
Key Points
-
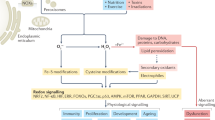
Reactive oxygen species (ROS) homeostatic pathways in microbes operate instant and feedback-regulated adjustments of intracellular ROS concentration. They use redox sensors that 'measure' ROS concentration and proportionally set the expression of ROS scavengers. Because of the high sensitivity of these pathways, sensors can be equated to receptors and ROS signals can be equated to agonists.
-
ROS chemistry dictates reactivity towards selective atomic targets in proteins. The hydroxyl radical HO·is indiscriminate; the superoxide anion O2− is active towards iron–sulphur ([Fe–S]) clusters and hydrogen peroxide (H2O2) targets reactive Cys residues. The subcellular colocalization of ROS and their targets contributes to mammalian ROS signalling specificity.
-
Among prokaryotic [Fe–S]-cluster-based sensors, SoxR is O2−-specific; both FNR (which senses oxygen) and IscR (which senses [Fe–S]-cluster biosynthesis status) also respond to H2O2 and O2−.
-
Among prokaryotic peroxide sensors, OxyR and OhrR use a reactive Cys residue that oxidizes to a disulphide bond, and PerR uses a non-haem iron centre, the two coordinating His residues of which oxidize to 2-oxo-His.
-
The Hsp33 chaperone and RsrA anti-sigma factor use a Cys–zinc redox centre to respond to a combination of H2O2 and heat (Hsp33) and to diamide (RsrA). Their sensitivity is much lower than that of the ROS receptors.
-
Thiol-based peroxiredoxins and GPX-like peroxidases carry H2O2 receptors and have redox transducing functions. Inactivation by overoxidation and reactivation by sulphiredoxin regulate peroxiredoxin antioxidant functions, H2O2 receptor functions and redox transduction functions.
-
Yeast H2O2 homeostatic pathways use redox relays that comprise a thiol peroxidase as the H2O2 receptor that oxidizes a transcription factor: Yap1 in Saccharomyces cerevisiae and Pap1 in Schizosaccharomyces pombe.
-
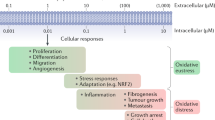
Instead of ROS homeostatic pathways, mammals use global differentiation programmes, such as those regulated by the tumour-suppressor p53, peroxisome proliferator-activated receptor-γ (PPARγ) coactivator-1α, the oncogene c-Myc and class O forkhead box transcription factors (FOXOs). These pathways provide either long-lasting oxidant-protective responses or cell death as clearance mechanisms for oxidatively damaged cells.
-
The KEAP1–NRF2 pathway constitutes the closest fit to a ROS receptor in mammals, and regulates oxidant and xenobiotic stress-protective responses. Sensing involves a Cys–zinc redox centre.
-
The lack of instant ROS homeostatic control in higher eukaryotes might be the inescapable consequence of using ROS as diffusible signals that modulate multiple intracellular signalling pathways.
Abstract
Reactive oxygen species (ROS) have been shown to be toxic but also function as signalling molecules. This biological paradox underlies mechanisms that are important for the integrity and fitness of living organisms and their ageing. The pathways that regulate ROS homeostasis are crucial for mitigating the toxicity of ROS and provide strong evidence about specificity in ROS signalling. By taking advantage of the chemistry of ROS, highly specific mechanisms have evolved that form the basis of oxidant scavenging and ROS signalling systems.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$189.00 per year
only $15.75 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

References
Nathan, C. Specificity of a third kind: reactive oxygen and nitrogen intermediates in cell signaling. J. Clin. Invest. 111, 769–778 (2003). An opinion paper that discusses the physiological and molecular basis of specificity in ROS and RNS signalling.
Halliwell, B. & Gutteridge, J. M. C. Free Radicals in Biology and Medicine (Oxford University Press, Oxford, 1999).
Imlay, J. A. Pathways of oxidative damage. Annu. Rev. Microbiol. 57, 395–418 (2003).
Poole, L. B., Karplus, P. A. & Claiborne, A. Protein sulfenic acids in redox signaling. Annu. Rev. Pharmacol. Toxicol. 44, 325–347 (2004).
Winterbourn, C. C. & Metodiewa, D. Reactivity of biologically important thiol compounds with superoxide and hydrogen peroxide. Free Radic. Biol. Med. 27, 322–328 (1999).
