
Abstract
Colorectal cancers (CRCs) often show a dense infiltrate of cytokine-producing immune/inflammatory cells. The exact contribution of each immune cell subset and cytokine in the activation of the intracellular pathways sustaining CRC cell growth is not understood. Herein, we isolate tumor-infiltrating leukocytes (TILs) and lamina propria mononuclear cells (LPMCs) from the tumor area and the macroscopically unaffected, adjacent, colonic mucosa of patients who underwent resection for sporadic CRC and show that the culture supernatants of TILs, but not of LPMCs, potently enhance the growth of human CRC cell lines through the activation of the oncogenic transcription factors signal transducer and activator of transcription 3 (STAT3) and nuclear factor-kappa B (NF-kB). Characterization of immune cell complexity of TILs and LPMCs reveals no differences in the percentages of T cells, natural killer T cells, natural killer (NK) cells, macrophages and B cells. However, T cells from TILs show a functional switch compared with those from LPMCs to produce large amounts of T helper type 17 (Th17)-related cytokines (that is, interleukin-17A (IL-17A), IL-17F, IL-21 and IL-22), tumor necrosis factor-α (TNF-α) and IL-6. Individual neutralization of IL-17A, IL-17F, IL-21, IL-22, TNF-α or IL-6 does not change TIL-derived supernatant-driven STAT3 and NF-kB activation, as well as their proproliferative effect in CRC cells. In contrast, simultaneous neutralization of both IL-17A and TNF-α, which abrogates NF-kB signaling, and IL-22 and IL-6, which abrogates STAT3 signaling, reduces the mitogenic effect of supernatants in CRC cells. IL-17A, IL-21, IL-22, TNF-α and IL-6 are also produced in excess in the early colonic lesions in a mouse model of sporadic CRC, associated with enhanced STAT3/NF-kB activation. Mice therapeutically given BP-1-102, an orally bioavailable compound targeting STAT3/NF-kB activation and cross-talk, exhibit reduced colon tumorigenesis and diminished expression of STAT3/NF-kB-activating cytokines in the neoplastic areas. These data suggest that strategies aimed at the cotargeting of STAT3/NF-kB activation and interaction between them might represent an attractive and novel approach to combat CRC.
Similar content being viewed by others

Introduction
Colorectal cancer (CRC) is the second leading cause of cancer-related death in the western world.1 The development of CRC is a multistage process, characterized by complex interactions between environmental carcinogens, genetic alterations and the host immune system, ultimately resulting in the uncontrolled growth of transformed cells.2 Similar to other common malignancies (for example, hepatocellular carcinoma, prostate carcinoma, gastric cancer), chronic inflammation is an independent risk factor for the development of CRC. For example, in patients with ulcerative colitis, there is a marked increase in the incidence of CRC.3 Experimental models of inflammation-associated colon carcinogenesis suggest that inflammatory cell-derived cytokines either directly or indirectly stimulate the growth of cancer cells.4, 5, 6, 7, 8, 9, 10 Nevertheless, under specific inflammatory conditions, immune cells can also mediate antitumor responses with the downstream effect of eliminating dysplastic and cancerous cells.11,12 Notably, sporadic CRC, which represent the majority of CRC cases, exhibit extensive inflammatory infiltrates with high levels of cytokine expression in the tumor microenvironment. In this context, the production of interferon γ (IFN-γ) by T helper type 1 (Th1) CD4+ cells, CD8+ cells and natural killer (NK) cells has been demonstrated to limit tumor growth by activating cytotoxic immunity,13, 14, 15, 16 and the presence of Th1 polarization markers correlates with reduced tumor recurrence in CRC patients.17 In contrast, tumor specific upregulation of cytokines produced by Th17 CD4+ cells, such as interleukin-17A (IL-17A) and IL-22, is detected in human CRC18, 19, 20, 21 and studies in mouse models of spontaneous intestinal tumorigenesis have proven the importance of these cytokines in facilitating tumor promotion and progression.5,21,22 Consistently, a Th17 immune cell infiltrate negatively influences the prognosis of CRC patients.23, 24, 25 Although progress has been made, the molecular mechanisms by which inflammation promotes CRC development are still being uncovered. This study was aimed at characterizing immune/inflammatory infiltrate and cytokine response in sporadic CRC and determining the signaling pathways by which cytokines produced by tumor-infiltrating leukocytes (TILs) modulate CRC cell growth.
Results
Culture supernatants of TILs increase CRC cell proliferation through the activation of STAT3 and NF-kB
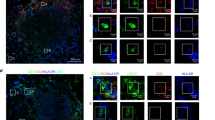
We isolated TILs and lamina propria mononuclear cells (LPMCs) from the tumor area and the macroscopically unaffected, adjacent, colonic mucosa of patients who had undergone resection for sporadic CRC and assessed whether TIL- and LPMC-derived supernatants modulate CRC cell proliferation. TIL-derived supernatants induced a robust proliferation of both DLD-1 and HT-29 cells after 24 h as compared with LPMC-derived supernatants (Figure 1a). No changes in the rate of DLD-1 or HT-29 cell death were seen (not shown). Next, we investigated the mechanism/s underlying this effect, and focused our attention on signal transducer and activator of transcription 3 (STAT3) and nuclear factor-kappa B (NF-kB), two transcription factors whose activation modulates cell proliferation and survival in transformed cells.26 TIL-derived supernatants induced a more pronounced activation of both STAT3 and NF-kB in DLD-1 and HT-29 cells compared with LPMC-derived supernatants (Figure 1b). Immunofluorescence confirmed STAT3 and NF-kB activation in DLD-1 cells and showed a nuclear colocalization of the two activated transcription factors in the presence of TIL-derived supernatants (Figure 1c). Similar results were obtained in HT-29 cells (not shown). Parallel experiments were performed in the presence of BP-1-102, a STAT3 inhibitor that also represses nuclear NF-kB retention both in vitro and in vivo, thereby attenuating NF-kB activation.27 Treatment of DLD-1 and HT-29 cells with BP-1-102 completely suppressed STAT3 and NF-kB activation, as well as the proproliferative effect of TIL-derived supernatants on CRC cells (Figures 1d and e).

