
Abstract
Varicella zoster infection in children can be complicated by acute idiopathic thrombocytopenic purpura (ITP). To determine the etiologic mechanism of this thrombocytopenia, we studied three children with clinically diagnosed varicella infection. Immunoblot analysis of these patients' anti-platelet antibodies identified a unique band at 85 kD. Characterization of this protein revealed that it was platelet surface glycoprotein V (GPV) because it was not affected by a disulfide bond reduction but was cleaved by thrombin. Bernard-Soulier syndrome (BSS) platelets deficient in GPIb-IX and GPV did not react with the sera from our varicella-infected study patients. There was no apparent cross-reactivity between anti-varicella antibody and patients' anti-GPV Ig. We report here the first cases of GPV as the target antigen in autoimmune thrombocytopenia.
Similar content being viewed by others

Main
Varicella-associated thrombocytopenia is well described in the pediatric literature(1–4). However, there is much controversy over the etiologic mechanism for thrombocytopenia in this clinical setting(5).
In the 1960s, most investigators argued that disseminated intravascular coagulation was the principal mechanism of platelet destruction(6). However, some researchers proposed a different mechanism, namely direct interaction between viruses and human blood platelets resulting in platelet aggregation, followed by phagocytosis or lysis(7,8). Espinoza and Kuhn(9) using electron microscopy identified viral particles in the megakaryocytes of a patient with varicella and proposed that direct viral invasion of platelet precursors caused thrombocytopenia. In 1978, Scott et al.(10) determined that viral-derived neuraminidase could cause enzymatic desialylation of platelets, leading to thrombocytopenia as the abnormal platelets were removed from the circulation by the reticuloendothelial system. In 1979, Feusner et al.(11) raised the possibility of an immune-mediated mechanism for platelet destruction. They observed that, in some patients with varicella and thrombocytopenia, megakaryocyte marrow pools were increased, platelet survival was markedly decreased, the rate of fibrinogen degradation was normal, and platelet-associated IgG was increased(11).
Credence to the theory of antibody-mediated platelet destruction was added by Winiarski(12) in 1989 when platelet-specific antibodies were identified in five children with active varicella infection and thrombocytopenia. This investigator used an immunoblotting technique to detect IgM antibody against three platelet surface GPs, namely GPIb, GPIIb, and GPIIIa.
In this study, we report three patients with varicella-associated thrombocytopenia who were found to have autoantibodies directed against platelet GPV. This is the first report of GPV as the target antigen in autoimmune thrombocytopenia.
METHODS
Patients. Three children (all male; ages 5,7, and 17 y) were diagnosed with varicella infection on the basis of clinical history and characteristic vesicular rash. All three patients had hemorrhagic skin lesions, and two had petechiae, ecchymoses, and hematuria 1-4 d after the appearance of the rash. The main laboratory indices of the patients are displayed in Table 1. Platelet counts at presentation ranged from 2,000 to 57,000/ μL. All three patients had elevated platelet-associated IgG levels. Two patients underwent bone marrow aspiration, which revealed large numbers of megakaryocytes. Treatment of these patients was as follows. 1) One patients with hematuria was initially transfused with 10 U of platelets without improvement in his platelet count. Subsequently, he received IVIgG 400 mg/kg/d for 5 d. 2) The second patient with hematuria was treated with 400 mg/kg/d IVIgG and oral prednisone.3) The third and least symptomatic patient received no therapy.
Materials. Unless otherwise specified all regent grade chemicals were obtained from Sigma Chemical Co., St. Louis, MO. Biotin-conjugated goat anti-human IgG and IgM, avidin, and biotin-conjugated horseradish peroxidase were obtained from Vector Laboratories, Burlingame, CA. Nitrocellulose paper was purchased from Schleicher & Schuell, Keene, NH. Blood from a patient with BSS was kindly supplied by Dr. Leonard Small, Dover, NH. Immobilized varicella proteins on nitrocellulose were a gift of Dr. Charles Grose, University of Iowa School of Medicine, Iowa City, IA(13).
