
Abstract
To improve the ‘personalized-medicine’ approach to the treatment of depression, we need to identify biomarkers that, assessed before starting treatment, predict future response to antidepressants (‘predictors’), as well as biomarkers that are targeted by antidepressants and change longitudinally during the treatment (‘targets’). In this study, we tested the leukocyte mRNA expression levels of genes belonging to glucocorticoid receptor (GR) function (FKBP-4, FKBP-5, and GR), inflammation (interleukin (IL)-1α, IL-1β, IL-4, IL-6, IL-7, IL-8, IL-10, macrophage inhibiting factor (MIF), and tumor necrosis factor (TNF)-α), and neuroplasticity (brain-derived neurotrophic factor (BDNF), p11 and VGF), in healthy controls (n=34) and depressed patients (n=74), before and after 8 weeks of treatment with escitalopram or nortriptyline, as part of the Genome-based Therapeutic Drugs for Depression study. Non-responders had higher baseline mRNA levels of IL-1β (+33%), MIF (+48%), and TNF-α (+39%). Antidepressants reduced the levels of IL-1β (−6%) and MIF (−24%), and increased the levels of GR (+5%) and p11 (+8%), but these changes were not associated with treatment response. In contrast, successful antidepressant response was associated with a reduction in the levels of IL-6 (−9%) and of FKBP5 (−11%), and with an increase in the levels of BDNF (+48%) and VGF (+20%)—that is, response was associated with changes in genes that did not predict, at the baseline, the response. Our findings indicate a dissociation between ‘predictors’ and ‘targets’ of antidepressant responders. Indeed, while higher levels of proinflammatory cytokines predict lack of future response to antidepressants, changes in inflammation associated with antidepressant response are not reflected by all cytokines at the same time. In contrast, modulation of the GR complex and of neuroplasticity is needed to observe a therapeutic antidepressant effect.
Similar content being viewed by others


INTRODUCTION
Antidepressants are commonly prescribed drugs, but the treatment protocols are dictated by clinical practice and personal preferences rather than by a ‘biomarker-based’ personalized medicine approach (Uher, 2011). These drugs are used in patients with major depression, one of the most common psychiatric disorders and a leading cause of disability worldwide (Gustavsson et al, 2011). However, despite the increasing variety of antidepressants currently available, only a third of patients respond adequately to treatment, and up to half of them relapse within 1 year (Thase, 2006). Unfortunately, we still cannot predict the likelihood of response of an individual patient to a specific drug (Uher, 2011). Therefore, there is a pressing need to identify biomarkers that, assessed before starting treatment, ‘predict’ future response, as well as biomarkers that are ‘targeted’ by antidepressants and change longitudinally during antidepressant treatment. In addition to fostering personalized medicine, establishing ‘predictors’ and ‘target’ biomarkers could lead to the identification of novel pathophysiological pathways relevant to depression, and thus novel mechanisms for designing therapeutic strategies. Based on the current conceptualization of depression, we suggest that hypothesis-driven, blood-based biomarker analysis should focus on the biological systems that have been more consistently described as abnormal in depression: the glucocorticoid receptor (GR) complex, inflammation, and neuroplasticity (Chopra et al, 2011).
One of the most consistent biological findings in depression is a hyperactivity of the hypothalamic–pituitary–adrenal (HPA) axis (Pariante and Lightman, 2008), as shown by a multitude of studies describing high levels of cortisol, the main HPA axis hormone, in the context of reduced function of the GR, the cortisol receptor primarily involved in HPA axis regulation during stress. This reduced GR function, or glucocorticoid resistance, is particularly evident in patients with treatment-resistant depression (Bauer et al, 2003; Juruena et al, 2009), and indeed persistent glucocorticoid resistance during antidepressant treatment is associated with early relapse (Ising et al, 2007; Ribeiro et al, 1993). Moreover, polymorphisms in the GR (or NR3C1) gene, and in the gene for the GR-associated, FKBP-5 co-chaperone protein, have been shown to regulate GR function and to predict antidepressant treatment response (Binder, 2009; Spijker and van Rossum, 2012; Uher et al, 2009). Therefore, the expression levels of GR-related genes are important candidate biomarkers in relation with antidepressant response.