TIL-derived supernatants (TIL SNs) increase CRC cell proliferation through activation of STAT3 and NF-kB. (a) TIL SNs, lamina propria mononuclear cell-derived supernatants (LPMC SNs) and RPMI 1640 complete medium (control) (all used at 1:20 final dilution) were added to DLD-1 and HT-29 cell cultures. After 24 h, cell proliferation was assessed by 5-bromodeoxyuridine (BrdU) assay. Data indicate mean±s.e.m. of four independent experiments in which culture supernatants derived from TILs and LPMCs isolated from the tumor area and the macroscopically unaffected, adjacent, colonic mucosa of four patients who had undergone resection for sporadic CRC were used. Differences between groups were compared using one-way analysis of variance (ANOVA) followed by Bonferroni’s post hoc test. DLD-1: TIL SN-stimulated cells vs either LPMC SN-stimulated cells or control, ***P<0.001; HT-29: TIL SN-stimulated cells vs either LPMC SN-stimulated cells or control, ***P<0.001. (b) DLD-1 and HT-29 cells were cultured in the presence of either TIL SNs or LPMC SNs or RPMI 1640 complete medium (control) (all used at 1:20 final dilution) for 15 min. P-STAT3 Tyr705, STAT3, p-NF-kB/p65 Ser536 and NF-kB/p65 expression was assessed by western blotting. β-Actin was used as loading control. Shown is one of four representative experiments in which culture supernatants derived from TILs and LPMCs isolated from the same patients described in (a) were used. (c) Representative immunofluorescence pictures showing activated STAT3 colocalization with activated NF-kB/p65 in DLD-1 cells cultured in the presence of TIL SNs for 15 min. Cells were cultured as indicated in (b), fixed and stained with anti-p-STAT3 Tyr705 antibody and secondary Alexa Fluor 546 antibody (red), anti-p-NF-kB/p65 Ser536 and secondary Alexa Fluor 488 antibody (green) or 4',6-diamidino-2-phenylindole (DAPI) nuclear staining (blue). The scale bars are 10 μm. The figure is representative of three separate experiments in which culture supernatants derived from TILs and LPMCs isolated from the same patients described in (a) were used. (d) Effect of STAT3 inhibitor BP-1-102 on TIL SN-mediated STAT3 and NF-kB activation. Representative western blotting showing p-STAT3 Tyr705, STAT3, p-NF-kB/p65 Ser536 and NF-kB/p65 expression in DLD-1 and HT-29 cells stimulated or not with TIL SNs in the presence or absence of BP-1-102. β-Actin was used as a loading control. Blots are representative of four independent experiments in which culture supernatants derived from TILs isolated from the tumor area of four patients who had undergone resection for sporadic CRC were used. (e) Inhibition of STAT3 and NF-kB activation by BP-1-102 completely suppresses TIL SN-mediated increase of CRC cell proliferation. Representative histograms showing cell proliferation of DLD-1 and HT-29 cells stimulated or not with TIL SNs in the presence or absence of BP-1-102 for 24 h. Data indicate mean±s.e.m. of four experiments in which culture supernatants derived from TILs isolated from the same patients described in (d) were used. Differences between groups were compared using one-way ANOVA followed by Bonferroni’s post hoc test. DLD-1: TIL SN+BP-1-102-treated cells vs TIL SN-treated cells, ***P<0.001; HT-29: TIL SN+BP-1-102-treated cells vs TIL SN-treated cells, ***P<0.001.
CRC is massively infiltrated with T cells producing Th17-related cytokines, TNF-α and IL-6
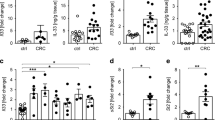
Next, LPMC- and TIL-derived supernatants were analyzed for cytokines known to control cancer cell proliferation. Higher expression of IL-17A, IL-17F, IL-21, IL-22, TNF-α and IL-6 was found in TIL-derived supernatants compared with LPMC-derived supernatants, whereas there was no difference in terms of IFN-γ concentrations (Figure 2a). To ascertain whether these changes in cytokine production reflected accumulation of specific immune cell types in the tumor area, LPMCs and TILs were analyzed by flow cytometry. The percentage of CD3+CD8−, CD3+CD8+, CD3+CD56+, CD3−CD56+, CD19+ and CD68+ cells did not differ between LPMCs and TILs (Figure 2b). However, while TILs and LPMCs contained similar fractions of IFN-γ-producing CD45+ cells, the percentages of IL-17A-, IL-17F-, IL-21-, IL-22-, TNF-α- and IL-6-expressing CD45+ cells were increased in TILs (Figure 2c). Further analysis showed that, in both LPMCs and TILs, IFN-γ was mainly produced by CD3+CD8− and CD3+CD8+ cells (Supplementary Figure 1). Lower production of IFN-γ by CD3+CD8− cells and higher production of IFN-γ derived from CD3+CD8+ cells was seen in TILs as compared with LPMCs (Supplementary Figure 1). IL-17A, IL-17F, IL-21, IL-22 and IL-6 were almost totally derived from CD3+CD8− cells. TNF-α was also produced mainly by CD3+CD8− cells and, to a lesser extent, by CD3+CD8+ cells in both LPMCs and TILs (Supplementary Figure 1). As the majority of IFN-γ, Th17-related cytokine-, TNF-α- and IL-6-producing CD45+ cells in both TILs and LPMCs were CD3+CD8− and CD3+CD8+ cells, we restricted our further analysis to these cell types. The percentage of IFN-γ-producing CD3+CD8− cells was not different between TILs and LPMCs (Figure 3a). In contrast, the percentage of CD3+CD8− cells expressing IL-17A, IL-17F, IL-21, IL-22, TNF-α and IL-6 was increased in TILs (Figure 3a). Next, we assessed the expression of T-bet and receptor-related orphan receptor gamma t (RORγt) in CD3+CD8− cells in TILs and LPMCs. The majority of CD3+CD8− cells coexpressed T-bet and RORγt both in TILs and LPMCs (Figure 3b). In contrast, TIL samples contained more T-bet-/RORγt+ CD3+CD8− cells and less T-bet+/RORγt− CD3+CD8− cells than LPMC samples (Figure 3b). As expected, T-bet+/RORγt− CD3+CD8− cells produced IFN-γ but not IL-17A, whereas T-bet−/RORγt+ CD3+CD8− cells produced IL-17A but not IFN-γ. Double-positive T-bet/RORγt CD3+CD8− cells coexpressed both IFN-γ and IL-17A, whereas double-negative T-bet/RORγt CD3+CD8− cells produced neither IFN-γ nor IL-17A (Figure 3c). Further analysis of cytokine production revealed that the double-positive T-bet/RORγt CD3+CD8− cells were the major source of IFN-γ, Th17-related cytokines, TNF-α and IL-6 in both TILs and LPMCs (Figure 3d). However, TILs contained higher fraction of T-bet−/RORγt+ CD3+CD8− cells producing Th17 cytokines, TNF-α and IL-6, as compared with LPMCs (Figure 3d). A higher fraction of IFN-γ- and TNF-α-producing CD3+CD8+ cells was also observed in TILs as compared with LPMCs, whereas IL-17A-, IL-17F-, IL-21-, IL-22- and IL-6-producing CD3+CD8+ cells were barely detectable in both TILs and LPMCs (Supplementary Figure 2A). The majority of CD3+CD8+ cells in both TILs and LPMCs coexpressed T-bet and RORγt (Supplementary Figure 2B). In contrast, TIL samples contained more T-bet+/RORγt+ CD3+CD8+ cells and less T-bet+/RORγt− CD3+CD8+ cells than LPMC samples (Supplementary Figure 2B). Analysis of cytokine production revealed that double-positive T-bet/RORγt CD3+CD8+ cells were the major source of IFN-γ and TNF-α in both TILs and LPMCs (Supplementary Figure 2C). Altogether, these findings show that transition from the uninvolved colonic mucosa to the neoplastic area is marked by a shift in the CD3+CD8− and CD3+CD8+ cell phenotype leading to the accumulation of Th17-related cytokines, TNF-α and IL-6.

The neoplastic area of CRC samples is massively infiltrated with Th17-related cytokine-, TNF-α- and IL-6-producing cells. (a) IFN-γ, IL-17A, IL-17F, IL-21, IL-22, TNF-α and IL-6 proteins were analyzed by enzyme-linked immunosorbent assay (ELISA) in LPMC-derived supernatants (LPMC SNs) and TIL-derived supernatants (TL SNs), and data are expressed as pg/ml supernatants. Data indicate mean±s.e.m. of six experiments. Differences were calculated using the two-tailed Student’s t-test. (b) Characterization of immune cell infiltrate in LPMCs and TIsL. Representative histograms showing the fraction of T cells, natural killer T (NKT) cells, NK cells, macrophages and B cells in LPMCs and TILs isolated from adjacent tumor and non-tumor areas of 14 patients undergoing colectomy for sporadic CRC. CD45+ cells were gated and analyzed for the indicated markers. Data are expressed as mean±s.e.m. and differences were calculated using the two-tailed Student’s t-test. (c) Representative histograms showing the fraction of IFN-γ-, IL-17A-, IL-17F-, IL-21-, IL-22-, TNF-α- and IL-6-expressing CD45+ cells in LPMCs and TILs isolated from adjacent tumor and non-tumor areas of 14 patients undergoing colectomy for sporadic CRC. Data are expressed as mean±s.e.m. and differences were calculated using the two-tailed Student’s t-test.

