
Abstract
Because the antiprotease defense in cystic fibrosis (CF) airways is overwhelmed by neutrophil elastase (NE), substitution of antiproteases such as secretory leukoprotease inhibitor (SLPI) seems to be a reasonable therapeutic approach. Knowing, however, that native antiproteases may be liable to rapid inactivation by the locally abundant oxidants, we comparatively investigated the interactions of CF sputum with recombinant native SLPI (rSLPI) and its partially oxidation-resistant variant (rSLPI-242), respectively, to estimate their therapeutic potentials. NE activity in supernatants from diluted CF sputum samples was dose-dependently inhibited by both rSLPI and rSLPI-242, with comparable potency. Addition of the oxidant N-chlorosuccinimide resulted in significant superiority of rSLPI-242 over rSLPI. When fresh neutrophil-rich CF sputum was incubated with rSLPI and rSLPI-242, respectively, rSLPI-242 inhibited sputum NE activity significantly more potently than did rSLPI; addition of the antioxidant superoxide dismutase significantly improved the effect of rSLPI. Furthermore, secretion of radiolabeled macromolecules from porcine tracheal glands induced by purified NE or by CF sputum was inhibited dose-dependently by rSLPI and even better by rSLPI-242. We conclude that both rSLPI and rSLPI-242 effectively inhibit NE activity and NE-induced gland hypersecretion in vitro. In vivo effects in CF remain to be analyzed; an advantage of the partially oxidation-resistant rSLPI-242 can be expected.
Similar content being viewed by others

Main
Bronchopulmonary disease in CF is characterized by neutrophil-dominated chronic inflammation. Sputum from patients with CF contains large numbers of neutrophils and high concentrations of catalytically active neutrophil-derived proteases(1–4). Prominent among these is NE (EC 3.4.21.37), which is released from neutrophils during cell activation, phagocytosis, and cell death(5, 6). NE has been implicated in a variety of conditions associated with CF lung pathophysiology such as destruction of lung connective tissue(7), impairment of host defense mechanisms(8, 9), and stimulation of airway gland secretion(10–12).
In healthy subjects, there are sufficient amounts of antiproteases present to protect the airways against NE; the protective shield of the lung against NE includes α1PI and SLPI. SLPI, a 12-kD, nonglycosylated, disulfide-linked antiprotease, is secreted by cells of mucosal surfaces including the epithelium of the airways, and it provides the major anti-NE protection of the large airways(13–16).
In CF airways, the endogenous anti-NE defense is over-whelmed: bothα1PI and SLPI are complexed with NE, proteolytically cleaved, or inactivated by oxidants released from inflammatory cells(2). The major imbalance between the large NE burden and the ineffective anti-NE defense is likely to be a key factor in progressive destruction of lung matrix and hypersecretion present in CF airways(7, 12). Hence, substitution of active antiproteases, e.g. rSLPI, to CF airways by aerosolization appears to be a reasonable therapeutic approach to bind and inactivate NE. However, also exogenously administered antiproteases may undergo rapid oxidative inactivation in the airways. Therefore, a variant of rSLPI with relative oxidation resistance (rSLPI-242) was constructed using recombinant DNA technology(17).
We designed the present study as an in vitro investigation to estimate the therapeutic potentials of rSLPI and rSLPI-242 as antiprotease replacement therapy for CF patients' airways. Our data show that both rSLPI and rSLPI-242 dose-dependently inhibit NE activity in CF sputum as well as airway gland secretion induced by sputum from CF patients. Due to its relative oxidation resistance, rSLPI-242 proved to be superior to rSLPI regarding inhibition of both NE activity and the secretagogue effect of CF sputum.
METHODS
Subjects and sputum samples. Sputum samples were collected from 26 patients with CF in clinically stable conditions after obtaining informed consent. The diagnosis of CF had been established using generally accepted criteria(18). The patients were 5-30-y-old (mean± SEM, 15.5 ± 1.8 y; 18 female, 8 male); in 16 of them, there was chronic airway colonization with Pseudomonas aeruginosa. As controls, sputum samples from 10 healthy nonsmoking volunteers (mean age± SEM, 26.4 ± 1.1 y; 7 female, 3 male) were collected after a 20-min inhalation of a 5% NaCl solution.