Gilbert, H. F. Molecular and cellular aspects of thiol-disulfide exchange. Adv. Enzymol. Relat. Areas Mol. Biol. 63, 69–172 (1990).
Hofmann, B., Hecht, H. J. & Flohe, L. Peroxiredoxins. Biol. Chem. 383, 347–364 (2002).
Wood, Z. A., Poole, L. B. & Karplus, P. A. Peroxiredoxin evolution and the regulation of hydrogen peroxide signaling. Science 300, 650–653 (2003). Describes the structural basis of the sensitivity of a subgroup of peroxiredoxins to inactivation by overoxidation.
Wood, Z. A., Schroder, E., Robin Harris, J. & Poole, L. B. Structure, mechanism and regulation of peroxiredoxins. Trends Biochem. Sci. 28, 32–40 (2003).
Le Moan, N., Clement, G., Le Maout, S., Tacnet, F. & Toledano, M. B. The Saccharomyces cerevisiae proteome of oxidized protein thiols: contrasted functions for the thioredoxin and glutathione pathways. J. Biol. Chem. 281, 10420–10430 (2006).
D'Autreaux, B. et al. Reversible redox- and zinc-dependent dimerization of the Escherichia coli Fur protein. Biochemistry 46, 1329–1342 (2007).
Ilbert, M., Graf, P. C. & Jakob, U. Zinc center as redox switch — new function for an old motif. Antioxid. Redox Signal 8, 835–846 (2006).
Maret, W. Zinc and sulfur: a critical biological partnership. Biochemistry 43, 3301–3309 (2004).
Conrad, M. et al. The nuclear form of phospholipid hydroperoxide glutathione peroxidase is a protein thiol peroxidase contributing to sperm chromatin stability. Mol. Cell Biol. 25, 7637–7644 (2005).
Maiorino, M. et al. Functional interaction of phospholipid hydroperoxide glutathione peroxidase with sperm mitochondrion-associated cysteine-rich protein discloses the adjacent cysteine motif as a new substrate of the selenoperoxidase. J. Biol. Chem. 280, 38395–38402 (2005).
Hidalgo, E. & Demple, B. An iron-sulfur center essential for transcriptional activation by the redox-sensing SoxR protein. EMBO J. 13, 138–146 (1994).
Mukhopadhyay, P., Zheng, M., Bedzyk, L. A., LaRossa, R. A. & Storz, G. Prominent roles of the NorR and Fur regulators in the Escherichia coli transcriptional response to reactive nitrogen species. Proc. Natl Acad. Sci. USA 101, 745–750 (2004).
Liochev, S. I. & Fridovich, I. Fumarase C, the stable fumarase of Escherichia coli, is controlled by the soxRS regulon. Proc. Natl Acad. Sci. USA 89, 5892–5896 (1992).
Koo, M. S. et al. A reducing system of the superoxide sensor SoxR in Escherichia coli. EMBO J. 22, 2614–2622 (2003).
Crack, J. C., Green, J., Cheesman, M. R., Le Brun, N. E. & Thomson, A. J. Superoxide-mediated amplification of the oxygen-induced switch from [4Fe-4S] to [2Fe-2S] clusters in the transcriptional regulator FNR. Proc. Natl Acad. Sci. USA 104, 2092–2097 (2007).
Khoroshilova, N., Popescu, C., Munck, E., Beinert, H. & Kiley, P. J. Iron-sulfur cluster disassembly in the FNR protein of Escherichia coli by O2: [4Fe-4S] to [2Fe-2S] conversion with loss of biological activity. Proc. Natl Acad. Sci. USA 94, 6087–6092 (1997).
Schwartz, C. J. et al. IscR, an Fe-S cluster-containing transcription factor, represses expression of Escherichia coli genes encoding Fe-S cluster assembly proteins. Proc. Natl Acad. Sci. USA 98, 14895–14900 (2001).
Yeo, W. S., Lee, J. H., Lee, K. C. & Roe, J. H. IscR acts as an activator in response to oxidative stress for the suf operon encoding Fe-S assembly proteins. Mol. Microbiol. 61, 206–218 (2006).
Pantopoulos, K. Iron metabolism and the IRE/IRP regulatory system: an update. Ann. NY Acad. Sci. 1012, 1–13 (2004).