Analysis of cytokine production and T-bet/RORγt expression in LPMC- and TIL-derived CD3+CD8− subsets. (a) Representative histograms showing the fraction of IFN-γ-, IL-17A-, IL-17F-, IL-21-, IL-22-, TNF-α- and IL-6-expressing CD3+CD8− cells in LPMCs and TILs isolated from adjacent tumor and non-tumor areas of 14 patients undergoing colectomy for sporadic CRC. Data are expressed as mean±s.e.m. and differences were calculated using the two-tailed Student’s t-test. Right insets. Representative dot plots showing the percentage of IFN-γ- and/or IL-21-, IL-17A- and/or IL-17F-, IL-22- and/or IL-6- and TNF-α-producing CD3+CD8− cells in LPMCs and TILs. The numbers indicate the percentage of cells in the designated quadrants. (b) Representative histograms showing the fraction of T-bet+ and/or Rorγt+ CD3+CD8− cells in LPMCs and TILs isolated from adjacent tumor and non-tumor areas of 14 patients undergoing colectomy for sporadic CRC. Data are expressed as mean±s.e.m. and differences were calculated using the two-tailed Student’s t-test. Right insets. Representative dot plots showing the percentage of T-bet+ and/or Rorγt+ CD3+CD8− cells in LPMCs and TILs. The numbers indicate the percentage of cells in the designated quadrants. Staining of LPMCs with APC- and PE-Cy7-conjugated control isotype IgG is also shown. (c) Representative dot plots showing the ability to produce IFN-γ and/or IL-17A by the indicated subsets of TILs. The numbers indicate the percentage of cells in the designated quadrants. (d) Representative histograms showing the percentage of IFN-γ-, IL-17A-, IL-17F-, IL-21-, IL-22-, TNF-α- and IL-6-producing CD3+CD8− cells in LPMCs and TILs isolated from adjacent tumor and non-tumor areas of one patient undergoing colectomy for sporadic CRC . IFN-γ-, IL-17A-, IL-17F-, IL-21-, IL-22-, TNF-α- and IL-6-producing CD3+CD8− cells were gated and analyzed for the indicated markers. The example is representative of 10 independent experiments in which cells isolated from 10 patients undergoing colectomy for sporadic CRC were analyzed.
IL-17A, IL-22, TNF-α and IL-6 activate STAT3 and NF-kB and contribute to the TIL-derived culture supernatant-mediated CRC cell proliferation
IL-22 and IL-6 activated STAT3 in DLD-1 and HT-29 cells, whereas activation of NF-kB was seen in cells stimulated with IL-17A or TNF-α. In contrast, no activation of STAT3 and NF-kB was seen in cells stimulated with IL-17F or IL-21 (Figure 4a). Both these cytokines were however biologically active, as IL-17F induced IL-6 in stromal cell cultures and IL-21 activated STAT3 in human LPMCs (not shown). To investigate the contribution of Th17-related cytokines, TNF-α and IL-6, in TIL-derived supernatant-driven CRC cell growth, supernatants were preincubated with neutralizing anti-cytokine antibodies before adding to CRC cell lines. Initially, we verified the functional activity of each neutralizing antibody by testing its ability to suppress the function of the corresponding cytokine in cultures of CRC cells (Supplementary Figure 3). Blockade of IL-6 partially reduced p-STAT3 Y705, but not p-NF-kB/p65 Ser536, in DLD-1 and HT-29 cells cultured with TIL-derived supernatants. In contrast, individual neutralization of IL-17A, IL-17F, IL-21, IL-22 or TNF-α did not change TIL-derived supernatant-driven STAT3 and NF-kB activation (Figure 4b). Moreover, no significant change in TIL-derived supernatant-driven CRC cell growth was seen by the blockade of a single cytokine (Figure 4c). Next, we assessed the effect of neutralization of multiple cytokines on STAT3 and NF-kB activation as well as on the promitogenic effect of TIL-derived supernatants. Neutralization of both IL-17A and TNF-α did not alter activation of STAT3 and NF-kB nor TIL-derived supernatant-induced CRC cell proliferation (Figures 4d and e). Both anti-IL-22 and anti-IL-6 reduced TIL-derived supernatant-driven p-STAT3 Y705 but not p-NF-kB/p65 Ser536, and this was associated with a decreased CRC cell growth (Figures 4d and e). Simultaneous neutralization of IL-17A, TNF-α, IL-22 and IL-6 reduced both STAT3 and NF-kB activation and significantly inhibited growth of cells induced by TIL-derived supernatants compared with cells preincubated with anti-IL-22 and anti-IL-6 (Figures 4d and e). No change in cell viability was observed, thus ruling out the possibility that changes in cell proliferation were secondary to induction of cell death (data not shown).

IL-17A, IL-22, TNF-α and IL-6 contribute to TIL-derived supernatant (TIL SN)-mediated STAT3/NF-kB activation and mitogenic effect in CRC cells. (a) Representative western blotting showing p-STAT3 Tyr705, STAT3, p-NF-kB/p65 Ser536 and NF-kB/p65 expression in DLD-1 and HT-29 cells stimulated or not with IL-17A, IL-17F, IL-21, IL-22, TNF-α- and IL-6 (all used at 25 ng/ml) for 15 min. β-Actin was used as a loading control. One of three representative experiments in which similar results were obtained is shown. (b) Representative western blotting showing p-STAT3 Tyr705, STAT3, p-p65 Ser536 and NF-kB/p65 expression in DLD-1 and HT-29 cells stimulated or not with TIL SNs in the presence or absence of anti-IL-17A, anti-IL-17F, anti-IL-21, anti-IL-22, anti-TNF-α and anti-IL-6 (all used at 10 μg/ml) as indicated. β-Actin was used as a loading control. One of three representative experiments in which similar results were obtained is shown. (c) Representative histograms showing cell proliferation of DLD-1 and HT-29 cells stimulated as indicated in (b). Data indicate mean±s.e.m. of four experiments. Differences between groups were compared using one-way analysis of variance (ANOVA) followed by Bonferroni’s post hoc test. (d) Representative western blotting showing p-STAT3 Tyr705, STAT3, p-NF-kB/p65 Ser536 and NF-kB/p65 expression in DLD-1 and HT-29 cells stimulated or not with TIL SNs in the presence or absence of anti-IL-17A, anti-IL-22, anti-TNF-α and anti-IL-6, used in combination as indicated. β-Actin was used as a loading control. One of three representative experiments in which similar results were obtained is shown. (e) Representative histograms showing cell proliferation of DLD-1 and HT-29 cells stimulated as indicated in (d). Data indicate mean±s.e.m. of five experiments. Differences between groups were compared using one-way analysis of variance (ANOVA) followed by Bonferroni’s post hoc test. DLD-1: TIL SN+anti-IL-22+anti-IL-6- vs TIL SN-treated cells, *P<0.05, TIL SN+anti-IL-17A+anti-TNF-α+anti-IL-22+anti-IL-6- vs TIL SN-treated cells, ***P<0.001; HT-29: TIL SN+anti-IL-22+anti-IL-6- vs TIL SN-treated cells, **P<0.01, TIL SN+anti-IL-17A+anti-TNF-α+anti-IL-22+anti-IL-6- vs TIL SN-treated cells, ***P<0.001.
