
Abstract
We have previously shown that the immature tubule is tolerant of prolonged anoxia. In addition, cellular ATP is maintained at 2-fold higher levels during anoxia in the immature tubules compared with the mature tubules. The purpose of this study was: 1) to determine whether anaerobic glycolysis contributes to the tolerance to anoxia and preservation of cellular ATP in immature tubules and 2) to evaluate whether the tolerance demonstrated by immature tubules is dependent on preservation of cellular ATP. Suspensions of proximal tubules from immature (8-10 d) and mature (8-10 wk) rats were subjected to 15 and 45 min of anoxia in a standard buffer and in buffers designed to inhibit glycolysis. Lactate dehydrogenase release was used to assess plasma membrane damage, ATP levels were determined as an index of cellular energy and total lactate production was measured to evaluate glycolytic activity. After 45 min of anoxia, total lactate production was less in immature tubules (101 ± 48 μg of lactate/mg of DNA) compared with mature tubules (148 ± 36 μg of lactate/mg of DNA). After inhibition of glycolytic metabolism, ATP decreased to similar levels in both immature and mature tubules. However, immature tubules remained resistant to anoxic damage(lactate dehydrogenase: mature tubules 38 ± 4%, immature tubules 29± 1.0%). Therefore, enhanced glycolytic activity does not play a dominant role in the tolerance of the developing kidney to anoxia, and this tolerance is not primarily dependent on preservation of cellular ATP.
Similar content being viewed by others

Main
Most organisms have retained the capacity to produce energy from glucose by anaerobic glycolysis in conditions lacking oxygen. Glucose is converted to lactate, and energy is generated and stored in the form of ATP. In general, it has been suggested that immature organisms have greater glycolytic capacity than the mature organism under anaerobic conditions(1). We have previously shown that the immature nephron is tolerant of a prolonged anoxic insult compared with the mature nephron. In addition, cellular ATP is maintained at 2-fold higher levels in IT compared with MT(2). The objective of this study is: 1) to determine whether anaerobic glycolysis contributes to the tolerance to anoxia and 2) to evaluate whether this tolerance is dependent on preservation of cellular ATP.
METHODS
Preparation of suspensions enriched in tubule segments. To obtain tubule segments from immature and mature rat kidneys, we used a modification of our previous procedures(3). Mature rats(8-10 wk of age) were anesthetized with Inactin (Byk Guiden, Konstanz Germany)(sodium thiobutabarbital, 80 mg/kg body weight, intraperitoneally), and the aorta was carefully cannulated with PE-90 tubing distal to the origin of both renal arteries. To initiate the harvest procedure, the left renal vein was incised and the kidneys were flushed with 15 mL of buffered Ringer's solution containing metabolic substrates that had been equilibrated with 95% O2/5% CO2 at 37 °C. The kidneys were perfused over 10 min with an enzyme solution which contained 90 mg of collagenase (Boehringer Mannheim, Indianapolis, IN) and 2500 caseinolytic units of dispase(Collaborative Biomedical Products, Bedford, MA) were dissolved in 50 mL of buffered Ringer's solution heated to 35 °C and then flushed with 10 mg of cold buffered Ringer's solution. Both kidneys were rapidly excised and placed on ice, the renal capsules were removed, and the cortex was gently scraped. The tubules were suspended in 25 mL of buffered Ringer's solution, placed on ice, and stirred for 15 min. The suspension was filtered through a 115-μm fabric mesh (Tetko, Briarcliff Manor, NY) and then washed and spun at 500 rpm three times for 2 min. The tubule suspension was equilibrated with 95% O2/5% CO2 and kept at 4 °C until studied. The final pellet was resuspended with the buffered Ringer's solution to a concentration of approximately 2 mg of protein/mL.