After expectoration, sputum samples were cooled at 4 °C and processed within 30 min. The viscous samples were diluted 1:3 with PBS (pH 7.4), agitated for 1 min on a vortex-mixer, and subsequently centrifuged for 20 min at 8800 × g to remove insoluble material. The supernatants were collected and stored at -70 °C until analyzed.
Six additional CF sputum samples were processed in a different way. Immediately after expectoration and 1:3 dilution, they were used for NE activity measurements after incubation with rSLPI and rSLPI-242. In contrast to the deep-frozen supernatants decribed above, there were great numbers of neutrophils present in these fresh sputum samples during the incubation period.
Neutrophil elastase activity. NE activities in the sputum samples were measured photometrically using the specific chromogenic peptide substrate methoxy-succinyl-L-alanyl-L-alanyl-L-prolyl-L-valine-p-nitroanilide(Sigma Chemical Co., Deisenhofen, FRG). The sputum supernatants were diluted in a buffer (pH 7.5) containing 0.1 M HEPES, 0.5 M NaCl, 0.1% Brij-35; then, 50 μL each were added to 100 μL of the chromogenic substrate dissolved in 1-methyl-2-pyrrolidinone (1 mM). The absorbance was recorded at 405 nm in a spectrophotometer (Titertek Multiscan Plus Mk11, ICN, Meckenheim, FRG). Purified human NE (Elastin Products Co., Pacific, MO) was used as a standard reference. One enzyme unit was defined as the release of 1 μmol ofp- nitroaniline/min at 24 °C(2). The lower detection limit of the assay is in the range of 0.0002 U/L.
To determine the effects of exogenous rSLPI and rSLPI-242 on NE activity in CF sputum supernatants (n = 8), 25 μL of the samples were preincubated at 24 °C for 30 min, either with 25 μL of buffer alone or with 25 μL of buffer containing either one of the inhibitors at different concentrations (0.003-10 μM) before addition of the substrate. To evaluate the effect of an oxidant, incubations were carried out in the presence and in the absence of N-chlorosuccinimide (4 mM; i.e. a molar ratio of ≥400:1, oxidant equivalents to SLPI).
In addition to the experiments with the neutrophil-free sputum supernatants, aliquots of six fresh CF sputum samples were incubated with rSLPI and rSLPI-242 (3 μM), respectively, immediately after expectoration and dilution of the samples; subsequently, NE activities were measured as described above. To evaluate the effect of oxidants presumably released from neutrophils in these fresh sputum samples, incubations were carried out in the presence and in the absence of the antioxidant SOD (400 U/mL).
Preparation of submucosal glands from porcine tracheas. Secretion studies were carried out with porcine tracheal explants which are considered a good model for human airways regarding secretion(19). To study secretion derived exclusively from submucosal glands, the epithelium was removed. Intercartilaginous areas containing the submucosal glands were dissected, transferred into a beaker containing nutrient mixture F-12 (Ham's) medium, placed in a Dubnoff shaker, and provided with 95% O2 and 5% CO2 at 37 °C.