Preparation of platelets for electrophoresis and immunoblotting. Blood from patients and normal blood group O controls was collected into acid citrate-dextrose anticoagulant with 5 mM EDTA (1:9, v:v) and kept at room temperature. Platelet-rich plasma was collected by differential centrifugation at 900 rpm for 15 min (Sorvall RC-3 centrifuge). Platelets were isolated by centrifugation at 2000 rpm for 15 min and then washed five times in TES-Tyrode's buffer containing 136 mM NaCl, 10 mM Tris-2-aminoethanesulfonic acid, 2.6 mM KCl, 0.5 mM NaH2PO4, 2 mM EDTA, 2 mM MgCl2, and 5 mM glucose at pH 7.4.
A similar method was used to isolate platelets from a patient with BSS. Differential centrifugation of whole blood at 600 rpm for 5 min permitted isolation of platelet-rich plasma. This centrifugation was repeated five times. After each centrifugation, the blood cells were resuspended in TES-Tyrode's buffer (see above) to their original test volume. The platelet-containing supernatants were pooled. Then BSS platelets were isolated by centrifugation at 2000 rpm for 15 min and washed five times in TES-Tyrode's buffer.
Immunoblotting assay of anti-platelet antibodies. Washed platelets from patients with primary varicella infection and normal controls were solubilized in SDS buffer and separated (108 platelets/lane) according to molecular weight by the method of Laemmli using 3% polyacrylamide stacking and 7.5% separating slab gels(14). Electrophoresis was performed at 15 V until the tracking dye (0.002% v:v bromphenol blue) reached the bottom of the gel. The platelet proteins were electrophoretically transferred to 0.45-μm pore size nitrocellulose paper and then exposed to patient sera for 30 min at 37 °C(15). The immunoblotting technique for identification of platelet proteins has been previously reported(16). Briefly, immobilized platelet proteins on nitrocellulose were blocked with irrelevant proteins and then incubated with patient sera in 1% BSA in PBS(1:50 dilution) for 1 h and washed five times in 1% BSA/PBS. Platelet proteins were next exposed to biotin-conjugated goat anti-human IgG or IgM (1:200 dilution), avidin, and biotin-conjugated horseradish peroxidase. Chloronaphthol and hydrogen peroxide were used to visualize the bands of Ig-bound antigen.
Characterization of the 85-kD antigenic protein. To assess the effect of reducing disulfide bonds in the platelet GP targets, platelets from patients and controls were exposed to 5% β-mercaptoethanol during solubilization. Then, SDS-PAGE and immunoblotting were performed as described above(14–16).
The effect of thrombin digestion on detection of Ig-bound platelet antigens was also assessed. Fresh, washed platelets (1 × 109 platelets/mL) in TES-Tyrode's buffer (described previously) were incubated with 2 U/mL human thrombin at 37 °C for 60 min. After centrifugation, the supernatant was discarded and the platelet pellets were immediately taken up in SDS-PAGE sample buffer for electrophoresis and immunoblotting(14–16).
BSS platelets deficient in GPIb-IX and GPV were used to further define the 85-kD antigenic site. BSS platelets isolated by differential centrifugation as described previously were exposed to SDS-PAGE (107 platelets/lane). The isolated platelet proteins were then incubated with patient sera antiglobulin, and detection reagents using the methods of the immunoblotting assay discussed above(16).
Assessment of cross-reactivity between anti-platelet antibodies and anti-varicella antibodies. Cross-reactivity experiments were done to determine whether the observed platelet autoantibody was identical to a subset of the same patient's anti-varicella antibodies. Each patient's serum Ig was exposed to SDS-PAGE separated varicella viral proteins that had been transferred to and immobilized on nitrocellulose paper by standard Western blotting techniques by Dr. Charles Grose, University of Iowa School of Medicine, Iowa City, IA(13). Anti-varicella antibodies were then eluted by overnight incubation of the nitrocellulose strip in 6 M guanidine-HCl in 0.5 mM Tris-HCl, pH 7.5. The eluate was renatured by dialysis against 1 L of the same buffer without guanidine-HCl over a 48-h period. This eluate was used to probe separated platelet proteins obtained by the immunoblotting methods described previously(16).