Zheng, M., Aslund, F. & Storz, G. Activation of the OxyR transcription factor by reversible disulfide bond formation. Science 279, 1718–1721 (1998). Identification of the OxyR regulatory disulphide bond by mass spectrometry.
Lee, J. W. & Helmann, J. D. The PerR transcription factor senses H2O2 by metal-catalysed histidine oxidation. Nature 440, 363–367 (2006). Identification of the PerR metal-catalysed mechanism of His oxidation by H 2 O 2.
Fuangthong, M., Atichartpongkul, S., Mongkolsuk, S. & Helmann, J. D. OhrR is a repressor of ohrA, a key organic hydroperoxide resistance determinant in Bacillus subtilis. J. Bacteriol. 183, 4134–4141 (2001).
Mongkolsuk, S. & Helmann, J. D. Regulation of inducible peroxide stress responses. Mol. Microbiol. 45, 9–15 (2002).
Sukchawalit, R., Loprasert, S., Atichartpongkul, S. & Mongkolsuk, S. Complex regulation of the organic hydroperoxide resistance gene (ohr) from Xanthomonas involves OhrR, a novel organic peroxide-inducible negative regulator, and posttranscriptional modifications. J. Bacteriol. 183, 4405–4412 (2001).
Aslund, F., Zheng, M., Beckwith, J. & Storz, G. Regulation of the OxyR transcription factor by hydrogen peroxide and the cellular thiol-disulfide status. Proc. Natl Acad. Sci. USA 96, 6161–6165 (1999).
Belousov, V. V. et al. Genetically encoded fluorescent indicator for intracellular hydrogen peroxide. Nature Methods 3, 281–286 (2006).
Choi, H. et al. Structural basis of the redox switch in the OxyR transcription factor. Cell 105, 103–113 (2001).
Lee, C. et al. Redox regulation of OxyR requires specific disulfide bond formation involving a rapid kinetic reaction path. Nature Struct. Mol. Biol. 11, 1179–1185 (2004).
Toledano, M. B. et al. Redox-dependent shift of OxyR–DNA contacts along an extended DNA-binding site: a mechanism for differential promoter selection. Cell 78, 897–909 (1994).
Kim, S. O. et al. OxyR: a molecular code for redox-related signaling. Cell 109, 383–396 (2002).
Toledano, M. B., Delaunay, A., Monceau, L. & Tacnet, F. Microbial H2O2 sensors as archetypical redox signaling modules. Trends Biochem. Sci. 29, 351–357 (2004).
Aslund, F. & Beckwith, J. Bridge over troubled waters: sensing stress by disulfide bond formation. Cell 96, 751–753 (1999).
Hausladen, A., Privalle, C. T., Keng, T., DeAngelo, J. & Stamler, J. S. Nitrosative stress: activation of the transcription factor OxyR. Cell 86, 719–729 (1996).
Fuangthong, M. & Helmann, J. D. The OhrR repressor senses organic hydroperoxides by reversible formation of a cysteine-sulfenic acid derivative. Proc. Natl Acad. Sci. USA 99, 6690–6695 (2002).
Panmanee, W., Vattanaviboon, P., Poole, L. B. & Mongkolsuk, S. Novel organic hydroperoxide-sensing and responding mechanisms for OhrR, a major bacterial sensor and regulator of organic hydroperoxide stress. J. Bacteriol. 188, 1389–1395 (2006).
Lee, J. W., Soonsanga, S. & Helmann, J. D. A complex thiolate switch regulates the Bacillus subtilis organic peroxide sensor OhrR. Proc. Natl Acad. Sci. USA 104, 8743–8748 (2007). Identification of an OhrR mechanism that involves regulation by protein S -thiolation.
Hong, M., Fuangthong, M., Helmann, J. D. & Brennan, R. G. Structure of an OhrR-OhrA operator complex reveals the DNA binding mechanism of the MarR family. Mol. Cell 20, 131–141 (2005).
Herbig, A. F. & Helmann, J. D. Roles of metal ions and hydrogen peroxide in modulating the interaction of the Bacillus subtilis PerR peroxide regulon repressor with operator DNA. Mol. Microbiol. 41, 849–859 (2001).
Helmann, J. D. et al. The global transcriptional response of Bacillus subtilis to peroxide stress is coordinated by three transcription factors. J. Bacteriol. 185, 243–253 (2003).