Release of 35S-labeled macromolecules from porcine tracheal submucosal glands. The principle of the experimental procedure has been described previously(11). In brief, intercartilaginous tissue pieces were incubated in 20 mL of nutrient mixture F-12 (Ham's) medium containing Na235SO4 (20 μCi/mL) to allow radiolabeling of mucus glycoconjugates. After 2 h of incubation, the medium containing excess radioisotope was removed, and the tissues were washed 10 times with PBS. Next, the tissue pieces were incubated in vials containing 2 mL of fresh medium. After 120 min of rest, the medium was collected and replaced every 30 min. At 210 min, the medium was replaced either by 1.8 mL of fresh medium alone or-for inhibition studies-with 1.8 mL of medium containing different concentrations of rSLPI and rSLPI-242 (0.001-5 μM), respectively, to allow for a 5-min preincubation of the tissues with the inhibitors. At 215 min, 0.2 mL either of medium alone (baseline control), or purified NE (final concentration, 1 μM;n ≥ 10 for each inhibitor concentration), or sputum supernatants(final dilution, 1:30; n ≥ 12 for each inhibitor concentration) were added. To evaluate a possible effect of tissue-derived oxidants on inhibitor activities, we also evaluated inhibition of NE (1 μM)-induced secretion by rSLPI and rSLPI-242 (0.3 μM), respectively, in the presence and in the absence of SOD (400 U/mL; n = 10 each). At 240 min, the medium was collected again. The harvested samples were dialyzed exhaustively against distilled water to remove unincorporated 35SO42-(dialysis tubing, molecular mass cutoff 12-14 kD, Medicell International Ltd., London, GB). Nondialyzable 35S-labeled macromolecules in the samples were counted after addition of 10 mL of scintillation fluid (Ready Safe, Beckman Instruments, Munich, FRG) in a scintillation counter (LS3801, Beckman Instruments). Secretion is expressed as percent increase of release of35 S-labeled macromolecules during incubation with the agonists ± inhibitor (min 210-240) over the release during the preceding time period (min 180-210). The values were corrected for the slightly declining baseline as determined in the controls incubated with medium alone. The threshold concentration for NE to induce secretion in this experimental setting is 10-8 M.
Assay of cytotoxicity. Possible cytotoxicity of purified NE and sputum supernatants was determined by measuring the release of LDH from unlabeled porcine tracheal tissues into the medium. The tissues were incubated for 60 min in nutrient mixture F-12 (Ham's) medium (2 mL/tissue piece). The medium was then replaced by fresh medium alone (negative control), medium containing 1% Triton-X-100 (positive control), medium containing NE (1 μM), or sputum supernatants (final dilution, 1:30) (n = 6 each). After 60 min of incubation at 37 °C, LDH activity was measured using a commercially available kit (Sigma Chemical Co., Deisenhofen, FRG).
Statistical analysis. Data are expressed as the arithmetic mean± SEM. IC50 values were determined by nonlinear curve-fitting analysis (Enzfitter, Biosoft, Cambridge, GB). Statistical analysis was performed by the Mann Whitney rank sum test or the Wilcoxon signed-ranks test, as applicable(20). p values of <0.05 were considered significant.
RESULTS
Inhibition of NE activity in CF sputum by rSLPI and rSLPI-242. Mean activity of NE in the CF sputum samples as determined from measurements in the thawed supernatants (n = 26) was 5.63 ± 0.70 U/L(range, 1.11-15.84 U/L), corresponding to 3.2 ± 0.4 μM (range, 0.63-9.00 μM) of catalytically active NE, whereas there was no NE activity measurable in the control sputum samples from healthy subjects.
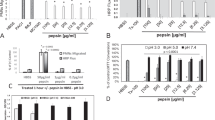
Incubation of CF sputum supernatants (final sputum dilution, 1:6;n = 8) for 30 min with different concentrations of rSLPI and rSLPI-242 (0.003-10 μM), respectively, resulted in dose-dependent inhibition of NE activity (Fig. 1). There was no significant difference between rSLPI and rSLPI-242 in their potency to inhibit NE activity in these CF sputum supernatants; complete (≥95%) inhibition of NE activity was achieved at concentrations ≥1 μM of either inhibitor(Fig. 1A). When N-chlorosuccinimide (4 mM) was added during the incubation period, both dose-response curves were shifted to the right. In the presence of the oxidant, however, inhibition of CF sputum NE activity was significantly higher by the partially oxidation-resistant rSLPI-242 (at ≥1 μM) compared with equal concentrations of rSLPI(p < 0.001; Fig. 1B).

Inhibition on NE activity in CF sputum supernatants by rSLPI (solid circles) and rSLPI-242 (open circles) in the absence (A) and in the presence (B) of the oxidantN- chlorosuccinimide (4 mM). Incubation of CF sputum supernatants with rSLPI and rSLPI-242, respectively, for 30 min at 24 °C resulted in dose-dependent inhibition of sputum NE activity. In the presence ofN- chlorosuccinimide, inhibition of NE activity by rSLPI-242 (≥1μM) was significantly higher than inhibition by equal concentrations of rSLPI (*p < 0.001). Values are mean ± SEM;n = 8 identical CF sputum samples for each data point.