Using analogous methods, anti-platelet antibody was eluted from the original immunoblotting assay and tested for its ability to bind varicella protein antigens immobilized on nitrocellulose blots, provided by Dr. Grose(13). These anti-platelet antibodies were subsequently reexposed to immobilized platelet proteins to determine whether the elution process had destroyed antibody activity.
RESULTS
Patients. The two patients who were treated with IVIgG for their thrombocytopenia achieved normal platelet counts within 5 d of treatment. The one patient who did not require any pharmacologic therapy spontaneously recovered a normal platelet count 2 wk after diagnosis. All three patients were followed for 6-18 mo without recurrence.
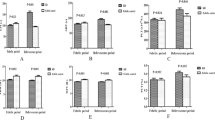
Immunoblotting assay. Incubating normal control platelet proteins with sera from our three varicella patients followed by exposure to enzyme conjugated anti-IgM revealed IgM binding to a unique band at 85 kDFig.1,lanes B, C, and D). This band was not observed when normal platelets were incubated with control sera (Fig. 1, lane A). The antigenic 85-kD protein was not of viral origin because the antibody reacted with platelets from an uninfected, normal donor in this immunoblotting experiment.

Immunoblotting results of sera from varicella-infected patients. A unique 85-kD antigenic site was identified by SDS-PAGE when normal control platelets were incubated with sera from three patients with varicella infection (lanes B, C, and D). This band was not observed when control serum was used (lane A).
In separate experiments, the binding of IgG antibodies was studied by a similar technique, substituting an anti-IgG antibody for an anti-IgM one. Only patient 2 had IgG antibody against the 85-kD antigen (seeTable 1).
To determine whether the antibodies against the 85-kD antigenic protein were autoantibodies, the patients' own platelets were also tested as antigenic targets. In all three patients, the antibody reacted equally well with autologous and homologous platelets, suggesting that these antibodies were directed against a common autoepitope (data not shown).
Characterization of the 85-kD antigenic protein. Platelet GPV is known to migrate in SDS-PAGE with an apparent molecular mass of 82-89 kD, depending upon the precise electrophoretic conditions(17, 18). Its mobility is not affected by reduction of disulfide bonds. The relative mobility of the 85-kD antigenic protein seen in our patients did not change in location or intensity after reduction of disulfide bonds (Fig. 2, lanes C and D).

Results of immunoblotting studies with platelets subjected to disulfide reduction. The 85-kD band was observed when patient serum was incubated with both nonreduced (NR) and disulfide-reduced(R) platelet proteins (lanes C and D).
GPV is the major platelet protein cleaved by thrombin(19). Its hydrolysis releases a soluble fragment known as GPVf1 (69.5 kD). In our experiments, thrombin digestion of control platelets before electrophoresis resulted in the elimination of the 85-kD band(Fig. 3, lane B). Antibody binding to the 69.5-kD proteolytic fragment was not observed in our immunoblotting assay. In the experimental methods used, soluble GPVf1 would have been discarded with the supernatant after thrombin digestion.

Results of immunoblotting experiment with varicella-infected patient's serum after thrombin digestion of the target platelets. There was no anti-IgM binding to 85- or 69.5-kD proteins (lane B).
BSS platelets have previously been shown by surface labeling techniques to be completely deficient in GPIb, GPIX, and GPV. No 85-kD band was detected upon incubation of BSS platelets with sera from varicella-infected patients(data not shown).
Assessment of cross-reactivity. Anti-varicella antibodies were detected in the sera of all three patients. To determine whether there was cross-reactivity between anti-varicella antibodies and anti-platelet antibodies, elution studies were undertaken. No binding of anti-varicella antibodies to platelet proteins was observed, nor were the eluates from immobilized platelet proteins cross-reactive against varicella viral proteins bound to nitrocellulose blots. However, the latter antibodies eluted from platelet proteins did rebind the 85-kD platelet antigen, indicating that the elution process had not destroyed antibody activity.
DISCUSSION
Acute ITP of childhood is an acquired hemorrhagic disorder which is generally self-limited. It is thought to be virally induced in most cases, but the inciting virus is rarely identified. The exception to this rule occurs in patients with varicella-zoster infection. As our study shows, the mechanism of thrombocytopenia in at least some patients with varicella infections may be immune-mediated platelet destruction via the production of autoantibodies against platelet surface GPV.