A second biological system potentially involved in antidepressant response is inflammation. Pro-inflammatory cytokines, and in particular interleukin (IL)-1β, IL-6, and tumor necrosis factor (TNF)-α, are increased in depressed patients as compared with controls (Dowlati et al, 2010); in turn, antidepressants have been shown to have anti-inflammatory effects (Hannestad et al, 2011), and anti-inflammatory drugs, such as celecoxib and TNF-α antagonists, have been shown to have antidepressant properties or to improve antidepressant response (Haroon et al, 2012). Interestingly in this context, higher levels of inflammation seem to identify depressed patients who are less likely to respond to antidepressant treatment (Benedetti et al, 2002; Lanquillon et al, 2000; Sluzewska et al, 1997). In addition, polymorphisms in immune genes, such as in IL-1β, IL-11, and TNF-α, have been associated with reduced responsiveness to antidepressant therapy (Uher et al, 2010; Wong et al, 2008; Yu et al, 2003). Consistent with this notion, the elevated levels of IL-1β, IL-6, and TNF-α tend to diminish in parallel with antidepressant response (Janssen et al, 2010; Yoshimura et al, 2009). Of note is also the proposed model that the above-mentioned glucocorticoid resistance results from the direct molecular action of the activated inflammatory pathways on the GR complex, and that, in turn, the glucocorticoid resistance maintains the inflammatory status by reducing the inhibitory control of endogenous glucocorticoids on the immune system (Zunszain et al, 2011). This model suggests a common pathogenic process underlying both the HPA axis and the inflammatory abnormalities in depression, and thus identifies expression levels of inflammatory genes as further important candidate biomarkers in relation with antidepressant response.
Finally, one of the potential mechanisms by which excessive HPA axis activity and inflammatory responses may contribute to the pathogenesis of depression is through inhibition of neurotrophic factors and disturbance of neuroplasticity (Lee and Kim, 2010). The neurotrophin, brain-derived neurotrophic factor (BDNF), is the most studied molecule within the neuroplasticity network, and we and others have shown that BDNF levels are reduced in the serum and in the leukocytes mRNA of depressed patients, and that, in turn, pharmacological and non-pharmacological antidepressant therapies increase BDNF to levels similar to those in healthy controls (Bocchio-Chiavetto et al, 2010; Cattaneo et al, 2010a, 2010b; Pandey et al, 2010). The expression levels of BDNF- and neuroplasticity-related genes are therefore another class of important candidate biomarkers in relation with antidepressant response.
In this study, to capture a comprehensive picture of the biological and clinical interaction between these three biological systems in relationship with antidepressant treatment, we examined the gene expression (blood mRNA via PaxGene tubes) of 15 genes belonging to the GR (three genes), inflammation (nine genes), and neuroplasticity (three genes) pathways. We studied a well-characterized group of depressed patients from the Genome-based Therapeutic Drugs for Depression (GENDEP) study (Uher et al, 2009, 2010), before and after 8 weeks of treatment with one of two pharmacologically different antidepressants: the selective serotonin reuptake inhibitor, escitalopram, and the tricyclic noradrenaline reuptake inhibitor, nortryptline (and in a group of matched controls). We selected all patients (n=74) who were drug-free for at least 2 weeks before enrolling into the trial, and had provided PaxGene tubes at both baseline and follow-up (after 8 weeks of antidepressants). We measured the transcriptional levels of the following genes: for the GR complex, FKBP-4, FKBP-5, and GR; for the inflammatory system, IL-1α, IL-1β, IL-4, IL-6, IL-7, IL-8, IL-10, macrophage inhibiting factor (MIF), and TNF-α; and for neuroplasticity, BDNF, p11, and VGF (non-acronymic). We wished to answer three questions: first, which genes at baseline (ie, before starting antidepressant treatment) differentiate depressed patients vs controls; second, which genes, again at baseline, predict treatment response to subsequent antidepressant treatment (‘predictors’); and third, which genes, assessed prospectively (ie, at baseline and after 8 weeks of antidepressant treatment) change in parallel with treatment response (‘targets’).