To obtain tubules from the immature rats (8-10 d of age), the pups were anesthetized with ether, and the kidneys were excised and placed in a buffered Ringer's solution. The capsules were removed, and the kidneys were immersed in 10 mL of buffered Ringer's solution which contained 20 mg of collagenase(Boehringer Mannheim) and 500 caseinolytic units of dispase (Collaborative Biomedical Products) and placed in a shaking water bath at 35 °C. After 15 min, the kidneys were placed on ice, and the cortex was gently scraped. Tubules were suspended in 25 mL of cold buffered Ringer's solution and stirred for 15 min. To eliminate glomeruli, the suspension was filtered through a 50-μm fabric mesh (Tetko). The suspension was equilibrated with 95% O2/5% CO2 then washed, spun at 500 rpm three times for 2 min, and kept at 4 °C until studied.
LDH. To assess the integrity of the suspension of tubules, LDH release was determined as an index of plasma membrane damage. LDH was assayed by the procedures described by Takano et al.(4). Released LDH (supernatant LDH) is expressed as a percent of total LDH (supernatant LDH plus cellular LDH), which is determined after sonication of tubules(2).
Determination of cellular ATP levels. ATP levels were measured as an index of the energy metabolism of the tubules. Intracellular ATP concentration was measured using a modification of the luciferase assay described by Uchida and Endou(5). Adenine nucleotides were extracted in a 1:1 ratio of 0.5 mg of perchloric acid. ATP was measured in 100-μL samples using a luminometer (Lumat LB 9501; Berthold, Wallac Inc., Gaithersburg, MD) and an ATP bioluminescent assay kit (Sigma Chemical Co., St. Louis, MO).
DNA content. To allow a direct comparison between IT and MT, the data are expressed per mg of DNA. Because IT increase in size by hypertrophy rather than hyperplasia, DNA more accurately reflects the number of cells being studied than the protein content of the tubule suspension. The DNA content of the tubule suspension was measured by the benzimide fluorimetric method (Sigma Chemical Co.)(6).
Determination of lactate levels. Lactate was measured on a model 27 industrial analyzer using a YSI 2746-lactate kit (Yellow Springs Instrument Co., Yellow Spring, OH) under basal oxygenated conditions and after 15 and 45 min of anoxia. Because it is unknown whether there is intracellular sequestration of lactate in the developing tubules, total lactate production was measured after lysing tubules by sonication. Because lactate produced during the equilibration period (10 min) represents glycolysis during rewarming to steady state, this value was subtracted from subsequent measurements.
Preparation of a buffer solution for studies of glycolysis. In suspensions of proximal tubules, lactate production can be used as an index of anaerobic metabolism because glycogen is not stored and the synthesis and degradation of glycogen does not contribute significantly to carbohydrate metabolism in this tissue(7). A common buffer solution which contained alanine, lactate, and glutamine was previously developed in which both MT and IT were stable(2). Adjustments needed to be made to the common buffer solution to eliminate lactate and alanine because alanine can be converted to lactate through pyruvate(7). In addition, glutamine is gluconeogenic in the proximal tubule and also needed to be removed(7). Thus, a new standard buffer solution was developed which contained the amino acids butyrate (10 mM) and malate (3 mM) as the principle substrates, as well as glucose (5.5 mM), NaCl (112 mM), KCl (3 mM), MgSO4 (1.2 mM), KH2PO4 (1 mM), CaCl2 (1.5 mM), and NaHCO3 (25 mM,) with a final osmolarity of 305-310 mosmol/L and a pH of 7.4. Two separate inhibitory buffers were designed. Inhibitory buffer A contained L-glucose, a nonmetabolizable sugar in substitution for D-glucose, the principal substrate required for glycolysis. Inhibitory buffer B contained 1 mM Iodoacetate which inhibits the degradation of glyceraldehyde 3-phosphate, thereby blocking the glycolytic pathway(8).
Induction of anoxia. Tubules from both the immature and mature rats were suspended in either new standard buffer or one of the inhibitory buffers and subjected to 15 and 45 min of anoxia. Before anoxia, the tubule suspension was preequilibrated at 37 °C and bubbled with 95% O2/5% CO2 for 10 min to allow for steady state equilibration. Anoxia was produced by gassing an aliquot for two minutes with 95% N2/5% CO2 by which time the Po2 fell to zero when measured by a polarographic oxygen electrode (Clark). The microtube was capped and incubated at 37 °C in a shaking water bath for the designated anoxic interval. All studies were performed at the end of the anoxic period.
