting that a high initial level of constriction could mask the myogenic response.
In conclusion, piglet MCA autoregulation seems to be endothelium-dependent. Intact segments were able to dilate during pressure decreases to values close to vessel collapse, reflecting a particular ability of newborn cerebral arteries to tolerate hypotension. Endothelial NO was involved in this vasodilation. The effect of NO seems to be mediated by Kca channels. Intact segments contracted during pressure increases. ET-1 participated in this vasoconstriction by activating ETA receptors. Prostanoids are also involved in this contraction. Therefore, endothelial impairment may be of particular importance in the pathophysiology of conditions associated with a loss of cerebral autoregulation in newborns.
Abbreviations
- ET:
-
endothelin
- KATP:
-
ATP-sensitive potassium
- KCa:
-
calcium-activated potassium
- L-NAME:
-
NG-nitro-L-arginine methylester
- MCA:
-
middle cerebral artery
- NE:
-
norepinephrine
- NO:
-
nitric oxide
- NOS:
-
NO synthase
- TEA:
-
tetraethylammonium
- TP:
-
transmural pressure
References
Volpe JJ 1989 Intraventricular hemorrhage and brain injury in the premature infant. Neuropathology and pathogenesis. Clin Perinatol 16: 361–386
Leffler CW, Busija DW, Beasley DG 1987 Effect of therapeutic dose of indomethacin on the cerebral circulation of newborn pigs. Pediatr Res 21: 188–192
Kontos HA, Wei EP, Navari RM, Levasseur JE, Rosenblum WI, Patterson JL 1978 Response of cerebral arteries and arterioles to acute hypotension and hypertension. Am J Physiol 234:H371–H383
Jackson PA, Duling BR 1989 Myogenic responses and wall mechanics of arterioles. Am J Physiol 257:H1147–H1155
Faraci FM 1991 Role of endothelium-derived relaxing factor in cerebral circulation: large arteries vs. microcirculation. Am J Physiol 261:H1038–H1042
Shimoda LA, Norins NA, Jeutter DC, Madden JA 1996 Flow-induced responses in piglet isolated cerebral arteries. Pediatr Res 39: 574–583
Tuor UI, Grewal D 1994 Autoregulation of cerebral blood flow: influence of local brain development and postnatal age. Am J Physiol 267:H2220–H2228
García-Roldán JL, Bevan JA 1990 Flow-induced constriction and dilation of cerebral resistance arteries. Circ Res 66: 1445–1448
Harder DR, Sánchez-Ferrer CF, Kauser K, Stekiel WJ, Rubanyi GM 1989 Pressure releases a transferable endothelial contractile factor in cat cerebral arteries. Circ Res 65: 193–198
Pearce WJ, Hull AD, Long DM, Longo LD 1991 Developmental changes in ovine cerebral artery composition and reactivity. Am J Physiol 261:R458–R465
Martínez-Orgado J, Salaices M, Rodríguez-Martínez MA, Sánchez-Ferrer CF, Marín J 1995 Age-related changes in nitric oxide-mediated vasodilation of cerebral arteries from newborn piglets. Pharm Sci 1: 35–37
Pearce WJ, Hull AD, Long DM, White CR 1994 Effects of maturation on cyclic GMP-dependent vasodilation in ovine basilar and carotid arteries. Pediatr Res 36: 25–33
Chemtob S, Beharry K, Rex J, Varna DR, Aranda JV 1990 Prostanoids determine the range of cerebral blood flow autoregulation in newborn piglets. Stroke 21: 777–784
Meadow W, Rudinsky B, Bell A, Lozon M, Randle C, Hipps R 1994 The role of prostaglandins and endothelium-derived relaxing factor in the regulation of cerebral blood flow and cerebral oxygen utilization in the piglet: operationalizing the concept of essential circulation. Pediatr Res 35: 649–656
Patel TR, McAuley MA, McCulloch J 1996 Endothelin receptor mediated vasoconstriction and dilation in feline cerebral arteriolesin vivo. Eur J Pharmacol 20: 41–48
Kitazono T, Faraci FM, Taguchi H, Heistad DD 1995 Role of potassium channels in cerebral blood vessels. Stroke 26: 1713–1723
Iadecola C, Pelligrino DA, Moskowitz MA, Lassen NA 1994 Nitric oxide synthase inhibition and cerebrovascular regulation. J Cereb Blood Flow Metab 14: 175–192
Buchanan JE, Phillis JW 1993 The role of nitric oxide in the regulation of cerebral blood flow. Brain Res 610: 248–255
Wang Q, Paulson OB, Lassen NA 1992 Is autoregulation of cerebral blood flow in rats influenced by nitro-L-arginine, a blocker of the synthesis of nitric oxide?. Acta Physiol Scand 145: 297–298
O'Neill JT, Sogn A, Hunt T, Palacino J 1995 Inhibition on nitric oxide synthase alters the cerebral blood flow response to hemorrhagic hypotension in piglets. Pediatr Res 37: 227A