Oral BP-1-102 reduces colon tumorigenesis in a mouse model of sporadic CRC
To link mechanistically the activation of STAT3 and NF-kB with CRC cell growth in vivo, we used multiple intestinal neoplasia (min) mice carrying a mutation in the adenomatous polyposis coli (Apc) gene, which spontaneously develop intestinal tumors. Treatment of Apcmin/+ mice with the carcinogen azoxymethane (AOM) increased tumor incidence, number and size, particularly in the colon.28 First, we examined the activation status of STAT3 and NF-kB and the expression of IFN-γ, Th17-related cytokines, TNF-α and IL-6 in early colonic lesions of Apcmin/+ mice. Some animals were treated intraperitoneally with AOM (10 mg/kg) once a week for 2 weeks and killed on day 21 (Supplementary Figure 4A). Both STAT3 and NF-kB were activated in early colonic lesions of Apcmin/+ mice (Supplementary Figures 4B–D). Microscopic examination of colonic sections revealed the accumulation of cells positive for p-STAT3 Y705 and p-NF-kB/p65 Ser536 in both transformed epithelial cells and tumor-infiltrating cells (Supplementary Figures 4C and D). STAT3 and NF-kB activation in early colonic lesions of Apcmin/+ mice was associated with elevated levels of transcripts for IL-17A, IL-21, IL-22, TNF-α and IL-6, whereas IFN-γ and IL-17F RNA transcripts remained unchanged (Supplementary Figure 5). Early colonic lesions of Apcmin/+ mice also exhibited elevated RNA levels of IL-11, a cytokine produced by cancer-associated fibroblasts and myeloid cells and involved in CRC progression (Supplementary Figure 5).29 Phenotypic analysis of immune cells in TILs isolated from the tumors of Apcmin/+ mice showed a preferential accumulation of CD3+CD8− cells in line with the human data (Supplementary Figure 6).
Next, we tested whether BP-1-102 could inhibit intestinal tumorigenesis in Apcmin/+ mice. Mice were treated intraperitoneally with AOM (10 mg/kg) once a week for 2 weeks. Two weeks after the last AOM injection, mice were randomly divided into two groups and given either BP-1-102 (in 1% dimethyl sulfoxide (DMSO) in phosphate-buffered saline (PBS)) or 1% DMSO in PBS (control) three times a week by oral gavage until being killed (day 56) (Figure 5a). Three control mice had to be killed before the end of the scheduled treatment, because of intestinal occlusion caused by large colonic tumors, whereas all mice treated with BP-1-102 survived until the end of the study.

Orally administered BP-1-102 reduces colonic tumorigenesis in Apcmin/+ mice. (a) Experimental protocol used to assess the effect of BP-1-102 treatment on colonic tumorigenesis in Apcmin/+ mice. (b) Upper panels show representative endoscopic pictures of colon tumors developed in mice treated with either dimethyl sulfoxide (DMSO) (CTR) or BP-1-102. Graphs show the number of lesions and the endoscopic scoring of tumors developed in mice treated with either DMSO (CTR) or BP-1-102. Data indicate mean±s.e.m. of three independent experiments in which at least four mice per group were considered. Differences were calculated using the two-tailed Student's t-test. (c) Representative images showing proliferating cell nuclear antigen (PCNA) immunostaining in colonic sections taken from Apcmin/+ mice treated with either DMSO (CTR) or BP-1-102. The scale bars are 20 μm. One of six representative experiments in which similar results were obtained is shown. NT, non-tumor area; T, tumor area.
Endoscopy on day 54 showed that control mice developed multiple large tumors, whereas the number and size of tumors were reduced in the colon of BP-1-102-treated mice (Figure 5b). These results were confirmed by direct assessment of tumors in mice killed on day 56 (unpublished data). Proliferating cell nuclear antigen staining confirmed the antiproliferative effect of BP-1-102 (Figure 5c). By contrast, there was no significant change in proliferating cell nuclear antigen staining in the normal colonic mucosa of mice treated with BP-1-102 (Figure 5c). To determine whether the reduced tumorigenesis observed in BP-1-102-treated mice was associated with a reduced activation of STAT3 and/or NF-kB, we compared p-STAT3 Y705 and p-NF-kB/p65 Ser536 expression in non-tumor and tumor tissue derived from colonic extracts of control and BP-1-102-treated mice killed on day 56. Robust activation of both STAT3 and NF-kB was seen in the neoplastic areas of control mice and inhibited by BP-1-102 treatment, whereas p-STAT3 Y705 and p-NF-kB/p65 Ser536 expression was barely detectable in non-tumor areas of both control and BP-1-102-treated mice (Supplementary Figure 7). Immunohistochemistry confirmed reduced activation of both STAT3 and NF-kB in colonic tumor sections of mice treated with BP-1-102 (Figure 6a). IL-17A, IL-21, IL-22, TNF-α and IL-6 transcripts were reduced in the tumor areas of BP-1-102-treated mice, whereas IFN-γ and IL-17F RNA expression remained unchanged (Figure 6b). Moreover, treatment of mice with BP-1-102 did not change IL-11 transcripts in the tumors as well as the fractions of immune cells in TILs (not shown).

Reduced STAT3/NF-kB activation and reduced expression of IL-17A, IL-21, IL-22, TNF-α and IL-6 are seen in the colonic tumors of BP-1-102-treated Apcmin/+ mice. (a) Representative images showing p-STAT3 Tyr705- or p-NF-kB/p65 Ser536-positive cells in colonic sections taken from the tumor areas Apcmin/+ mice treated with either dimethyl sulfoxide (DMSO) (CTR) or BP-1-102 and killed on day 56. The scale bars are 20 μm. One of six representative experiments in which similar results were obtained is shown. Right insets. Quantification of p-STAT3 Tyr705- or p-NF-kB/p65 Ser536-positive infiltrating and epithelial cells in colonic sections taken from the tumor areas of Apcmin/+ mice treated with either DMSO (CTR) or BP-1-102 and killed on day 56. Data are presented as mean values of positive cells per high power field (h.p.f.)±s.e.m. of three independent experiments in which two sections per group were analyzed. Differences were calculated using the two-tailed Student's t-test. NT, non-tumor area; T, tumor area. (b) IFN-γ, IL-17A, IL-17F, IL-21, IL-22, TNF-α- and IL-6 expression was assessed by real-time PCR in colonic tissues taken from Apcmin/+ mice treated with either DMSO (CTR) or BP-1-102 and killed on day 56. Values are mean±s.e.m. of two independent experiments containing at least three mice per group. Differences were calculated using the two-tailed Student's t-test. ND, not detectable; NT, non-tumor area; T, tumor area.
Discussion
This study was undertaken to characterize the immune/inflammatory infiltrate in sporadic CRC and to clarify which factors released by TILs modulate CRC cell growth and survival. Initially, we showed that TIL-derived supernatants, but not LPMC-derived supernatants, increased proliferation of DLD-1 and HT-29 cells and this was associated with the activation of STAT3 and NF-kB, two transcription factors involved in the interplay between malignant cells and inflammatory/immune cells. To ascertain the role of STAT3 and NF-kB activation in the proproliferative effect mediated by TIL-derived supernatant, DLD-1 and HT-29 cells were treated with BP-1-102, which disrupts STAT3 dimerization, thereby inhibiting its activation.27 BP-1-102 also represses NF-kB transcriptional functions as well as STAT3-NF-kB cross-talk in human breast and lung cancer cells.27 Indeed, preincubation of DLD-1 and HT-29 cells with BP-1-102 suppressed STAT3 and NF-kB activation and abrogated TIL-derived supernatant-induced cell growth.