MATERIALS AND METHODS
Study Design
The GENDEP project is an open-label part-randomized multicentre pharmacogenetic study with two active pharmacological treatment arms (Keers et al, 2010; Uher et al, 2009, 2010). It was designed to establish clinical and genetic determinants of therapeutic response to two antidepressants with different primary modes of action: nortriptyline and escitalopram. The overall study has been extensively described before (see Supplementary Material for details). In total, 9 psychiatric centers in 8 European countries recruited 811 adult outpatients (296 men and 514 women), aged between 19 and 72 (mean age 42.5, SD=11.8) suffering from unipolar depression of at least moderate severity according to International Classification of Diseases 10/Diagnostic and Statistical Manual of Mental Disorders, fourth edition, and established by the semi-structured SCAN interview.
Severity of depressive symptoms and treatment response was assessed by weekly administration of three established measures of depression severity: the clinician-rated 10-item Montgomery–Asberg Depression Rating Scale (MADRS; Montgomery and Asberg, 1979), the 17-item Hamilton Rating Scale for Depression (HRSD–17; Hamilton, 1967), and the Beck Depression Inventory (BDI; Beck et al, 1961). The average participant in the original sample was in his/her second episode of depression and scored 28.7 (SD=6.7) on the MADRS, 21.8 (SD=5.3) on the 17-item HDRS, and 28.2 (SD=9.7) on the BDI.
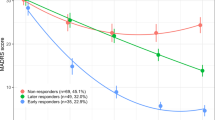
A psychometric analysis has found that the MADRS was the most internally consistent and informative of the three scales (Uher et al, 2008), and therefore in the pharmacogenetic analyses already published (Uher et al, 2010), and in the current gene expression study, we have used the MADRS score as a primary outcome measure of ‘treatment response’. Response to antidepressant medication was quantified as percentage reduction in the MADRS score from baseline to week 12, and responders were identified as patients with a reduction in MADRS>50%.
Subjects
For this study, we selected all patients who had been drug-free for at least 2 weeks before entering into the trial and provided a baseline and a follow-up PaxGene tube (n=74). Their average (SD) age was 38.3±10.9 and, there were 31 males and 43 females. The average participant in our study was in his/her second episode of moderately severe depression and scored, at baseline, 28.7 (SD=4.2) on the MADRS, 20.7 (SD=4.1) on the HRSD–17, and 27.5 (SD=10.2) on the BDI. There were no significant differences between patients treated with escitalopram (n=38) or nortryptiline (n=36), in age (38±12.4 vs 36±9.4, p=0.25), gender (F/M was 20/18 vs 23/13, p=0.2), and in the response rate (responders/non-responders was 26/12 vs 25/11, p=0.6).
Controls were recruited in London (UK), through advertisement in local newspapers, hospitals, and job centers, as well as from existing volunteer databases. Controls were screened using the Psychosis Screening Questionnaire (Bebbington and Nayani, 1995), and were excluded if they met criteria for a present or past psychotic disorder, or if taking any kind of hormonal treatment; their average age was 35.2 (SD=8), and there were 19 males and 15 females. There were no significant differences in age and gender between patients and controls (p=0.14 for age, and p=0.13 for gender distribution).