D'Autreaux, B. et al. Spectroscopic description of the two nitrosyl-iron complexes responsible for fur inhibition by nitric oxide. J. Am. Chem. Soc. 126, 6005–6016 (2004).
Moore, C. M., Nakano, M. M., Wang, T., Ye, R. W. & Helmann, J. D. Response of Bacillus subtilis to nitric oxide and the nitrosating agent sodium nitroprusside. J. Bacteriol. 186, 4655–4664 (2004).
Chen, P. R. et al. An oxidation-sensing mechanism is used by the global regulator MgrA in Staphylococcus aureus. Nature Chem. Biol. 2, 591–595 (2006).
Jakob, U., Eser, M. & Bardwell, J. C. Redox switch of Hsp33 has a novel zinc-binding motif. J. Biol. Chem. 275, 38302–38310 (2000).
Winter, J., Linke, K., Jatzek, A. & Jakob, U. Severe oxidative stress causes inactivation of DnaK and activation of the redox-regulated chaperone Hsp33. Mol. Cell. 17, 381–392 (2005). Demonstration of the physiological role of Hsp33 as an alternate chaperone system replacing DnaK under severe oxidative stress.
Li, W. et al. The role of zinc in the disulphide stress-regulated anti-sigma factor RsrA from Streptomyces coelicolor. J. Mol. Biol. 333, 461–472 (2003).
Graf, P. C. et al. Activation of the redox-regulated chaperone Hsp33 by domain unfolding. J. Biol. Chem. 279, 20529–20538 (2004).
Graumann, J. et al. Activation of the redox-regulated molecular chaperone Hsp33 — a two-step mechanism. Structure 9, 377–387 (2001).
Kang, J. G. et al. RsrA, an anti-sigma factor regulated by redox change. EMBO J. 18, 4292–4298 (1999).
Zdanowski, K. et al. Assignment of the zinc ligands in RsrA, a redox-sensing ZAS protein from Streptomyces coelicolor. Biochemistry 45, 8294–8300 (2006).
Delaunay, A., Pflieger, D., Barrault, M. B., Vinh, J. & Toledano, M. B. A thiol peroxidase is an H2O2 receptor and redox-transducer in gene activation. Cell 111, 471–481 (2002). Identification of a GPX-like enzyme as the H 2 O 2 receptor that activates Yap1 by oxidation.
Gulshan, K., Rovinsky, S. A., Coleman, S. T. & Moye-Rowley, W. S. Oxidant-specific folding of Yap1p regulates both transcriptional activation and nuclear localization. J. Biol. Chem. 280, 40524–40533 (2005).
Veal, E. A., Ross, S. J., Malakasi, P., Peacock, E. & Morgan, B. A. Ybp1 is required for the hydrogen peroxide-induced oxidation of the Yap1 transcription factor. J. Biol. Chem. 278, 30896–30904 (2003).
Kuge, S., Toda, T., Iizuka, N. & Nomoto, A. Crm1 (XpoI) dependent nuclear export of the budding yeast transcription factor yAP-1 is sensitive to oxidative stress. Genes Cells 3, 521–532 (1998).
Wood, M. J., Storz, G. & Tjandra, N. Structural basis for redox regulation of Yap1 transcription factor localization. Nature 430, 917–921 (2004). Describes the NMR-solved structure of the oxidized form of the Yap1 redox-responsive domain.
Izawa, S. et al. Thioredoxin deficiency causes the constitutive activation of Yap1, an AP-1-like transcription factor in Saccharomyces cerevisiae. J. Biol. Chem. 274, 28459–28465 (1999).
Delaunay, A., Isnard, A. D. & Toledano, M. B. H2O2 sensing through oxidation of the Yap1 transcription factor. EMBO J. 19, 5157–5166 (2000).
Carmel-Harel, O. et al. Role of thioredoxin reductase in the Yap1p-dependent response to oxidative stress in Saccharomyces cerevisiae. Mol. Microbiol. 39, 595–605 (2001).
Yan, C., Lee, L. H. & Davis, L. I. Crm1p mediates regulated nuclear export of a yeast AP-1-like transcription factor. EMBO J. 17, 7416–7429 (1998).
Ikner, A. & Shiozaki, K. Yeast signaling pathways in the oxidative stress response. Mutat. Res. 569, 13–27 (2005).