The ability of immune/inflammatory cells to control cancer cell proliferation is mostly dependent on cytokines, which directly target CRC cells. Our results indicate that TILs produce elevated levels of IL-17A, IL-17F, IL-21, IL-22, TNF-α and IL-6. Characterization of immune cell complexity of sporadic CRC and adjacent normal colonic tissues revealed no changes in the percentages of different cell subsets and suggested that changes in the features of specific immune cell subsets might account for the increased production of Th17-related cytokines, TNF-α and IL-6.
Following triggering of their T-cell receptor and in the presence of appropriate costimulatory signals and specific cytokines, naive CD4+ lymphocytes differentiate into various effector or regulatory cells characterized by distinct functions and specific cytokine production profiles.30 Terminally differentiated Th1 cells express the transcription factor T-bet and produce IFN-γ,31 whereas Th17 cells express RORγt and produce IL-17A, IL-17F, IL-21 and IL-22.32,33 Flow cytometry analysis revealed that T-bet-/RORγt+ Th cells producing IL-17A, IL-17F, IL-21, IL-22, TNF-α and IL-6 accumulate in TILs. Interestingly, the majority of Th cells in both TILs and LPMCs coexpressed T-bet and RORγt. However, double-positive T-bet/RORγt Th cells in TILs produced higher amount of IL-17A, IL-17F, IL-21, IL-22, TNF-α and IL-6 than those in LPMCs. CD8+ T cells have a crucial role in tumor immunity resulting in part from the release of different cytotoxic molecules (for example, granzyme B, perforin). However, besides cytotoxic activity, CD8+ T cells expressing RORγt have been reported to produce Th17-related cytokines that sustain tumor growth.34 Flow cytometry analysis revealed that double-positive T-bet/RORγt CD8+ cells, which produce high levels of IFN-γ and TNF-α, accumulate in TILs. In contrast, the production of IL-17A, IL-17F, IL-21, IL-22 and IL-6 was barely detectable in CD8+ cells in both TILs and LPMCs. Although cytokines in the tumor microenvironment are produced mostly by hematopoietic cells, some of the cytokines such as IL-6 and TNF-α can also be produced by the malignant cells themselves to establish an autocrine tumor-promoting signaling loop that further enhances NF-kB and STAT3 activation.26 However, DLD-1 and HT-29 cells did not produce IL-6 and TNF-α in the presence or absence of TIL-derived supernatants (unpublished observations).
In subsequent studies, we tested the ability of each of the cytokines overproduced by TILs to activate STAT3 and/or NF-kB in DLD-1 and HT-29 cells. IL-22 and IL-6 activated STAT3 in both cells lines, whereas activation of NF-kB was seen in cells stimulated with IL-17A or TNF-α. These results are consistent with the notion that STAT3 as well as NF-kB activators are redundant in cancer cells and fit with our demonstration that single neutralization of IL-17A, IL-22, TNF-α and IL-6 did not affect the ability of TIL-derived supernatants to activate STAT3 and NF-kB and promote CRC cell proliferation. Combined neutralization of IL-22 and IL-6 reduced but did not abrogate TIL supernatant-induced STAT3 activation and CRC cell proliferation, probably because of the presence of additional STAT3-activating cytokines/factors in TIL-derived supernatants (for example, IL-11 or EGF). In contrast, combined neutralization of IL-17A and TNF-α only slightly affected NF-kB activation and the proproliferative effect of TIL-derived supernatants, and this again could be related to the presence of other NF-kB-activating cytokines/factors (for example, IL-1β) in TIL-derived supernatants. Another possibility is that the simultaneous activation of STAT3 in these cells could sustain NF-kB activity as it was demonstrated that STAT3 prolongs NF-kB nuclear retention through acetyltransferase p300-mediated p65 acetylation.35 Interestingly, simultaneous inhibition of IL-17A, TNF-α, IL-22 and IL-6 almost completely blocked STAT3 and NF-kB activation and the increased CRC cell growth induced by TIL-derived supernatants. These results collectively indicate that TILs secrete multiple cytokines that exert mitogenic effects because of their ability to trigger simultaneously STAT3 and NF-kB in CRC cells. Our data also confirm that inhibition of STAT3 is much more important than inhibition of NF-kB in the negative control of CRC cell growth.4,36,37
Our in vitro results were supported by studies in Apcmin/+ mice showing that inhibition of STAT3 and NF-kB with BP-1-102 reduced the growth of colonic tumors. Although BP-1-102 is a specific inhibitor of STAT3 activity, BP-1-102-mediated downregulation of NF-kB activation relies on the demonstration that STAT3 interacts with and sustains NF-kB activity.35 STAT3 and NF-kB activation was seen in both tumor cells and tumor-infiltrating immune cells in early colonic lesions of Apcmin/+ mice and both transcription factors can regulate not only the cancer cell behavior but also the production of cytokines and other inflammatory mediators by immune cells.38,39 Mice given BP-1-102 exhibited reduced levels of proinflammatory cytokines in the tumor areas, probably based on the fact that, in immune cells, the activity of STAT3 either directly or indirectly controls the production of Th17-related cytokines, IL-6 and TNF-α.40, 41, 42
Because STAT3 is required for the survival of intestinal epithelial cells and maintenance of mucosal integrity,4 excessive interference with systemic STAT3 activation could potentially cause gastrointestinal damage. However, we show here that BP-1-102 inhibited the proliferation of neoplastic cells but not normal colonic crypt cells, and did not affect normal mucosal regeneration processes. Thus, it is tempting to speculate that BP-1-102 interferes exclusively with the excessive STAT3 and NF-kB activation that sustains colon carcinogenesis.
Taken together, our data show that transition from the uninvolved colonic mucosa to the neoplastic area is marked by a shift in the Th program leading to the accumulation of IL-17A, IL-22, TNF-α and IL-6 that ultimately sustain CRC cell growth through the activation of STAT3 and NF-kB. Although reagents that inhibit the above-mentioned cytokines are already available and most have been found to be safe and effective for treatment of autoimmune disorders,43 our data suggest that single neutralization of STAT3- and/or NF-kB-activating cytokines may not be effective owing to the redundant effects of such molecules on target cells.
In this context, compounds inhibiting simultaneously STAT3 and NF-kB (for example, BP-1-102) might help combat CRC, given that these drugs not only affect growth of malignant cells but also negatively regulate production of protumorigenic cytokines by tumor-infiltrating cells.
Materials and methods
Patients and samples
Paired tissue samples were taken from the tumor area and the macroscopically unaffected, adjacent, colonic mucosa of 14 patients who underwent colon resection for sporadic CRC (all with TNM stages II–III) at the Tor Vergata University Hospital (Rome, Italy). These samples were used to extract TILs and LPMCs as described below. No patients received radiotherapy or chemotherapy before undergoing surgery. The human studies were approved by the local ethics committee and each patient gave written informed consent.
Animals
Apcmin/+ mice were obtained from the Jackson Laboratory (Bar Harbor, ME, USA) and maintained in filter-topped cages on autoclaved food and water at the Plaisant animal facility (Rome, Italy). Mice were routinely tested (every 3 months) for health status and infections according to the Federation of European Laboratory Animal Science Associations (FELASA) guidelines. Mice were negative for all pathogens included in this protocol. Mice were also negative for Helicobacter hepaticus and Helicobacter bilis. All animal experiments were approved by the local Institutional Animal Care and Use Committee.