Gene Expression Analyses
We measured the leukocytes mRNA levels of the above-mentioned candidate genes involved in GR function (FKBP-4, FKBP-5, and GR), inflammatory system (IL1-α, IL-1β, IL-4, IL-6, IL-7, IL-8, IL-10, MIF, and TNF-α), and neuroplasticity (BDNF, p11, and VGF). Each sample was assayed in duplicate, and each target gene was normalized to the expression of three reference genes, glyceraldehyde 3-phosphate dehydrogenase, beta-actin, and beta-2-microglobulin. The expression levels of each target gene were normalized to the geometric mean of all three reference genes, and the Pfaffl method was used to determine relative target gene expression of each gene in patients as compared with controls (see Supplementary Material for details).
Data Analyses
Data were analyzed using the Statistical Package for Social Sciences, version 17.0 (SPSS). Continuous variables are presented as mean±SD or SEM, as indicated. Categorical variables were tested by means of χ2 and Fisher’s tests. Univariate analysis of variance was used for comparing the mean values of the mRNA levels of the genes of interest, at baseline, in patients vs controls and in responders vs non-responders. Changes over time were analyzed using the repeated-measures General Linear Model with time (T0 and T8) and response (yes/no) as factors. The Greenhouse–Geisser correction was applied to degrees of freedom when the sphericity assumption was violated. Parametric correlation analyses using Pearson’s coefficient were used to test the association between genes and the improvement in the depressive symptoms measured as changes in the MADRS score. Linear regression analyses were used to test for predictors of the treatment outcome.
RESULTS
Biomarkers Differences between Patients at Baseline, and Controls
Patients (at baseline) and controls differed in the expression of most of the examined genes (Table 1). Specifically, we found that depressed patients, as compared with controls, had higher FKBP-5 mRNA levels (+27%, F=69.4, p<0.0001) and lower GR mRNA levels (−18%, F=63.2, p<0.0001). Moreover, they had higher mRNA levels of IL-1β, (+48%, F=117.9, p<0.0001), IL-6 (+24%, F=86.3, p<0.0001), MIF (+32%, F=34.8, p<0.0001), and TNF-α (+58%, F=87.7, p<0.0001), and lower levels of IL-4 (−9%, F=5.6, p=0.02). Finally, depressed patients had lower mRNA levels of BDNF (−24%, F=46.5, p<0.0001), p11 (−16%, F=12.1, p=0.001), and VGF (−36%, F=37.3, p<0.0001).
Baseline Differences in Biomarkers between Responders and Non-Responders (‘Predictors’)
As mentioned above, treatment response was defined as a percentage reduction >50% in the MADRS score from baseline to week 12. In this sample, we had 51 responders and 23 non-responders: 26 responders to escitalopram, 12 non-responders to escitalopram, 25 responders to nortryptline, and 11 non-responders to nortryptline. There were no differences in age (38.3±1.6 vs 38.4±2.2 years, F<0.1, p=0.98), gender distribution (F/M=31/20 vs 12/11, χ2=0.5, p=0.3), or baseline MADRS (26.8±0.6 vs 25.0±0.8, F=3.0, p=0.09) between responders and non-responders; moreover, study center also did not influence treatment response (p=0.3).
We compared the baseline mRNA levels of each gene in patients who did not respond to treatment vs patients who did (Table 2). Only three genes were differentially expressed: specifically, non-responders had higher mRNA levels of the three pro-inflammatory cytokines, IL-1β (+33%, F=55.9, p<0.0001), MIF (+48%, F=14.6, p<0.0001), and TNF-α (+39%, F=39.4, p<0.0001). Moreover, for MIF levels we observed a significant drug × response interaction (F=4.4, p=0.04): this was due to the difference between non-responders and responders being larger for non-responders to nortriptyline (+56%, F=73.2, p<0.0001) than for those non-responders to escitalopram (+39%, F=36.9, p<0.0001). There was no drug × response interaction for any of the other genes.