However, when freshly expectorated, neutrophil-rich CF sputum samples were immediately incubated with rSLPI and rSLPI-242 (3 μM; n = 6), respectively, inhibition of sputum NE activity by rSLPI-242 was significantly higher than inhibition by rSLPI (p < 0.01; Fig. 2). The NE inhibitory activity of rSLPI-242 on fresh CF sputum lay in the same order of magnitude as its NE inhibitory activity on CF sputum supernatant, whereas inhibition of NE activity by rSLPI was significantly lower in fresh sputum compared with sputum supernatants. Addition of the antioxidant SOD (400 U/mL) during the incubation period significantly improved inhibition of sputum NE activity by rSLPI (p < 0.02, rSLPI plus SOD versus rSLPI without SOD), whereas in had no significant effect on NE inhibition by rSLPI-242. In the control experiments, SOD alone was shown to have no effect on sputum NE activity.

Effect of the presence of neutrophils on inhibition of NE activity in CF sputum by rSLPI (black columns) and rSLPI-242(white columns), 3 μM each. When neutrophil-free sputum supernatants (-PMN; n = 8 each) were assayed, there was no significant difference in NE inhibition by rSLPI compared with rSLPI-242. When neutrophil-rich sputum (+PMN) was incubated with the respective inhibitors immediately after expectoration, inhibition of NE activity by the relatively oxidation-resistant rSLPI-242 was significantly higher than inhibition by rSLPI (*p < 0.01); addition of an antioxidant (SOD) during the incubation period (+PMN +SOD) significantly improved inhibition of sputum NE activity by rSLPI(+p < 0.02; n = 6 each). Values are mean± SEM.
Inhibition of NE-induced secretion of 35S-labeled macromolecules from porcine tracheal submucosal glands by rSLPI and rSLPI-242
Purified NE (1 μM) stimulated secretion of 35S-labeled macromolecules from porcine tracheal submucosal glands to 1499 ± 347% above baseline (Fig. 3). Release of radiolabeled macromolecules was not due to a cytotoxic effect of NE; even during a 60-min exposure period, 1 μM NE did not cause a significant LDH release from tracheal tissues (47 ± 47 U/mL for NE, 27 ± 20 U/mL for medium alone, 1755 ± 33 U/mL for 1% Triton-X-100; n = 6 each). Incubation of the tracheal tissues with different concentrations of either rSLPI or rSLPI-242 (0.05-5 μM; n ≥ 10 each) resulted in dose-dependent inhibition of NE-induced secretion (Fig. 3); the inhibitors alone did not have any effect on secretion. As shown in the dose-response curves, the inhibitory effects of rSLPI-242 were significantly higher compared with rSLPI (p < 0.05). IC50 was 0.28 μM for rSLPI and 0.16 μM for rSLPI-242. When the antioxidant SOD (400 U/mL) was added during the incubation period, the difference between rSLPI and rSLPI-242 (0.3 μM each) in their potency to inhibit NE-induced secretion was abrogated (n = 10; p < 0.05;Fig. 4).

Inhibitory effects of rSLPI (solid circles) and rSLPI-242 (open circles) on NE-induced secretion (hatched column) of 35S-labeled macromolecules from porcine tracheal tissues. Both rSLPI and rSLPI-242 inhibited secretion induced by purified NE (1 μM) in a dose-dependent fashion. Inhibition by rSLPI-242 was significantly higher than inhibition by rSLPI (*p < 0.05). Values are mean± SEM; n ≥ 10 for each data point.

Effect of SOD on inhibition of NE (1 μM)-induced secretion from porcine tracheal tissues by rSLPI (black columns) and rSLPI-242 (white columns) (0.3 μM each). Addition of the antioxidant (+SOD; n = 10 each) resulted in abrogation of the difference between the inhibitory effects of rSLPI and rSLPI-242. Values are mean ± SEM; *p < 0.05.