Statistics. All values are reported as mean ± SD. Comparisons between groups are made by the t test with Bonferroni's modification for multiple comparisons as well as analysis of variance. Values were considered significantly different if p < 0.05.
RESULTS
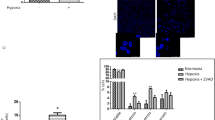
Effects of anoxia on cell ATP, injury, and lactate production. Under basal conditions, cellular ATP levels and LDH release in both IT and MT suspended in the new standard buffer were comparable to those previously reported(2). To study the effect of anoxia in the new standard buffer, ATP levels and LDH release were determined after 15 and 45 min of anoxia in MT and IT (Fig. 1, A and B). Because there is a difference in absolute values of ATP under basal conditions between the mature (245 ± 72; nmol of ATP/mg of DNA) and IT (96 ± 18 nmol of ATP/mg of DNA), alterations in ATP are expressed as percent basal values for comparison between groups. At both anoxic periods, MT had a significantly greater fall in cellular ATP than IT (p < 0.05) (Fig. 1A), and both groups of tubules reached levels similar to those previously reported(2).

(A) Intracellular ATP as percentage of control values in the MT (closed bars) and IT (open boxes) after 15 and 45 min of anoxia. (B) Percentage of LDH released from MT (closed bars) and IT (open bars) after 15 and 45 min of anoxia. (C) Total lactate production (μg/mg of DNA) in MT(closed bars) and IT (open bars) after 15 and 45 min of anoxia. Results are expressed as mean ± SD. *p < 0.05 compared with values between MT and IT at each time interval.
The degree of injury was determined after 15 and 45 min of anoxia (Fig. 1B). The release of LDH increased from 29 ± 4% at 15 min to 37 ± 6% at 45 min in the MT (p < 0.05), indicating more severe damage with prolonged anoxia. As previously reported, membrane integrity stabilized in the IT after 15 min of anoxia (LDH 26± 2%) with a slight, but not significant, increase (LDH 28 ± 2%) after 45 min of anoxia. The LDH values of 45 min of anoxia were significantly different between the MT and IT (p < 0.05). Therefore, the relative resistance of the IT to anoxia was confirmed(2).
Total lactate production was determined after 15 and 45 min of anoxia (Fig. 1C). After 15 min of anoxia, total lactate production in MT (56 ± 15 μg of lactate/mg of DNA) was greater than in IT (37 ± 18 μg of lactate/mg of DNA) (p < 0.05). After prolonged anoxia, total lactate production increased 3-fold in both MT(148 ± 36 μg of lactate/mg of DNA) and IT (101 ± 48 μg of lactate/mg of DNA), but was still significantly lower in IT.
The effect of anoxia after inhibition of the glycolytic pathway(Table 1)
Total lactate production was successfully inhibited to a similar degree in the MT and IT in buffer A with substitution of L-glucose (MT 73 ± 13%, IT 75 ± 16% inhibition) and in buffer B with the addition of iodoacetate (MT 93 ± 7%, IT 95 ± 10% inhibition). In both IT and MT greater inhibition of glycolytic metabolism was achieved in buffer B.
Inhibition of glycolytic metabolism during 45 min of anoxia diminished intracellular ATP slightly but not significantly in the MT (from 19 ± 2% in standard buffer to 19 ± 2% in buffer A, and 12 ± 3% in buffer B). Inhibition of glycolysis in IT resulted in a significant decrease in intracellular ATP in both inhibitory buffers (from 30 ± 5% in standard buffer to 17 ± 8% in buffer A, 11 ± 3% in buffer B)(p < 0.05). Of note, the ATP levels in both MT and IT were decreased to a similar degree in the respective inhibitory buffers.
In the MT, the LDH release after 45 min of anoxia was not affected by inhibition of glycolytic metabolism (38 ± 4%, buffer A, 41 ± 6% buffer B). If the glycolytic pathway were a dominant contributor to the tolerance of the IT to anoxia, one would anticipate that blocking this pathway would result in greater injury of the IT reflected by an increase in LDH release after anoxia. However, inhibition of glycolytic activity did not alter the LDH release in the IT (29 ± 1%, buffer A, 32 ± 2% buffer B). LDH release remained significantly different comparing values in each inhibitory buffer between MT and IT (p < 0.05). Therefore, data in Table 1 demonstrate that the tolerance of the IT to anoxia is not affected by inhibition of glycolytic metabolism.