We further examined the relative contributions of the three cytokines in predicting treatment response measured as changes in the MADRS score between week 0 and week 12 (ΔMADRS), both in the overall sample and separately based on the drug used. As expected, the expression levels of IL-1β, MIF, and TNF-α at baseline were all strongly and negatively correlated with the treatment outcome, both in the entire group (IL-1β, r=−0.56; MIF, r=−0.62; and TNF-α, r=−0.44; all p<0.0001), and also separately in the two samples based on the drug used (for escitalopram: IL-1β, r=−0.54, p=0.001; MIF, r=−0.73, p<0.0001; and TNF-α, r=−0.39, p=0.016; and for nortriptyline, IL-1β, r=−0.65 p<0.0001; MIF, r=−0.56 p<0.0001; and TNF-α, r=−0.68, p<0.0001). We then run a linear regression model to identify the relative contributions of the three cytokines to the prediction of response. As shown in Table 3, the best predictive model was obtained when the three cytokines were all included in the model, both in the overall samples (46% of the variance) and separately in the escitalopram-treated group (53% of the variance) and in nortriptyline-treated group (48% of the variance).
Change in Biomarkers and Relationship with Treatment Response (‘Targets’)
To investigate the effect of 8 weeks of antidepressant treatment with escitalopram or nortriptyline on gene expression (and its relationship with treatment response), we compared the change in mRNA levels of each gene between baseline (T0) and week 8 (T8). These data are presented in Table 4.
Three genes were regulated by antidepressant treatment but in responders only, and regardless of the antidepressant used, as shown by significant response × time interactions but no drug × time interactions. Specifically, antidepressant treatment significantly reduced FKBP5 levels only in patients who responded to the treatment (−11%, F=16.4, p<0.0001), whereas no effect was observed in non-responders (−2%, F=0.6, p=0.45; response × time interaction, F=4.4, p=0.04; drug × timeinteraction, F=0.05, p=0.8). Moreover, antidepressant treatment significantly increased VGF expression only in responders (+20%, F=15.4, p<0.0001) but not in non-responders (−3%, F=0.002, p=0.97; response × time interaction, F=4.4, p=0.039; drug × timeinteraction, F=0.03, p=0.8). Finally, antidepressant treatment increased BDNF expression more in the responders (+48%, F=126.4, p<0.0001) than in the non-responders (+21%, F=49.4, p<0.0001; response × time interaction, F=17.8, p<0.0001; drug × time interaction, F=3.6, p=0.062).
IL-6 was regulated by antidepressant treatment but in a drug- and response-specific way, that is, in the presence of both response × time (F=10.0, p=0.002) and drug × time (F=4.4, p=0.039) interactions. Namely, IL-6 levels decreased significantly in responders (−9%, F=20.3, p<0.0001), and this was present for both responders to escitalopram (−12%, F=14.0, p=0.001) and to nortriptyline (−6%, 0.2, F=6.6, p=0.02). In non-responders there was no overall effect (+1%, F=0.4, p=0.5) but, when the two drugs were analyzed separately, IL-6 did not change in the non-responders to escitalopram (−2%, F=0.5, p=0.5) and increased in the non-responders to nortriptyline (+7%, F=5.8, p=0.037).
Finally, four genes were regulated by antidepressant treatment, irrespective of the antidepressant used or of treatment response, as shown by a main effect of time in the absence of any response × time or drug × timeinteractions. Specifically, antidepressant treatment significantly reduced the expression levels of IL-1β (−6%, F=7.9, p=0.006) and MIF (−24%, F=16.4, p<0.0001), and increased GR mRNA levels (+5%, F=7.3, p=0.009) and p11 levels (+8%, F=8.4, p=0.005).