Quinn, J. et al. Distinct regulatory proteins control the graded transcriptional response to increasing H(2)O(2) levels in fission yeast Schizosaccharomyces pombe. Mol. Biol. Cell. 13, 805–816 (2002).
Vivancos, A. P., Castillo, E. A., Jones, N., Ayte, J. & Hidalgo, E. Activation of the redox sensor Pap1 by hydrogen peroxide requires modulation of the intracellular oxidant concentration. Mol. Microbiol. 52, 1427–1435 (2004).
Bozonet, S. M. et al. Oxidation of a eukaryotic 2-Cys peroxiredoxin is a molecular switch controlling the transcriptional response to increasing levels of hydrogen peroxide. J. Biol. Chem. 280, 23319–23327 (2005).
Vivancos, A. P. et al. A cysteine-sulfinic acid in peroxiredoxin regulates H2O2-sensing by the antioxidant Pap1 pathway. Proc. Natl Acad. Sci. USA 102, 8875–8880 (2005). References 67 and 68 identified the molecular mechanism that restricts the Pap1 response to low levels of H 2 O 2.
Veal, E. A. et al. A 2-Cys peroxiredoxin regulates peroxide-induced oxidation and activation of a stress-activated MAP kinase. Mol. Cell 15, 129–139 (2004).
Mason, J. T., Kim, S. K., Knaff, D. B. & Wood, M. J. Thermodynamic basis for redox regulation of the Yap1 signal transduction pathway. Biochemistry 45, 13409–13417 (2006).
Azevedo, D., Tacnet, F., Delaunay, A., Rodrigues-Pousada, C. & Toledano, M. B. Two redox centers within Yap1 for H2O2 and thiol-reactive chemicals signaling. Free Radic. Biol. Med. 35, 889–900 (2003).
Castillo, E. A. et al. Diethylmaleate activates the transcription factor Pap1 by covalent modification of critical cysteine residues. Mol. Microbiol. 45, 243–254 (2002).
Miao, Y. et al. An Arabidopsis glutathione peroxidase functions as both a redox transducer and a scavenger in abscisic acid and drought stress responses. Plant Cell 18, 2749–2766 (2006).
Rhee, S. G., Chang, T. S., Bae, Y. S., Lee, S. R. & Kang, S. W. Cellular regulation by hydrogen peroxide. J. Am. Soc. Nephrol. 14, S211–S215 (2003).
Conway, J. P. & Kinter, M. Dual role of peroxiredoxin I in macrophage-derived foam cells. J. Biol. Chem. 281, 27991–28001 (2006).
Chang, T. S. et al. Peroxiredoxin III, a mitochondrion-specific peroxidase, regulates apoptotic signaling by mitochondria. J. Biol. Chem. 279, 41975–41984 (2004).
Choi, M. H. et al. Regulation of PDGF signalling and vascular remodelling by peroxiredoxin II. Nature 435, 347–353 (2005). Genetic demonstration of the role of peroxiredoxin-II in modulating signalling downstream of the PDGF receptor.
Chang, T. S. et al. Regulation of peroxiredoxin I activity by Cdc2-mediated phosphorylation. J. Biol. Chem. 277, 25370–25376 (2002).
Cao, C., Leng, Y., Huang, W., Liu, X. & Kufe, D. Glutathione peroxidase 1 is regulated by the c-Abl and Arg tyrosine kinases. J. Biol. Chem. 278, 39609–39614 (2003).
Cao, C., Leng, Y. & Kufe, D. Catalase activity is regulated by c-Abl and Arg in the oxidative stress response. J. Biol. Chem. 278, 29667–29675 (2003).
Sablina, A. A. et al. The antioxidant function of the p53 tumor suppressor. Nature Med. 11, 1306–1313 (2005). Documentation of the antioxidant function of p53.
St-Pierre, J. et al. Suppression of reactive oxygen species and neurodegeneration by the PGC-1 transcriptional coactivators. Cell 127, 397–408 (2006).
Benassi, B. et al. c-Myc phosphorylation is required for cellular response to oxidative stress. Mol. Cell 21, 509–519 (2006).
Burgering, B. M. & Kops, G. J. Cell cycle and death control: long live Forkheads. Trends Biochem. Sci. 27, 352–360 (2002).
Tothova, Z. et al. FoxOs are critical mediators of hematopoietic stem cell resistance to physiologic oxidative stress. Cell 128, 325–339 (2007).