Inhibition of CF sputum-induced secretion of35S-labeled macromolecules from porcine tracheal submucosal glands by rSLPI and rSLPI-242. Supernatants from CF sputum samples (final sputum dilution, 1:30) markedly stimulated secretion of35 S-labeled macromolecules from porcine tracheal submucosal glands with a mean secretory response of 607.7 ± 134.0% above baseline. Release of radiolabeled macromolecules was not due to a cytotoxic effect of the sputum samples; even during a 60-min exposure period, incubation with CF sputum did not cause a significant LDH release from tracheal tissues (LDH concentrations in medium, 106 ± 80 U/mL for 1:30 diluted sputum samples alone, 196± 129 U/mL for tracheal tissues incubated with CF sputum at a final concentration of 1:30, and 1755 ± 33 U/mL for 1% Triton X-100;n = 6 each). Incubation of the tracheal tissues with different concentrations of either rSLPI or rSLPI-242 (0.001-0.3 μM) resulted in dose-dependent inhibition of CF sputum-induced secretion (n ≥ 12 observations with identical CF sputum samples each; Fig. 5). Again, the inhibitory effect of the relatively oxidation-resistant rSLPI-242 was significantly higher than that of rSLPI. At 0.01 μM inhibitor concentrations, inhibition of CF sputum-induced secretion by rSLPI-242 was 84.8 ± 5.1% compared with 60.8 ± 10.1% inhibition by rSLPI(p < 0.05). Inhibition of >90% of CF sputum-induced secretion was achieved at 0.3 μM of either rSLPI or rSLPI-242. In contrast to CF sputum, sputum from healthy controls did not induce any secretory response from the porcine tracheal tissues (-6.2 ± 16.7% above baseline;n = 10).

Inhibitory effects of rSLPI (black columns) and rSLPI-242 (white columns) on secretion of 35S-labeled macromolecules from porcine tracheal tissues induced by CF sputum supernatants(hatched column). Both rSLPI and rSLPI-242 (0.001-0.3 μM) inhibited secretion induced by CF sputum supernatants in a dose-dependent fashion. At 0.01 μM, inhibition by the relatively oxidation-resistant rSLPI-242 was significantly higher than inhibition by rSLPI(*p < 0.05). Values are mean ± SEM; n≥ 12 observations with identical CF sputum samples each.
DISCUSSION
This study focused on inhibition of NE activities in CF sputum by rSLPI compared with its variant rSLPI-242. Both substances proved to inhibit cleavage of a NE-specific peptide substrate by CF sputum as well as secretagogue activity of CF sputum in a dose-dependent fashion. Our results indicate that rSLPI-242, due to its relative oxidation resistance, is superior to rSLPI as an inhibitor of NE activity in CF sputum.
High NE activities in bronchopulmonary secretions are considered to play an important role in the pathogenesis of pulmonary deterioration in CF(21, 22). Our measurements of NE activities in the supernatants of CF sputum samples confirm the results of earlier studies that demonstrated high concentrations of catalytically active NE in CF sputum(1–3). Various mechanisms by which the natural anti-NE screen becomes inactivated in the CF lung are feasible, including proteolysis of the inhibitors by overwhelming activities of NE itself(23), and oxidation of the inhibitors by the effects of factors released from neutrophils, such as superoxide anion and myeloperoxidase, which have been demonstrated to be present in high concentrations in CF sputum(2, 24, 25). It is known that the SLPI molecule is sensitive to oxidative inactivation due to its methionine residues(17), and in fact, there is evidence that inactivation of SLPI in CF airways is due to exposure to a heavy burden of both NE and oxidants(26).