Isolation of LPMCs and TILs from human colonic samples
TILs and LPMCs were isolated from CRC samples and adjacent, normal, colonic mucosa using dithiothreitol–ethylenediaminetetraacetic acid and collagenase method. Briefly, pieces of intestinal mucosa and tumor tissue were dissected from surgical specimens within 1 h of resection and washed in Hank’s balanced salt solution containing 1 mM dithiothreitol and antibiotics for 15 min at room temperature to remove mucus. Samples were then minced and incubated in Hank’s balanced salt solution containing 1 mM ethylenediaminetetraacetic acid and antibiotics for 45 min at 37 °C to remove epithelial cells. After two washes in Hank’s balanced salt solution, samples were incubated in type D collagenase (0.75 mg/ml; Roche Diagnostic, Monza, Italy) for 1 h at 37 °C. After collagenase digestion, media containing the mononuclear cells were collected and washed two times in Hank’s balanced salt solution. Subsequently, the pellets were resuspended in RPMI 1640 and then layered on a Percoll density gradient as described previously44 to isolate either LPMCs or TILs. The isolated cells were counted and checked for viability using 0.1% trypan blue (viability ranged from 89 to 97%).
Assessment of cell proliferation and death
Cell proliferation was assessed by using a commercially available 5-bromodeoxyuridine assay kit (Roche Diagnostic) and confirmed by flow cytometry using carboxyfluorescein diacetate succinimidyl ester (Life Technologies, Milan, Italy) as described previously.45 The percentage of cell death was evaluated by flow cytometry at the same time point of cell proliferation after staining with propidium iodide.
Cell cultures
All reagents were from Sigma-Aldrich (Milan, Italy) unless specified. The human CRC cell lines DLD-1 and HT-29 were obtained from the American Type Culture Collection (ATCC) and maintained in RPMI 1640 and McCoy’s 5A medium, respectively, all supplemented with 10% fetal bovine serum, 1% penicillin/streptomycin (all from Lonza, Verviers, Belgium), and 50 μg/ml gentamicin in a 37 °C, 5% CO2, fully humidified incubator. Cell lines have been recently authenticated by STR DNA fingerprinting using the PowerPlex 18D System Kit (Promega, Milan, Italy) according to the manufacturer’s instructions. The STR profiles of all the cell lines matched the known DNA fingerprints.
Freshly isolated TILs and LPMCs were resuspended in complete RPMI 1640 medium. One million LPMCs or TILs were cultured in 48-well plates to produce 0.5 ml of supernatant. Cell-free supernatants were harvested after 48 h. To characterize the immune cell subsets and their cytokine production, an aliquot of cells was cultured in the presence of phorbol 12-myristate 13-acetate (80 pM), ionomycin (1 mg/ml) and monensin (2 μM; eBioscience, San Diego, CA, USA) for 5 h, and then assessed by flow cytometry following staining with specific fluorochrome-conjugated antibodies.
To examine whether LPMC- and TIL-derived supernatants affect CRC cell proliferation and/or viability, DLD-1 and HT-29 cells were cultured in the presence of either LPMC- or TIL-derived supernatants or RPMI 1640 medium (vehicle) (all used at 1:20 final dilution) for 24 h.
To assess whether LPMC- and TIL-derived supernatants activate STAT3 and/or NF-kB in CRC cells, DLD-1 and HT-29 cells were cultured in the presence of either LPMC- or TIL-derived supernatants or RPMI 1640 medium (vehicle) (all used at 1:20 final dilution) for 15 min.
To investigate whether the TIL-derived supernatant-mediated increase of CRC cell proliferation relies on STAT3 and NF-kB activation, DLD-1 and HT-29 cells were preincubated either with BP-1-102 (1 and 3 μg/ml, respectively) or DMSO (vehicle) for 1 h and then stimulated either with TIL-derived supernatants or RPMI 1640 medium (vehicle) (both used at 1:20 final dilution). After 15 min, STAT3 and NF-kB activation was evaluated by western blotting. Cell proliferation was assessed after 24 h by 5-bromodeoxyuridine assay.
To determine whether Th17-related cytokines, TNF-α and IL-6 activate STAT3 and/or NF-kB in DLD-1 and HT-29 cells, cells were stimulated with recombinant human IL-17A (Peprotech, London, UK), IL-17F (Peprotech), IL-21 (Life Technologies), IL-22 (R&D Systems, Minneapolis, MN, USA), IL-6 (Peprotech) or TNF-α (R&D Systems) (all used at 25 ng/ml) for 15 min. STAT3 and NF-kB activation was assessed by western blotting.
To assess the contribution of each Th17-related cytokines, TNF-α and IL-6 in TIL-derived supernatant-mediated increase of CRC cell proliferation, TIL-derived supernatants were preincubated with specific antibodies neutralizing IL-17A (anti-IL-17A; R&D Systems), IL-17F (anti-IL-17F; Acris Antibodies GmbH, Herford, Germany), IL-21 (anti-IL-21; Giuliani SpA, Milan, Italy), IL-22 (anti-IL-22; R&D Systems), TNF-α (anti-TNF-α; R&D Systems) and IL-6 (anti-IL-6; R&D Systems) (all used at 10 μg/ml) and then added to DLD-1 and HT-29 cells. After 15 min, STAT3 and NF-kB activation was evaluated by western blotting. Cell proliferation was assessed after 24 h by 5-bromodeoxyuridine.
Western blotting
Total proteins were extracted from human CRC cells or mouse colonic tissues as described elsewhere,45 and separated on sodium dodecyl sulfate–polyacrylamide gel electrophoresis gel. Blots were incubated with p-STAT3 Tyr705, p-NF-κB/p65 Ser536 (both from Cell Signaling, Danvers, MA, USA), STAT3 and p65 antibodies (both from Santa Cruz Biotechnology, Santa Cruz, CA, USA). To ascertain equivalent loading of the lanes, blots were stripped and incubated with an anti-β-actin antibody.
Immunofluorescence
CRC cells were grown on glass coverslips in multiwell plates and cultured in the presence of either LPMC- or TIL-derived supernatants or RPMI 1640 medium (vehicle) (all used at 1:20 final dilution) for 15 min. Cells were fixed with ice-cold 70% ethanol for 15 min and permeabilized with 0.2% Triton X-100 for 10 min. Cells were then washed with PBS, blocked in 1% bovine serum albumin for 30 min and incubated with anti-p-STAT3 Tyr705 (Santa Cruz Biotechnology) or anti-p-NF-κB/p65 Ser536 antibody (Cell Signaling) in 0.1% bovine serum albumin at 4 °C overnight. Subsequently, cells were rinsed with PBS and incubated with Alexa Fluor 546 (goat anti-mouse) and Alexa Fluor 488 (donkey anti-rabbit) (both from Life Technologies) for p-STAT3 Tyr705 and p-NF-κB/p65 Ser536 detection, respectively, for 1 h at room temperature in the dark. Specimens were then washed with PBS, counterstained with 4′,6-diamidino-2-phenylindole (Life Technologies), mounted and examined using a fluorescence microscope (Olympus, Milan, Italy).
Analysis of cytokine expression in LPMC- and TIL-derived supernatants
Freshly isolated TILs and LPMCs were resuspended and cultured in complete RPMI 1640 medium and the cell-free supernatants were harvested after 48 h. IFN-γ, IL-17A, IL-17F, IL-22, TNF-α and IL-6 concentrations were measured by enzyme-linked immunosorbent assay according to the manufacturer’s protocol (R&D Systems). IL-21 protein concentrations were measured by enzyme-linked immunosorbent assay according to the manufacturer’s protocol (eBioscience).