DISCUSSION
This study addresses two important issues regarding the resistance of the IT to anoxia. First, despite traditional teachings(9), the data demonstrates that anaerobic glycolysis is not primarily responsible for the tolerance of the developing kidney to anoxia. Early investigators postulated the tolerance of the newborn to be due, in part, to nonoxidative glycolytic metabolism(10, 11). Whittam(10) demonstrated rabbit renal cortical slices had a 5-fold greater rate of glycolysis during anoxia compared with adult tissue by comparing active transport of Na and K during aerobic and anaerobic incubation. Dicker and Shirley(11) confirmed these observations in the newborn rat. Wachstein and Bradshaw(12) showed an age dependent decline of oxidative enzyme activity in the renal tubules of the newborn rat and rabbit.
To investigate the role of anaerobic metabolism in the tolerance of the IT to anoxia, lactate was measured as an index of glycolytic activity. The common buffer solution with essential substrates to support both MT and IT needed to be altered because it contained lactate, alanine and glutamine which would directly interfere with the measurement of lactate. In addition, because alanine and glycine protect against hypoxic injury in renal proximal tubules the effect of the removal of these substances on the renal injury needed to be defined(13). To avoid any contribution of heat stress to the tolerance of the IT to anoxia, we also modified the harvest procedure so the temperature was 35 °C or less throughout (see Methods). Our present studies show that the integrity and metabolic stability of the proximal tubule suspensions were not affected by the new standard buffer or harvest technique under control conditions. In addition, as shown in Figure 1, A and B, we confirmed the resistance of the IT to prolonged anoxia in the new standard buffer.
The present studies, however, demonstrate that the glycolytic pathway is not primarily responsible for the tolerance of IT to prolonged anoxia. Investigations of the developing CNS demonstrate findings similar to this study. Duffy et al.(14) demonstrated that tolerance of the fetal rat to anoxia was inversely proportional to age. Although early explanations for resistance to anoxia was a greater dependence upon anaerobic glycolysis for energy production, later studies emphasized low energy requirements of the newborn brain as the principle factor(15, 16). Swaab and Boer(17) showed energy consumption in 7-d-old rats was 1/10 of adult, and others showed lower cerebral glycolytic capacity in developing mice(18). In addition, Duffy et al.(14) calculated the percentage of cerebral energy requirements that could be generated at maximal glycolytic flux for fetal and 1d-old rats and noted the 2-fold longer survival of fetal rats in nitrogen could not be attributed to an increased capacity for anaerobic glycolysis. Comparable findings have been noted in immature renal tubules. Jacobs et al.(19) confirmed the stability of LDH levels in young rabbits (age 2-4 wk) after 30 min of anoxia compared with the adult and demonstrated that lactate production was not increased from baseline in young rabbits after anoxia. The present data are consistent with these findings and demonstrate, as well, that the inhibition of glycolysis does not diminish the tolerance of the IT to anoxia. Because Madin-Darby canine kidney cells are more resistant overall to toxic injuries, Sheridan et al.(20) investigated the resistance of these cells to chemical anoxia compared with mouse proximal tubule cells in primary culture. This resistance was shown to be independent of glycolysis and was demonstrated to be due, in part, to the marked differences in the amount of FFA accumulated(20).