In our present study, we compared the interactions of CF sputum with native rSLPI to those with rSLPI-242. The variant rSLPI-242 has been constructed by recombinant DNA technology via site-directed mutagenesis where methionine in the active center was replaced by leucine to achieve relative oxidation resistance of the molecule(17). Under standardin vitro conditions, native rSLPI and rSLPI-242 have been shown to have similar inhibitory characteristics toward NE on a molar basis(17). We hypothesized that rSLPI-242 might be advantageous over rSLPI as a therapeutic agent for anti-NE augmentation therapy in CF. Before a clinical study with aerosolized SLPI in CF patients, it was our aim to estimate and compare the therapeutic potentials of rSLPI and rSLPI-242 in vitro. In our first experiments with CF sputum samples, diluted and centrifuged to remove insoluble material, we showed that both rSLPI and rSLPI-242 dose-dependently inhibited NE activity in the CF sputum supernatants. Although there was no difference between the two inhibitors with these sputum supernatants, addition of the oxidantN- chlorosuccinimide resulted in significantly more potent inhibition of sputum NE activity by rSLPI-242 compared with rSLPI. When fresly expectorated, neutrophil-rich CF sputum was immediately incubated with rSLPI and rSLPI-242 to evaluate the effects of oxidants released from neutrophils, inhibition of sputum NE activity by rSLPI-242 proved to be significantly higher than inhibition by rSLPI. Hence, due to its relative oxidation resistance rSLPI-242 seems to be superior to rSLPI in its ability to reduce NE activity in CF sputum.
Our results suggest that the superoxide anion radical is of major importance in the oxidation of rSLPI by fresh CF sputum, because the addition of SOD to the experimental incubation led to a significant improvement of sputum NE inhibition by rSLPI (Fig. 2). Nevertheless, our data also show that the superoxide anion radical does not account for all oxidative inactivation of rSLPI. There are multiple oxidants which may be present in CF sputum and be responsible for what goes beyond the oxidative effect of the superoxide anion radical. H2O2, however, seems to be an unlikely candidate, because there is no catalase deficiency in CF airways.
Our data also show how experimental conditions fundamentally influence the results. Although most in vitro studies on CF sputum are carried out with neutrophil-free sputum supernatants, which can easily be frozen and stored, our experiments with fresh, neutrophil-rich sputum provide quite different results that add important scientific information and are even more clinically relevant than those obtained with neutrophil-free sputum.
Not only did we study inhibition of CF sputum-induced cleavage of a NE-specific peptide substrate by rSLPI and rSLPI-242, but also inhibition of CF sputum-induced secretion from submucosal airway glands. Hypersecretion is a characteristic feature of bronchopulmonary disease in CF, and accumulation of secretions in the airways is an important cause of death in the disease(27). In humans, airway secretions are believed to originate predominantly from airway submucosal gland secretory cells whose volume is 40 times that of goblet cells in normal airways(28). In CF, with gland hypertrophy and hyperplasia(28, 29), submucosal gland secretory cells are even more conspicuous. It has already been demonstrated that CF sputum is a potent secretagogue for airway submucosal glands and that this secretagogue activity is mainly due to NE activity in the sputum(12). In the present study, a model of tracheal explants with abundant submucosal glands was used to investigate a paradigm of NE effects in CF pathophysiology, namely induction of hypersecretion and its inhibition by rSLPI and rSLPI-242. Our results show that rSLPI-242 inhibited NE-and CF sputum-induced secretion more potently than did rSLPI. For logistic reasons of the experimental design, the secretion studies with CF sputum could be carried out only with sputum supernatants, not with fresh, neutrophil-rich sputum. Superiority of rSLPI-242 over rSLPI in these experiments may have been due to oxidants released from the tracheal tissue explants; it is known that oxidants are generated within and outside of cells as a result of normal biochemical processes(30, 31). We speculate, however, that the difference between the inhibitory activity of rSLPI-242 and rSLPI on sputum-induced submucosal gland secretion will be even more pronounced in vivo where there are abundant polymorphonuclear leukocytes present in CF airways.
In summary, both rSLPI and rSLPI-242 were shown to inhibit CF sputum-induced cleavage of a NE-specific peptide substrate as well as CF sputum-induced secretion from submucosal airway glands in a dose-dependent fashion. Due to its relative oxidation resistance, rSLPI-242 appeared to be advantageous compared with rSLPI. As clinical studies on rSLPI aerosol therapy for CF have yielded promising, but yet unsatisfactory, results(32), we suggest that rSLPI-242 might be a more promising therapeutic agent than the native molecule.