Essers, M. A. et al. FOXO transcription factor activation by oxidative stress mediated by the small GTPase Ral and JNK. EMBO J. 23, 4802–4812 (2004).
Lehtinen, M. K. et al. A conserved MST–FOXO signaling pathway mediates oxidative-stress responses and extends life span. Cell 125, 987–1001 (2006).
van der Horst A. et al. FOXO4 is acetylated upon peroxide stress and deacetylated by the longevity protein hSir2(SIRT1). J. Biol. Chem. 279, 28873–28879 (2004).
Brunet, A. et al. Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase. Science 303, 2011–2015 (2004).
Itoh, K. et al. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev. 13, 76–86 (1999). The identification of KEAP1 as a negative regulator of NRF2.
Cullinan, S. B., Gordan, J. D., Jin, J., Harper, J. W. & Diehl, J. A. The Keap1-BTB protein is an adaptor that bridges Nrf2 to a Cul3-based E3 ligase: oxidative stress sensing by a Cul3-Keap1 ligase. Mol. Cell Biol. 24, 8477–8486 (2004).
Furukawa, M. & Xiong, Y. BTB protein Keap1 targets antioxidant transcription factor Nrf2 for ubiquitination by the Cullin 3–Roc1 ligase. Mol. Cell Biol. 25, 162–171 (2005).
Kobayashi, A. et al. Oxidative stress sensor Keap1 functions as an adaptor for Cul3-based E3 ligase to regulate proteasomal degradation of Nrf2. Mol. Cell Biol. 24, 7130–7139 (2004).
Zhang, D. D., Lo, S. C., Cross, J. V., Templeton, D. J. & Hannink, M. Keap1 is a redox-regulated substrate adaptor protein for a Cul3-dependent ubiquitin ligase complex. Mol. Cell. Biol. 24, 10941–10953 (2004). References 91–94 identified KEAP1 as an adaptor of a CUL3-based ubiquitin ligase.
Dinkova-Kostova, A. T., Holtzclaw, W. D. & Kensler, T. W. The role of Keap1 in cellular protective responses. Chem. Res. Toxicol. 18, 1779–1791 (2005).
Kobayashi, M. & Yamamoto, M. Nrf2-Keap1 regulation of cellular defense mechanisms against electrophiles and reactive oxygen species. Adv. Enzyme Regul. 46, 113–140 (2006).
Dinkova-Kostova, A. T. et al. Direct evidence that sulfhydryl groups of Keap1 are the sensors regulating induction of phase 2 enzymes that protect against carcinogens and oxidants. Proc. Natl Acad. Sci. USA 99, 11908–11913 (2002). The first documentation of the role of Cys residues in the regulation of KEAP1.
Levonen, A. L. et al. Cellular mechanisms of redox cell signalling: role of cysteine modification in controlling antioxidant defences in response to electrophilic lipid oxidation products. Biochem. J. 378, 373–382 (2004).
Zhang, D. D. & Hannink, M. Distinct cysteine residues in Keap1 are required for Keap1-dependent ubiquitination of Nrf2 and for stabilization of Nrf2 by chemopreventive agents and oxidative stress. Mol. Cell. Biol. 23, 8137–8151 (2003).
Wakabayashi, N. et al. Keap1-null mutation leads to postnatal lethality due to constitutive Nrf2 activation. Nature Genet. 35, 238–245 (2003).
Eggler, A. L., Liu, G., Pezzuto, J. M., van Breemen, R. B. & Mesecar, A. D. Modifying specific cysteines of the electrophile-sensing human Keap1 protein is insufficient to disrupt binding to the Nrf2 domain Neh2. Proc. Natl Acad. Sci. USA 102, 10070–10075 (2005).
Gao, L. et al. Novel n-3 fatty acid oxidation products activate Nrf2 by destabilizing the association between Keap1 and Cullin3. J. Biol. Chem. 282, 2529–2537 (2007).
Tong, K. I. et al. Keap1 recruits Neh2 through binding to ETGE and DLG motifs: characterization of the two-site molecular recognition model. Mol. Cell. Biol. 26, 2887–2900 (2006).
Lo, S. C., Li, X., Henzl, M. T., Beamer, L. J. & Hannink, M. Structure of the Keap1:Nrf2 interface provides mechanistic insight into Nrf2 signaling. EMBO J. 25, 3605–3617 (2006).