The second issue this study addresses is that the tolerance of the IT to oxygen deprivation is not dependent on maintenance of intracellular ATP levels. A decline in ATP is the hallmark of an anoxic or ischemic renal injury. Preservation of ATP appears to be a fundamental response of the immature animal because it occurs in many tissues such as hepatocytes(21), central neurons(22), and kidney and has been thought to play a role in the tolerance of the immature animal to oxygen deprivation. It is well documented that the neonatal brain is more resistant to anoxia than that of the adult(22). The underlying mechanisms for this tolerance are unclear, but it is postulated that the newborn brain can decrease the metabolic rate to preserve cellular ATP(23). In rat brain slices, Kass and Lipton(23) have shown that ATP levels in the young animal are higher than those in the adult after anoxia. We and others have shown preservation of cellular ATP in the immature kidney which has been subjected to anoxia(2, 9). This study confirmed preservation of cellular ATP at 2-fold higher levels in the IT during anoxia. Inhibition of glycolysis during anoxia eliminated the contribution of anaerobic metabolism to ATP synthesis, and the cellular ATP in the IT was concomitantly reduced to values similar to that of the MT (Table 1). When lactate production is inhibited to greater than 90% and ATP levels are reduced to 9-12% of control values, there is no significant change in tubule integrity as measured by LDH release in either the MT or IT. Thus, maintenance of ATP levels cannot be the sole explanation of the tolerance of the IT to injury.
Several possible explanations exist for the tolerance of the IT to anoxia. Over the past several years, studies have shown a central role for disruption and reorganization of the cytoskeleton in ischemic renal injury(24). In the developing kidney the lack of complete polarization may be a protective mechanism. Alternatively a low intracellular pH (pHi) is a factor which is known to provide tolerance of MT to anoxia(25). IT may have a lower steady-state pHi than MT or the pHi response of the IT to oxygen deprivation may differ from that of MT. Also, an elevation of intracellular calcium concentration is associated with cellular dysfunction(26). There may exist a difference between calcium handling in the IT and MT. Lastly, heat stress proteins have been associated with cytoprotection in the mature kidney(27). Perhaps there are differences in constitutively expressed or stressed induced proteins in the immature kidney compared with the mature organ. If the stress response proteins are more abundantly expressed or more rapidly induced in the IT, one would anticipate an enhanced tolerance to an anoxic insult. Preliminary data would suggest that differences are present in levels of stress proteins during development(28).
The data in the present study demonstrate 1) the developing kidney is more tolerant of prolonged anoxia documented by preservation of cellular ATP and membrane integrity; 2) total lactate production during anoxia is less in IT compared with MT and, therefore, enhanced glycolytic activity does not play a central role in this tolerance;3) inhibition of glycolysis results in lower ATP levels in IT but does not worsen the injury; and 4) maintenance of intracellular ATP does not provide the primary explanation of the tolerance of the developing kidney.
Abbreviations
- LDH:
-
lactate dehydrogenase
- IT:
-
immature tubule
- MT:
-
mature tubule
- pHi:
-
intracellular pH
References
Barac-Nieto M, Spitzer A 1988 The relationship between renal metabolism and proximal tubule transplant during ontogeny. Pediatr Nephrol 2: 356–367
Gaudio KM, Thulin G, Ardito T, Kashgarian M, Siegel NJ 1994 Immature renal tubules are resistant to prolonged anoxia. Pediatr Nephrol 35: 152–156
Gaudio KM, Thulin G, Ardito T, Kashgarian M, Siegel NJ 1989 Metabolic alterations in proximal tubule suspensions obtained from ischemic kidneys. hysiol 257:F383–F389
Takano T, Soltoff S, Murdaugh S, Mandel L 1985 Intracellular respiratory dysfunction and cell injury in short term anoxia of rabbit renal proximal tubules. J Clin Invest 76: 2377–2384
Uchicha S, Endou H 1988 Substrate specificity to maintain cellular ATP along the mouse nephron. Am J Physiol 255:F977–F983