Acknowledgements. The authors gratefully acknowledge the support from our colleagues at the CF clinics in Cologne (Dr. Ernst Rietschel) and Aachen (Dr. Gregor Kusenbach). We thank the patients for their participation in the study, and the physiotherapists for their help. Recombinant native SLPI (rSLPI) and its variant (rSLPI-242) were kindly provided by Dr. R. Heinzel-Wieland, Prof. Dr. G. Steffens, and Dr. D. Saunders, Grünenthal GmbH, Aachen, FRG.
Abbreviations
- CF:
-
cystic fibrosis
- α1 PI:
-
α1-proteinase inhibitor
- NE:
-
neutrophil elastase
- SLPI:
-
secretory leukoprotease inhibitor
- rSLPI:
-
recombinant SLPI
- SOD:
-
superoxide dismutase
- LDH:
-
lactic acid dehydrogenase
- IC50:
-
concentration that inhibits 50%
References
Tournier J-M, Jacquot J, Puchelle E, Bieth JG 1985 Evidence that Pseudomonas elastase does not inactivate the bronchial inhibitor in the presence of leukocyte elastase. Am Rev Respir Dis 132: 524–528.
Goldstein W, Döring G 1986 Lysosomal enzymes from polymorphonuclear leukocytes and proteinase inhibitors in patients with cystic fibrosis. Am Rev Respir Dis 134: 49–56.
Suter S, Schaad UB, Tegner H, Ohlsson K, Desgrandchamps D, Waldvogel FA 1986 Levels of free granulocyte elastase in bronchial secretions from patients with cystic fibrosis: effect of antimicrobial treatment against Pseudomonas aeruginosa. J Infect Dis 153: 902–909.
Koch C, Høiby N 1993 Pathogenesis of cystic fibrosis. Lancet 341: 1065–1069.
Weissmann G, Zurier RB, Hoffstein S 1972 Leukocytic proteases and the immunological release of lysosomal enzymes. Am J Pathol 68: 539–563.
Ohlsson K, Olsson I 1977 The extracellular release of granulocyte collagenase and elastase during phagocytosis and inflammatory processes. Scand J Haematol 19: 145–152.
Bruce MC, Poncz L, Klinger JD, Stern RC, Tomashefski JR, Dearborn DG 1985 Biochemical and pathologic evidence for proteolytic destruction of lung connective tissue in cystic fibrosis. Am Rev Respir Dis 132: 529–535.
Berger M, Soerensen RU, Tosi MF, Dearborn DG, Döring G 1989 Complement receptor expression on neutrophils at an inflammatory site, the Pseudomonas-infected lung in cystic fibrosis. J Clin Invest 84: 1302–1313.
Fick RB, Naegel GP, Squier S, Wood RE, Gee JBL, Reynolds HY 1984 Proteins of the cystic fibrosis respiratory tract: fragmented immunoglobulin G opsonic antibody causing defective opsonophagocytosis. J Clin Invest 74: 236–248.
Sommerhoff CP, Nadel JA, Basbaum CB, Caughey GH 1990 Neutrophil elastase and cathepsin G stimulate secretion from cultured bovine airway gland serous cells. J Clin Invest 85: 682–689.
Schuster A, Ueki I, Nadel JA 1992 Neutrophil elastase stimulates tracheal submucosal gland secretion that is inhibited by ICI 200:355. Am J Physiol 262:L86–L91.
Schuster A, Fahy JV, Ueki I, Nadel JA 1995 Cystic fibrosis sputum induces a secretory response from airway gland serous cells that can be prevented by neutrophil protease inhibitors. Eur Respir J 8: 10–14.
Thompson RC, Ohlsson K 1986 Isolation properties and complete amino acid sequence of human secretory leukocyte protease inhibitor, a potent inhibitor of leukocyte elastase. Proc Natl Acad Sci USA 83: 6692–6696.
Boudier C, Pelletier A, Gast A, Tournier JM, Pauli G, Bieth JG 1987 The elastase inhibitory capacity and theα1-proteinase inhibitor and bronchial inhibitor content of bronchoalveolar lavage fluids from healthy subjects. Biol Chem Hoppe-Seyler 368: 981–990.