Nguyen, T., Sherratt, P. J., Nioi, P., Yang, C. S. & Pickett, C. B. Nrf2 controls constitutive and inducible expression of ARE-driven genes through a dynamic pathway involving nucleocytoplasmic shuttling by Keap1. J. Biol. Chem. 280, 32485–32492 (2005).
Velichkova, M. & Hasson, T. Keap1 regulates the oxidation-sensitive shuttling of Nrf2 into and out of the nucleus via a Crm1-dependent nuclear export mechanism. Mol. Cell. Biol. 25, 4501–4513 (2005).
Desaint, S., Luriau, S., Aude, J. C., Rousselet, G. & Toledano, M. B. Mammalian antioxidant defenses are not inducible by H2O2 . J. Biol. Chem. 279, 31157–31163 (2004).
Finkel, T. Oxidant signals and oxidative stress. Curr. Opin. Cell. Biol. 15, 247–254 (2003).
Tonks, N. K. Redox redux: revisiting PTPs and the control of cell signaling. Cell 121, 667–670 (2005).
Li, Q. et al. Nox2 and Rac1 regulate H2O2-dependent recruitment of TRAF6 to endosomal interleukin-1 receptor complexes. Mol. Cell Biol. 26, 140–154 (2006).
Ushio-Fukai, M. Localizing NADPH oxidase-derived ROS. Sci. STKE 349, re8 (2006).
Vilhardt, F. & van Deurs, B. The phagocyte NADPH oxidase depends on cholesterol-enriched membrane microdomains for assembly. EMBO J. 23, 739–748 (2004).
Miller, E. W., Tulyanthan, O., Isacoff, E. Y. & Chang, C. J. Molecular imaging of hydrogen peroxide produced for cell signaling. Nature Chem. Biol. 3, 263–267 (2007).
Bienert, G. P., Schjoerring, J. K. & Jahn, T. P. Membrane transport of hydrogen peroxide. Biochim. Biophys. Acta 1758, 994–1003 (2006).
Salmeen, A. et al. Redox regulation of protein tyrosine phosphatase 1B involves a sulphenyl-amide intermediate. Nature 423, 769–773 (2003).
van Montfort, R. L., Congreve, M., Tisi, D., Carr, R. & Jhoti, H. Oxidation state of the active-site cysteine in protein tyrosine phosphatase 1B. Nature 423, 773–777 (2003).
Denu, J. M. & Tanner, K. G. Specific and reversible inactivation of protein tyrosine phosphatases by hydrogen peroxide: evidence for a sulfenic acid intermediate and implications for redox regulation. Biochemistry 37, 5633–5642 (1998).
Baker, L. M. & Poole, L. B. Catalytic mechanism of thiol peroxidase from Escherichia coli. Sulfenic acid formation and overoxidation of essential Cys61. J. Biol. Chem. 278, 9203–9211 (2003).
Ghezzi, P. Regulation of protein function by glutathionylation. Free Radic. Res. 39, 573–580 (2005).
Seaver, L. C. & Imlay, J. A. Alkyl hydroperoxide reductase is the primary scavenger of endogenous hydrogen peroxide in Escherichia coli. J. Bacteriol. 183, 7173–7181 (2001).
Parsonage, D. et al. Analysis of the link between enzymatic activity and oligomeric state in AhpC, a bacterial peroxiredoxin. Biochemistry 44, 10583–10592 (2005). Demonstration that peroxiredoxin oligomerization changes catalytic Cys reactivity and enzymatic activity.
Woo, H. A. et al. Reversing the inactivation of peroxiredoxins caused by cysteine sulfinic acid formation. Science 300, 653–656 (2003). Documentation of the reversibility of the Cys sulphinic acid form of peroxiredoxin.
Rabilloud, T. et al. Proteomics analysis of cellular response to oxidative stress. Evidence for in vivo overoxidation of peroxiredoxins at their active site. J. Biol. Chem. 277, 19396–19401 (2002).
Budanov, A. V., Sablina, A. A., Feinstein, E., Koonin, E. V. & Chumakov, P. M. Regeneration of peroxiredoxins by p53-regulated sestrins, homologs of bacterial AhpD. Science 304, 596–600 (2004).