Tanaka K, Fukuuchi Y, Gomi S, Mihara B, Shirai T, Nogawa S, Nozaki H, Nagata E 1993 Inhibition of nitric oxide synthesis impairs autoregulation of cerebral blood flow in the rat. Neuroreport 4: 267–270
Kobari M, Fukuuchi Y, Tomita M, Tanahashi N, Takeda H 1994 Role of nitric oxide in regulation of cerebral microvascular tone and autoregulation of cerebral blood flow in cats. Brain Res 667: 255–262
Norins NA, Wendelberger K, Hoffman RG, Keller PA, Madden JA 1992 Effects of indomethacin on myogenic contractile activation and responses to changes in O2 and CO2 in isolated feline cerebral arteries. J Cereb Blood Flow Metab 12: 866–872
Halpern W, Kelley M 1991 In vitro methodology for resistance arteries. Blood Vessels 28: 245–251
Norins NA, Madden JA 1992 Developmental changes in cerebral artery autoregulation in piglets. Pediatr Res 31: 214A
Pearce WJ, Ashwall S, Cuevas J 1989 Direct effects of graded hypoxia on intact and denuded rabbit cranial arteries. Am J Physiol 257:H824–H833
Liu Y, Harder DR, Lombard JH 1994 Myogenic activation of canine small renal arteries after nonchemical removal of endothelium. Am J Physiol 267:H302–H307
Eidson TH, Edrington JL, Albuquerque MLC, Zuckerman SL, Leffler CW 1995 Light/dye microvascular injury eliminates pial arteriolar dilation in hypotensive piglets. Pediatr Res 37: 10–14
Busija DW, Leffler CW 1991 Selective attenuation by perivascular blood of prostanoid-dependent cerebrovascular dilation in piglets. Stroke 22: 484–488
Harada J, Takaku A, Endo S, Kuwayama N, Fukuda O 1991 Differences in critical cerebral blood flow with age in swine. J Neurosurg 75: 103–107
Martínez-Orgado J, Salaices M, Rodríguez-Martínez MA, Sánchez-Ferrer CF, Marín J 1994 Role of nitric oxide on the endothelium-dependent vasodilation in newborn piglet cerebral arteries. Gen Pharmacol 25: 899–902
Traystman RJ, Moore LE, Helfaer MA, Davis S, Banasiak K, Williams M, Hurn P 1995 Nitro-L-arginine analogues. Dose- and time-related nitric oxide synthase inhibition in brain. Stroke 26: 864–869
Brayden JE, Nelson MT 1992 Regulation of arterial tone by activation of calcium-dependent potassium channels. Science 256: 532–535
Brayden JE 1990 Membrane hyperpolarization is a mechanism of endothelium-dependent cerebral vasodilation. Am J Physiol 259:H668–H673
Khan SA, Mathews R, Meisheri KD 1993 Role of calcium-activated K+ channels in vasodilation induced by nitroglycerine, acetylcholine and nitric oxide. J Pharmacol Exp Ther 267: 1327–1335
Robertson BE, Schubert R, Hescheler J, Nelson MT 1993 c-GMP-dependent protein kinase activates Ca-activated K channels in cerebral artery smooth muscle cells Am J Physiol 265:C299–C303
Armstead WM 1996 Role of ATP-sensitive K+ channels in cGMP-mediated pial artery vasodilation. Am J Physiol 270:H423–H426
Lee WS, Kwon Y, Yu SS, Rhim BY, Hong KW 1993 Disturbances in autoregulatory responses of rat pial arteries by sulfonylureas. Life Sci 52: 1527–1534
Leffler CW, Busija DW 1987 Prostanoids and pial arteriolar diameter in hypotensive newborn pigs. Am J Physiol 252:H687–H691
Karaki H, Sudjarwo SA, Hori M 1994 Novel antagonist of endothelin ETB1 and ETB2 receptors BQ-788: effects on blood vessel and small intestine. Biochem Biophys Res Commun 205: 168–173
Armstead WM, Mirro R, Leffler CW, Busija DW 1989 Influence of endothelin on piglet cerebral microcirculation. Am J Physiol 257:H707–H710
Acknowledgements
The authors thank George F. R. Marsden for correction and critical evaluation of the manuscript.
Author information
Authors and Affiliations
Additional information
Supported by grants from Direccion General de Industria, Ciencia y Technologia (PB94-0152), FIS (95/1954), Comunidad de Madrid (I+D 0017/94 and AE 00339/95), and Bayer España. This work is ascribed to Biomed Project No. BMH1-CT94-1375 of the European Economic Community.
Rights and permissions
About this article
Cite this article
Martínez-Orgado, J., González, R., Alonso, M. et al. Endothelial Factors and Autoregulation during Pressure Changes in Isolated Newborn Piglet Cerebral Arteries. Pediatr Res 44, 161–167 (1998). https://doi.org/10.1203/00006450-199808000-00004
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-199808000-00004
This article is cited by
-
Cannabidiol reduces lipopolysaccharide-induced vascular changes and inflammation in the mouse brain: an intravital microscopy study
Journal of Neuroinflammation (2011)