Kramps JA, Franken C, Dijkman JH 1988 Quantity of antileukoprotease relative to α1-proteinase inhibitor in peripheral air spaces of the human lung. Clin Sci 75: 351–353.
Morrison HM, Kramps JA, Dijkman JH, Stockley RA 1986 Comparison of concentrations of two proteinase inhibitors, porcine pancreatic elastase inhibitory capacity, and cell profiles in sequential bronchoalveolar lavage samples. Thorax 41: 435–441.
Heinzel-Wieland R, Steffens GJ, Flohé L 1990 Inhibitory characteristics and oxidation resistance of site-specific variants of recombinant human antileukoproteinase. Biomed Biochim Acta 50: 677–681.
Wood RE, Boat TF, Doershuk CF 1976 Cystic fibrosis. Am Rev Respir Dis 113: 833–878.
Goco RV, Kress MB, Brantigan OC 1963 Comparison of mucus glands in the tracheobronchial tree of man and animals. Ann NY Acad Sci 106: 555–571.
Zar JH 1984 Biostatistical Analysis, 2nd Ed. Prentice-Hall, Englewood Cliffs, NJ, pp 122–160.
Berger M 1991 Inflammation in the lung in cystic fibrosis. A vicious cycle that does more harm than good?. Clin Rev Allergy 9: 119–142.
O'Connor CM, Gaffney K, Keane J, Southey A, Byrne N, O'Mahoney S, Fitzgerald MX 1993 α1-Proteinase inhibitor, elastase activity, and lung disease severity in cystic fibrosis. Am Rev Respir Dis 148: 1665–1670.
Cantin A, Bilodeau G, Begin R 1989 Granulocyte elastase-mediated proteolysis of α1-antitrypsin in cystic fibrosis bronchopulmonary secretions. Pediatr Pulmonol 7: 12–17.
Ossanna P, Test S, Matheson N, Regiani S, Weiss S 1986 Oxidative regulation of neutrophil elastase-α1-proteinase inhibitor interactions. J Clin Invest 77: 1939–1951.
Kramps JA, Van Twisk Ch, Klasen EC, Dijkman JH 1988 Interactions among stimulated human polymorphonuclear leucocytes, released elastase and bronchial antileucoprotease. Clin Sci 75: 53–62.
Vogelmeier C, Hubbard RC, Fells GA, Schnebli H-P, Thompson RC, Fritz H, Crystal RG 1991 Anti-neutrophil elastase defense of the normal human respiratory epithelial surface provided by the secretory leukoprotease inhibitor. J Clin Invest 87: 442–448.
Tizzano EF, Buchwald M 1992 Cystic fibrosis: beyond the gene to therapy. J Pediatr 120: 337–349.
Reid L 1960 Measurement of the bronchial mucous gland layer: a diagnostic yardstick in chronic bronchitis. Thorax 15: 132–141.
Sobonya RE, Taussig LM 1986 Quantitative aspects of lung pathology in cystic fibrosis. Am Rev Respir Dis 134: 290–295.
Crystal RG 1991 Oxidants and respiratory tract epithelial injury: pathogenesis and strategies for therapeutic intervention. Am J Med 91 ( suppl 3C): 39S–44S.
Wallaert B, Aerts C, Gressier B, Gosset P, Voisin C 1993 Oxidative inactivation of α1-proteinase inhibitor by alveolar epithelial type II cells. J Appl Physiol 75: 2376–2382.
McElvaney NG, Doujaiji B, Moan MJ, Burnham MR, Wu MC, Crystal RG 1993 Pharmacokinetics of recombinant secretory leukoprotease inhibitor aerosolized to normals and individuals with cystic fibrosis. Am Rev Respir Dis 148: 1056–1060.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Schuster, A., Hansen, G., Zubrod-Eichert, C. et al. Effects of Native and Oxidation-Resistant Secretory Leukoprotease Inhibitor on Cystic Fibrosis Sputum: Inhibition of Neutrophil Elastase Activity and of Sputum-Induced Secretion from Porcine Tracheal Submuc. Pediatr Res 40, 732–737 (1996). https://doi.org/10.1203/00006450-199611000-00013
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-199611000-00013