Flow cytometry analysis
Flow cytometry analysis was performed by using a BD FACSVerse (BD Biosciences, Milan, Italy) and the following monoclonal anti-human antibodies: CD45-APC-H7, CD3ɛ-PerCP, CD3ɛ-FITC, CD8α-V500, CD56-V450, CD56-PE-Cy7, CD19-FITC, IFN-γ-FITC, IFN-γ-PE, IL-17A-V450, IL-17F-PE, TNFα-PE (all from BD Pharmingen, Milan, Italy), IL-21-PE, IL-22-APC, IL-6-PerCP, T-bet-PE-Cy7 (clone 4B10), RORγt-APC (clone AFKJS-9), RORγt-PE (clone AFKJS-9) (all from eBioscience) and CD68 (BioLegend, San Diego, CA, USA). TILs isolated from mouse colon tumors were stained with the following antibodies: CD3ɛ-Pacific Blue, CD8α-FITC, CD45-APC-Cy7, CD49b-PE (clone DX5), CD19-APC (all from BD Pharmingen) and F4/80-Pacific Blue (BioLegend). In parallel, cells were stained with the respective control isotype antibodies.
Experimental model of sporadic CRC
Cohoused 6- to 7-week-old female Apcmin/+ mice received intraperitoneal injections of 10 mg/kg AOM once a week for 2 weeks to increase colon tumorigenesis as previously reported.28 Two weeks after the last AOM injection, some mice were killed and tumor and non-tumor colonic tissues collected to perform real-time PCR, western blotting and histochemistry experiments. The remaining mice were randomly divided into two groups and given either 5 mg/kg BP-1-102 (in 1% DMSO in PBS) or 1% DMSO in PBS (control) every 2 days by oral gavage (100 μl) until being killed on day 56. The dose of BP-1-102 was selected in accordance with a previous report by Zhang et al.27 Colonoscopy was performed in a blinded manner for monitoring of tumorigenesis using the Coloview high-resolution mouse endoscopic system (Karl-Storz, Tuttlingen, Germany). Tumors observed during endoscopy were counted to obtain the overall number of lesions. Tumor sizes of all tumors in a given mouse were scored using the protocol described by Becker et al.46
Immunohistochemistry
Colonic cryosections of Apcmin/+ mice were stained with p-STAT3 Tyr705 and p-NF-κB/p65 Ser536 (both from Cell Signaling). Isotype control-stained sections were prepared under identical conditions replacing the primary antibody with a rabbit IgG control antibody (R&D Systems). Proliferating cells were evaluated using a PCNA Staining Kit (Zymed Laboratories, Carlsbad, CA, USA) according to the manufacturer’s instructions. Negative control-stained sections were prepared under identical immunohistochemical conditions omitting the primary antibody.
Isolation of TILs from mouse colon tumors
TILs were isolated from colon tumors of Apcmin/+ mice as previousy described.10
RNA extraction, cDNA preparation and real-time PCR
Total RNA was extracted from tumor and non-tumor tissue of Apcmin/+ mice using TRIzol reagent according to the manufacturer’s instructions (Life Technology). Primers used were as follows: IFN-γ—sense, 5′-CAATAGACGCTACACACTGC-3′ and antisense, 5′-CCACATCTATGCCACTTGAG-3′; IL-17A—sense, 5′-TCAGACTACCTCAACCGTTC-3′ and antisense, 5′-TTCAGGACCAGGATCTCTTG-3′; IL-17F—sense, 5′-CCAAAACCAGGGCATTTCTGTC-3′ and antisense, 5′-GAGTTCATGGTGCTGTCTTCC-3′; TNF-α—sense, 5′-ACCCTCACACTCAGATCATC-3′ and antisense, 5′-GAGTAGACAAGGTACAACCC-3′; IL-6—sense, 5′-AGCCAGAGTCCTTCAGAGAG-3′ and antisense, 5′-GATGGTCTTGGTCCTTAGCC-3′; IL-11—sense, 5′-GCTTCCTGGTGTGCTGAC-3′ and antisense, 5′-CCAGGCGAGACATCAAGAG-3′. IL-21 and IL-22 RNA expression was evaluated using a TaqMan assay (Life Technologies). RNA expression was calculated relative to the housekeeping β-actin gene on the base of the ΔΔCt algorithm.
Statistical analysis
Values are expressed as mean±s.e.m. and results were analyzed using the two-tailed Student's t-test for comparison between two groups or one-way analysis of variance followed by Bonferroni’s post hoc test for multiple comparisons. Significance was defined as P-values <0.05.
References
Center MM, Jemal A, Smith RA, Ward E . Worldwide variations in colorectal cancer. CA Cancer J Clin 2009; 59: 366–378.
Fearon ER, Vogelstein B . A genetic model for colorectal tumorigenesis. Cell 1990; 61: 759–767.
Eaden JA, Abrams KR, Mayberry JF . The risk of colorectal cancer in ulcerative colitis: a meta-analysis. Gut 2001; 48: 526–535.
Grivennikov S, Karin E, Terzic J, Mucida D, Yu GY, Vallabhapurapu S et al. IL-6 and Stat3 are required for survival of intestinal epithelial cells and development of colitis-associated cancer. Cancer Cell 2009; 15: 103–113.
Huber S, Gagliani N, Zenewicz LA, Huber FJ, Bosurgi L, Hu B et al. IL-22BP is regulated by the inflammasome and modulates tumorigenesis in the intestine. Nature 2012; 491: 259–263.
Hyun YS, Han DS, Lee AR, Eun CS, Youn J, Kim HY . Role of IL-17A in the development of colitis-associated cancer. Carcinogenesis 2012; 33: 931–936.
Jauch D, Martin M, Schiechl G, Kesselring R, Schlitt HJ, Geissler EK et al. Interleukin 21 controls tumour growth and tumour immunosurveillance in colitis-associated tumorigenesis in mice. Gut 2011; 60: 1678–1686.
Kirchberger S, Royston DJ, Boulard O, Thornton E, Franchini F, Szabady RL et al. Innate lymphoid cells sustain colon cancer through production of interleukin-22 in a mouse model. J Exp Med 2013; 210: 917–931.
Popivanova BK, Kitamura K, Wu Y, Kondo T, Kagaya T, Kaneko S et al. Blocking TNF-alpha in mice reduces colorectal carcinogenesis associated with chronic colitis. J Clin Invest 2008; 118: 560–570.
Stolfi C, Rizzo A, Franze E, Rotondi A, Fantini MC, Sarra M et al. Involvement of interleukin-21 in the regulation of colitis-associated colon cancer. J Exp Med 2011; 208: 2279–2290.
Lanca T, Silva-Santos B . The split nature of tumor-infiltrating leukocytes Implications for cancer surveillance and immunotherapy. Oncoimmunology 2012; 1: 717–725.
Monteleone G, Pallone F, Stolfi C . The dual role of inflammation in colon carcinogenesis. Int J Mol Sci 2012; 13: 11071–11084.
Xu X, Fu XY, Plate J, Chong AS . IFN-gamma induces cell growth inhibition by Fas-mediated apoptosis: requirement of STAT1 protein for up-regulation of Fas and FasL expression. Cancer Res 1998; 58: 2832–2837.
Senik A, Stefanos S, Kolb JP, Lucero M, Falcoff E . Enhancement of mouse natural killer cell activity by type II interferon. Ann Immunol (Paris) 1980; 131C: 349–361.
Tannenbaum CS, Hamilton TA . Immune-inflammatory mechanisms in IFNgamma-mediated anti-tumor activity. Semin Cancer Biol 2000; 10: 113–123.
Street SE, Trapani JA, MacGregor D, Smyth MJ . Suppression of lymphoma and epithelial malignancies effected by interferon gamma. J Exp Med 2002; 196: 129–134.
Galon J, Costes A, Sanchez-Cabo F, Kirilovsky A, Mlecnik B, Lagorce-Pages C et al. Type, density, and location of immune cells within human colorectal tumors predict clinical outcome. Science 2006; 313: 1960–1964.
Song X, Gao H, Lin Y, Yao Y, Zhu S, Wang J et al. Alterations in the microbiota drive interleukin-17C production from intestinal epithelial cells to promote tumorigenesis. Immunity 2014; 40: 140–152.