Biteau, B., Labarre, J. & Toledano, M. B. ATP-dependent reduction of cysteine-sulphinic acid by S. cerevisiae sulphiredoxin. Nature 425, 980–984 (2003). Identification of the enzyme that catalyses Cys sulphinic-acid reduction in peroxiredoxins.
Yang, K. S. et al. Inactivation of human peroxiredoxin I during catalysis as the result of the oxidation of the catalytic site cysteine to cysteine-sulfinic acid. J. Biol. Chem. 277, 38029–38036 (2002).
Buck, V. et al. Peroxide sensors for the fission yeast stress-activated mitogen-activated protein kinase pathway. Mol. Biol. Cell 12, 407–419 (2001).
Nguyen, A. N., Lee, A., Place, W. & Shiozaki, K. Multistep phosphorelay proteins transmit oxidative stress signals to the fission yeast stress-activated protein kinase. Mol. Biol. Cell 11, 1169–1181 (2000).
Galperin, M. Y., Nikolskaya, A. N. & Koonin, E. V. Novel domains of the prokaryotic two-component signal transduction systems. FEMS Microbiol. Lett. 203, 11–21 (2001).
Gilles-Gonzalez, M. A. & Gonzalez, G. Heme-based sensors: defining characteristics, recent developments, and regulatory hypotheses. J. Inorg. Biochem. 99, 1–22 (2005).
Padmanabhan, B. et al. Structural basis for defects of Keap1 activity provoked by its point mutations in lung cancer. Mol. Cell. 21, 689–700 (2006).
Acknowledgements
We acknowledge members of the Toledano laboratory for discussions, C. Mann for critical reading of the manuscript, and grants from the Agence Nationale pour la Recherche (ANR), Association de Recherche pour le Cancer (ARC) and the CEA ToxNuc programme to M.B.T. We regret that we were unable to cite much interesting work in this area because of space limitations.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary information S1 (table)
Kinetic parameters of selected peroxiredoxins and catalases (PDF 181 kb)
Related links
Related links
DATABASES
Entrez Genome Project
FURTHER INFORMATION
Glossary
- Iron–sulphur clusters
-
Iron–sulphur clusters are metal centres that consist of sulphide (S2−) and iron. The most common structures are the diamond [2Fe–2S] and the cubane [4Fe–4S] clusters. Iron–sulphur clusters are usually coordinated by Cys or His residues at the iron atoms.
- Reactive nitrogen species
-
These are derived from the reaction of nitric oxide (NO) with oxygen or superoxide and include nitrogen trioxide (N2O3), peroxinitrite (ONOO−) and nitrogen dioxide (NO2).
- Peroxidase
-
Peroxide-reducing enzymes that function by heterolytic cleavage of the O–O bond. Peroxidases fall into different classes depending on the nature of their catalytic site or according to the mechanisms that regenerate the active form.
- pKa
-
The equilibrium constant of proton (H+) exchange reactions between acids and bases according to the Brönsted theory. It reflects the strength of an acid to donate its proton as pKa decreases.
- S-nitrosylation
-
A modification to the –SNO form of the thiol moiety of a Cys residue, caused by reaction with peroxinitrite (ONOO−) or nitrogen trioxide (N2O3).
- Alkylation
-
A chemical modification that involves the transfer of a carbon chain to any other atom.
Rights and permissions
About this article
Cite this article
D'Autréaux, B., Toledano, M. ROS as signalling molecules: mechanisms that generate specificity in ROS homeostasis. Nat Rev Mol Cell Biol 8, 813–824 (2007). https://doi.org/10.1038/nrm2256
Issue Date:
DOI: https://doi.org/10.1038/nrm2256
This article is cited by
-
Proteolytic activation of fatty acid synthase signals pan-stress resolution
Nature Metabolism (2024)
-
The effect of psychoactive bacteria, Bifidobacterium longum Rosell®-175 and Lactobacillus rhamnosus JB-1, on brain proteome profiles in mice
Psychopharmacology (2024)
-
Nimbolide Exhibits Potent Anticancer Activity Through ROS-Mediated ER Stress and DNA Damage in Human Non-small Cell Lung Cancer Cells
Applied Biochemistry and Biotechnology (2024)
-
An apoptosis-inducing factor controls programmed cell death and laccase expression during fungal interactions
Applied Microbiology and Biotechnology (2024)
-
Aa-Z2 triggers ROS-induced apoptosis of osteosarcoma by targeting PDK-1
Journal of Translational Medicine (2023)