Our three patients with varicella and thrombocytopenia had a unique IgM antibody directed against an 85-kD platelet protein. This band was observed when uninfected control platelets were used as targets, suggesting that viral alteration of the platelet surface was not responsible for immune destruction.
Further characterization of the 85-kD platelet protein revealed that it was unaffected by disulfide bond reduction, but its antigenicity was destroyed by thrombin. These findings are typical features of platelet surface GPV and suggested that this GP contained our observed antigenic epitope. GPV has an apparent molecular mass of 82-89 kD, is not affected by disulfide bond reduction, and is cleaved by thrombin(17–20). To confirm this identification, BSS platelets were used as targets for immunoblotting. The 85-kD band was absent when BSS platelets were incubated with the patients' sera.
Our investigations also indicated that the unique IgM antibody found in our three patients with varicella-associated thrombocy-topenia was an autoantibody because it bound to the patients' own platelets. This autoantibody did not bind to varicella viral proteins. Elution assays failed to show any cross-reactivity between anti-platelet antibody and the patients' anti-varicella antibodies. However, viral cross-reactive anti-platelet Ig has been reported previously in a patient with varicella-associated ITP(21). Our negative results may reflect either a true absence of cross-reactivity with the GPV antigen or perhaps a sensitivity difference between the two studies in their respective abilities to detect very low titer antibody reactivity.
Alloantibodies against GPV have been reported to occur in several clinical situations. Patients who are polytransfused for hematologic diseases often develop leukocyte and platelet antibodies. In a large study of 106 patients receiving platelet transfusions, 45 developed platelet-specific antibodies(22). Immunoblotting analysis of 27 of these patients revealed that 78% reacted against GPV. Similarly, Ertem et al.(23) have identified a new platelet alloantigen, P1T, on GPV that is associated with neonatal alloimmune thrombocytopenia.
In this study, we report the first cases of varicella-associated autoimmune thrombocytopenia resulting from anti-GPV antibodies. We did not find Ig directed against GPIb, GPIIb, or GPIIIa as reported by Winiarski(12). The most likely explanation for these disparate results is that varicella-induced acute ITP is a heterogeneous disorder in which different platelet antigens may serve as epitopes for autoantibody production in different patients. A similar model exists in patients with chronic ITP. Autoantibodies against GPIIb-IIIa, GPIb-IX, GPIa-IIa, and GPIV have been reported in chronic ITP patients(24).
All three of our patients with varicella-associated thrombocytopenia did well clinically, and the two patients who were treated with IVIgG responded promptly. None of our patients went on to develop chronic ITP, although a very low level of anti-GPV Ig (IgG only) was detectable in one patient 1 y after resolution of his thrombocytopenia.
We conclude that autoantibodies to platelet surface GPV can be associated with clinically significant ITP in some patients with varicella-zoster infection. Investigation of the potential role of GPV in other autoimmune thrombocytopenic states is warranted.