Wu P, Wu D, Ni C, Ye J, Chen W, Hu G et al. GammadeltaT17 cells promote the accumulation and expansion of myeloid-derived suppressor cells in human colorectal cancer. Immunity 2014; 40: 785–800.
Kryczek I, Lin Y, Nagarsheth N, Peng D, Zhao L, Zhao E et al. IL-22(+)CD4(+) T cells promote colorectal cancer stemness via STAT3 transcription factor activation and induction of the methyltransferase DOT1L. Immunity 2014; 40: 772–784.
Grivennikov SI, Wang K, Mucida D, Stewart CA, Schnabl B, Jauch D et al. Adenoma-linked barrier defects and microbial products drive IL-23/IL-17-mediated tumour growth. Nature 2012; 491: 254–258.
Chae WJ, Gibson TF, Zelterman D, Hao L, Henegariu O, Bothwell AL . Ablation of IL-17A abrogates progression of spontaneous intestinal tumorigenesis. Proc Natl Acad Sci USA 2010; 107: 5540–5544.
Fridman WH, Galon J, Pages F, Tartour E, Sautes-Fridman C, Kroemer G . Prognostic and predictive impact of intra- and peritumoral immune infiltrates. Cancer Res 2011; 71: 5601–5605.
Fridman WH, Pages F, Sautes-Fridman C, Galon J . The immune contexture in human tumours: impact on clinical outcome. Nat Rev Cancer 2012; 12: 298–306.
Tosolini M, Kirilovsky A, Mlecnik B, Fredriksen T, Mauger S, Bindea G et al. Clinical impact of different classes of infiltrating T cytotoxic and helper cells (Th1, th2, treg, th17) in patients with colorectal cancer. Cancer Res 2011; 71: 1263–1271.
Grivennikov SI, Karin M . Dangerous liaisons: STAT3 and NF-kappaB collaboration and crosstalk in cancer. Cytokine Growth Factor Rev 2010; 21: 11–19.
Zhang X, Yue P, Page BD, Li T, Zhao W, Namanja AT et al. Orally bioavailable small-molecule inhibitor of transcription factor Stat3 regresses human breast and lung cancer xenografts. Proc Natl Acad Sci USA 2012; 109: 9623–9628.
Issa AY, Volate SR, Muga SJ, Nitcheva D, Smith T, Wargovich MJ . Green tea selectively targets initial stages of intestinal carcinogenesis in the AOM-ApcMin mouse model. Carcinogenesis 2007; 28: 1978–1984.
Putoczki TL, Thiem S, Loving A, Busuttil RA, Wilson NJ, Ziegler PK et al. Interleukin-11 is the dominant IL-6 family cytokine during gastrointestinal tumorigenesis and can be targeted therapeutically. Cancer Cell 2013; 24: 257–271.
Yamane H, Paul WE . Early signaling events that underlie fate decisions of naive CD4(+) T cells toward distinct T-helper cell subsets. Immunol Rev 2013; 252: 12–23.
Szabo SJ, Kim ST, Costa GL, Zhang X, Fathman CG, Glimcher LH . A novel transcription factor, T-bet, directs Th1 lineage commitment. Cell 2000; 100: 655–669.
Ivanov II, McKenzie BS, Zhou L, Tadokoro CE, Lepelley A, Lafaille JJ et al. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 2006; 126: 1121–1133.
Korn T, Bettelli E, Oukka M, Kuchroo VK . IL-17 and Th17 Cells. Annu Rev Immunol 2009; 27: 485–517.
Yen HR, Harris TJ, Wada S, Grosso JF, Getnet D, Goldberg MV et al. Tc17 CD8 T cells: functional plasticity and subset diversity. J Immunol 2009; 183: 7161–7168.
Lee H, Herrmann A, Deng JH, Kujawski M, Niu G, Li Z et al. Persistently activated Stat3 maintains constitutive NF-kappaB activity in tumors. Cancer Cell 2009; 15: 283–293.
Greten FR, Eckmann L, Greten TF, Park JM, Li ZW, Egan LJ et al. IKKbeta links inflammation and tumorigenesis in a mouse model of colitis-associated cancer. Cell 2004; 118: 285–296.
Bollrath J, Phesse TJ, von Burstin VA, Putoczki T, Bennecke M, Bateman T et al. Gp130-mediated Stat3 activation in enterocytes regulates cell survival and cell-cycle progression during colitis-associated tumorigenesis. Cancer Cell 2009; 15: 91–102.
Hayden MS, West AP, Ghosh S . NF-kappaB and the immune response. Oncogene 2006; 25: 6758–6780.
Carpenter RL, Lo HW . STAT3 target genes relevant to human cancers. Cancers (Basel) 2014; 6: 897–925.
Yu H, Kortylewski M, Pardoll D . Crosstalk between cancer and immune cells: role of STAT3 in the tumour microenvironment. Nat Rev Immunol 2007; 7: 41–51.
Caprioli F, Sarra M, Caruso R, Stolfi C, Fina D, Sica G et al. Autocrine regulation of IL-21 production in human T lymphocytes. J Immunol 2008; 180: 1800–1807.
Yeste A, Mascanfroni ID, Nadeau M, Burns EJ, Tukpah AM, Santiago A et al. IL-21 induces IL-22 production in CD4+ T cells. Nat Commun 2014; 5: 3753.
Wang K, Grivennikov SI, Karin M . Implications of anti-cytokine therapy in colorectal cancer and autoimmune diseases. Ann Rheum Dis 2013; 72 (Suppl 2): ii100–ii103.
Monteleone G, Biancone L, Marasco R, Morrone G, Marasco O, Luzza F et al. Interleukin 12 is expressed and actively released by Crohn's disease intestinal lamina propria mononuclear cells. Gastroenterology 1997; 112: 1169–1178.
Stolfi C, Fina D, Caruso R, Caprioli F, Sarra M, Fantini MC et al. Cyclooxygenase-2-dependent and -independent inhibition of proliferation of colon cancer cells by 5-aminosalicylic acid. Biochem Pharmacol 2008; 75: 668–676.
Becker C, Fantini MC, Wirtz S, Nikolaev A, Kiesslich R, Lehr HA et al. In vivo imaging of colitis and colon cancer development in mice using high resolution chromoendoscopy. Gut 2005; 54: 950–954.
Acknowledgements
This work was supported by the ‘Fondazione Umberto di Mario ONLUS’, Rome, and AIRC (MFAG-12108 to CS and IG-13049 to GM).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on the Oncogene website
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
De Simone, V., Franzè, E., Ronchetti, G. et al. Th17-type cytokines, IL-6 and TNF-α synergistically activate STAT3 and NF-kB to promote colorectal cancer cell growth. Oncogene 34, 3493–3503 (2015). https://doi.org/10.1038/onc.2014.286
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/onc.2014.286
This article is cited by
-
Extracellular vesicle encapsulated Homer1a as novel nanotherapeutics against intracerebral hemorrhage in a mouse model
Journal of Neuroinflammation (2024)
-
Long-chain polyunsaturated fatty acids influence colorectal cancer progression via the interactions between the intestinal microflora and the macrophages
Molecular and Cellular Biochemistry (2024)
-
New gold(III) complexes TGS 121, 404, and 702 show anti-tumor activity in colitis-induced colorectal cancer: an in vitro and in vivo study
Pharmacological Reports (2024)
-
Exploring the therapeutic efficacy of crocetin in oncology: an evidence-based review
Naunyn-Schmiedeberg's Archives of Pharmacology (2024)
-
Dietary diosgenin transcriptionally down-regulated intestinal NPC1L1 expression to prevent cholesterol gallstone formation in mice
Journal of Biomedical Science (2023)