LaBanca C, Paigen K A simple rapid and sensitive DNA assay procedure. Anal Biochem 102: 344–352
Mandel LJ 1985 Metabolic substrates, cellular energy production and the regulation of proximal tubular transport. Annu Rev Physiol 47: 85–101
Misfeldt D, Hamamoto S, Pitelka D 1976 Transepithelial transport in cell culture. Proc Natl Acad Sci USA 73: 1212–1216
Fisher J, Isselhard W 1975 Metabolic patterns in severe tissues of newborn rabbits during ischemia. Biol Neonate 27: 235–250
Whittam R 1961 Metaboic changes in rabbit kidney cortex during the few weeks after birth. Biochim Biophys Acta 54: 574–576
Dicker S, Shirley D 1971 Rates of oxygen consumption and of anaerobic glycolysis in renal cortex and medulla of adult and newborn rats and guinea pigs. J Physiol (Paris) 212: 235–243
Wachstein M, Bradshaw M 1965 Histochemical localization of enzyme activity in the kidneys of three mammalian species during their postnatal development. J Histochemi Cytochem 13: 44–56
Garza-Quintero R, Ortega-Lopez J, Stein JH, Venkatachalam MA 1990 Alanine protects rabbit proximal tubules against anoxic injury in vitro. Am J Physiol 258:F1075–F1083
Duffy TE, Kohle SJ, Vanucci RC 1975 Carbohydrate and energy metabolism in perinatal rat brain: relation to survival in anoxia. J Neurochem 24: 271–276
Fazekas SJ, Alexander F, Himwich H 1941 Tolerance of the newborn to anoxia. Am J Physiol 151:F479–F483
Lawry OH, Passonneau JV, Hasselberger FX, Schulz DW 1964 Effect of ischemia on known substrates and cofactors of the glycolytic pathway in brain. J Biol Chem 239: 18
Swaab DF, Boer K 1972 The presence of biologically labile compounds during ischemia and their relationship to the EEG in the rat cerebral cortex and hypothalamus. J Neurochem 19: 2843–2853
Thurston JH, McDougal DB 1969 Effect of ischemia on metabolism of the brain of the newborn mouse. Am J Physiol 216:F348–F352
Jacobs WR, Slott P, Moore E 1992 Lactate production during anoxia in renal proximal tubules from young and adult rabbits. Pediatr Res 31: 335A( abstr)
Sheridan AM, Schwartz JH, Kroshain VM, Tercyak AM, Larcia J, Masino S, Lieverthal W 1993 Renal mouse proximal tubular cells are more susceptible than MDCK cells to chemical anoxia. Am J Physiol 265:F342–F350
Aw, TY, Jones DP 1987 Respiratory characteristics of neonatal rat hepatocytes. Pediatrics 21: 492–496
Haddad GG, Jiang C 1993 O2 deprivation in the central nervous system: on mechanisms of neuronal response differential sensitivity and injury. Prog Neurobiol 40: 277–318
Kass IS, Lipton P 1989 Protection of hypocampal slices from young rats against anoxic transmission damage is due to better maintenance of ATP. J Physiol (Lond) 413: 1–11
Spiegel DM, Molitoris BA 1989 Epithelial polarity following ischemia: A requirement for normal cell function. Am J Physiol 256:F430–F436
Bate MI, Ingwall JS 1994 Acidosis during ischemia promotes adenosine triphosphate resynthesis in postischemic rat heart:in vivo regulation of 5 ′ nucleotidase. J Clin Invest 93: 40–49
Kribben A, Wieder ED, Wetyels JF, Yu L, Gengaro PE, Burke TJ, Schner RW 1994 Evidence for role of cytosolic free calcium in hypoxia induced proximal tubule injury. J Clin Invest 93: 1922–1929
Borkum SC, Emami A, Schwartz JH 1993 Heat stress protein-associated cytoprotection of inner medullary collecting direct cells from rat kidney. Am J Physiol 265:F333–F341
Udelsman R, Blake M, Stagg O, Li D, Putney D, Holbrook N 1993 Vascular heat shock protein expression in response to stress. J Clin Invest 91: 465–473
Acknowledgements
The authors thank Kelli Chung for assistance in carrying out the experiments. They are also grateful to Melanie-Dawn Belanger and Marie Campbell for secretarial support in preparation of the manuscript.
Author information
Authors and Affiliations
Additional information
Supported by National Institutes of Health Grants DK44336 and HD32573.
Rights and permissions
About this article
Cite this article
Gaudio, K., Thulin, G. & Siegel, N. Glycolysis Is Not Responsible for the Tolerance of Immature Renal Tubules to Anoxia. Pediatr Res 40, 457–461 (1996). https://doi.org/10.1203/00006450-199609000-00015
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-199609000-00015
This article is cited by
-
Modulation of renal cell injury by heat shock proteins: lessons learned from the immature kidney
Nature Clinical Practice Nephrology (2006)