Abbreviations
- ITP:
-
idiopathic thrombocytopenic purpura
- GP:
-
glycoprotein
- BSS:
-
Bernard-Soulier syndrome
- IVIgG:
-
i.v. gamma globulin
References
Fernbach DJ 1973 Bleeding and Infection. Am J Dis Child 125: 900
Ho-Yen Do, Hardie R, Sommerville RG 1984 Varicella-induced thrombocytopenia. J Infect 8: 274–276
Smith H 1967 Pupura fulminans complicating varicella: recovery with low molecular weight dextran and steroids. Med J Aust 2: 685–687
Ware R, Kurtzberg J, Friedman HS, Falletta JM, Kinney TR 1988 Chronic immunemediated thrombocytopenia after varicella infection. J Pediatr 112: 742–744
Yeager AM, Zinkham WH 1980 Varicella-associated thrombocytopenia: clues to the etiology of childhood idiopathic thrombocytopenic purpura. Johns Hopkins Med J 146: 270–274
McKay DG, Margaretten W 1967 Disseminated intravascular coagulation in virus diseases. Arch Intern Med 120: 129–152
Jerushalmy Z, Kohn A, deVries A 1961 Interaction of myxoviruses with human blood platelets in vitro. Proc Soc Exp Biol Med 106: 462–466
Terada H, Baldini M, Ebbe S, Madoff MA 1966 Interaction of influenza virus with blood platelets. Blood 28: 213–228
Espinoza C, Kuhn C 1974 Viral infection of megakaryocytes in varicella with purpura. Am J Clin Pathol 61: 203–208
Scott S, Reimers HJ, Chernesky MA, Greenberg JP, Kinlough-Rathbone RL, Packham MA, Mustard JF 1978 Effect of viruses on platelet aggregation and platelet survival in rabbits. Blood 52: 47–55
Feusner JH, Slichter SJ, Harker LA 1979 Mechanisms of thrombocytopenia in varicella. Am J Hematol 7: 255–264
Winiarski J 1989 Platelet antigens in varicella associated thrombocytopenia. Arch Dis Child 65: 137–139
Harper DR, Grose C 1989 IgM and IgG Responses to varicella-zoster virus p32/p36 complex after chickenpox and zoster, congenital and subclinical infections, and vaccination. J Infect Dis 159: 444–451
Laemmli UK 1970 Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680–685
Towbin H, Staehlin T, Gordon J 1979 Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA 76: 4350–4354
Beardsley DS, Spiegel JE, Jacobs MM, Handin RI, Lux SE 1984 Platelet membrane glycoprotein IIIa contains target antigens that bind anti-platelet antibodies in immune thrombocytopenia. J Clin Invest 74: 1701–1707
Berndt MC, Phillips DR 1981 Purification and preliminary physicochemical Characterization of human platelet membrane glycoprotein V. J Biol Chem 256: 59–65
Phillips DR, Agin PP 1977 Platelet plasma membrane glycoproteins: identification of a proteolytic substrate for thrombin. Biochem Biophys Res Commun 75: 940–947
Modderman PW, Admiraal LG, Sonnenberg A, Von dem Bonne AEGK 1992 Glycoproteins V and Ib-IX form a noncovalent complex in the platelet membrane. J Biol Chem 267: 364–369
Roth GJ, Church TA, McMullen BA, Williams SA 1990 Human platelet glycoprotein V: a surface leucine-rich glycoprotein related to adhesion. Biochem Biophys Res Commun 170: 153–161
Wright JF, Blanchette VS, Freedman J 1994 Virus-reactive antibodies cross-react with autologous platelets in a patient with varicella zoster virus (VZV)-associated idiopathic thrombocytopenic purpura (ITP). Blood 84( suppl): 185a.
Meenaghan M, Judson PA, Yousaf K, Lewis L, Pamphilon DH 1993 Antibodies to platelet glycoprotein V in polytransfused patients with haematologic diseases. Vox Sang 64: 167–170
Ertem M, Moulton T, Ho JS, Beardsley DS 1994 A new platelet alloantigen, P1T, on glycoprotein V associated with neonatal alloimmune thrombocytopenia. Pediatr Res 35: 160A
He R, Reid DM, Jones CE, Shulman WR 1994 Spectrum of Ig classes, specificities, and titers of serum antiglycoproteins in chronic idiopathic thrombocytopenic purpura. Blood 83: 1024–1032
Acknowledgements
The authors gratefully acknowledge the laboratory assistance of Joyce S. Ho, the generous gift of BSS platelets from Dr. Leonard Small, Dover, NH, the varicella viral proteins kindly supplied by Dr. Charles Grose, University of Iowa School of Medicine, Iowa City, IA, and the secretarial assistance of Lynn Puglisi, Pat Spica, and Heidi Zeitler.
Author information
Authors and Affiliations
Additional information
Supported in part by National Institutes of Health Grants HL01485 and HL38579 (to D.S.B.).
Rights and permissions
About this article
Cite this article
Mayer, J., Beardsley, D. Varicella-Associated Thrombocytopenia: Autoantibodies against Platelet Surface Glycoprotein V. Pediatr Res 40, 615–619 (1996). https://doi.org/10.1203/00006450-199610000-00017
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-199610000-00017
This article is cited by
-
Relapse of idiopathic thrombocytopenic purpura caused by influenza A virus infection: a case report
Journal of Infection and Chemotherapy (2004)