DISCUSSION
To provide evidence supporting a personalized-medicine approach to the treatment of depression, we have assessed the blood mRNA expression of 15 candidate genes across three biological systems implicated in the pathogenesis of depression and in the action of antidepressants: GR complex, inflammation, and neuroplasticity. We have used a well-characterized group of drug-free depressed patients who entered a randomized trial with two different antidepressants, and we have assessed genes that differentiate patients from controls, predict future antidepressants response, or change in association with response. The main finding is a dissociation between genes that predict treatment response (‘predictors’) and genes that change longitudinally in patients who respond (‘targets’). Specifically, among the 15 genes, only higher levels of the three inflammation-related genes, IL-1β, MIF, and TNF-α, predict lack of response to antidepressants, and successful antidepressant response is not associated with a reduction in the levels of these genes. In contrast, successful antidepressant response is associated with a reduction in the levels of the inflammation-related gene, IL-6, and of the GR-associated gene, FKBP-5; and with an increase in the neuroplasticity-associated genes, VGF and BDNF—that is, in genes that are not associated with the baseline prediction of treatment response.
The baseline levels of TNF-α, IL1-β, and MIF, together predict the treatment outcome for both antidepressants, with clinically significant effect sizes of around 50% of the variance (Uher et al, 2012). Moreover, although each single cytokine strongly correlates with the treatment response, the best predictive model is present when we include all three cytokines in the linear regression, suggesting that each cytokine taps also in non-shared molecular mechanisms. For example, IL1-β and TNF-α, but not MIF, activate the indoleamine 2,3-dioxygenase pathway (Capuron et al, 2003; O’Connor et al, 2009), which is responsible for the catabolism of tryptophan to kynurenine, and results in the production of the neurotoxic and depressogenic metabolites, 3-hydroxykynurenine and quinolinic acid (Raison et al, 2010). Indeed, we have recently shown that this pathway is activated by IL-1β also in human neurons (Zunszain et al, 2012). In contrast, MIF, but not IL-1β and TNF-α, is released in response to glucocorticoids, and, when secreted, it renders immune cells less sensitive to the anti-inflammatory effects of glucocorticoids (Kitaichi et al, 2000).
It is also of note that antidepressant treatment in our study reduces the levels of IL1-β and MIF, but this reduction is not associated with treatment response. In contrast, levels of TNF-α, which are elevated in non-responders, are not modified by antidepressant treatment; and levels of IL-6 decrease following antidepressants, but in responders only. Of note, this last finding is remarkably consistent with the previous paper by Yoshimura et al (2009), who also found that IL-6 plasma levels after 8 weeks of antidepressant treatment were reduced in responders only. Although these data confirm previous evidence showing that antidepressants have anti-inflammatory properties (Hiles et al, 2012; Taler et al, 2007), here we also demonstrate that the changes in inflammation associated with antidepressant response are not reflected by all cytokines at the same time. This is perhaps not surprising, because the action of a single cytokine is regulated within a complex network, where multiple pro-inflammatory cytokines maybe involved in the antidepressant response, and a reduction in inflammation may be signaled by an increase in the expression of negative regulators of cytokine action as well as by a reduction in cytokines level. It is also possible that a simple reduction in the levels of some inflammatory biomarkers, such as IL1-β and MIF, is not sufficient for reversing the biological changes underlying the depressive symptoms, because potentially downstream effectors pathways, such as MAPK and nuclear factor kappa B, remain abnormal (Haroon et al, 2012). At the opposite end, persistently high levels of other inflammatory cytokines, such as TNF-α, may be a marker of specific forms of depression that are less responsive to antidepressant treatment, eg, depressed patients with a history of childhood trauma tend to have higher inflammatory biomarkers (Danese et al, 2008) and to be less responsive to pharmacological intervention (Nanni et al, 2012; Nemeroff et al, 2003); in this case, the increased cytokine levels may be an underlying ‘trait feature’ conferring treatment resistance, and may not be modifiable by antidepressants.
The expression of four genes changes only in patients who respond to treatment, but not in those who do not respond. These genes are across the three biological systems investigated: the GR-associated gene, FKBP-5; the pro-inflammatory cytokine, IL-6 (as mentioned above); and the neuroplasticity associated genes, VGF and BDNF. The association between reduction in FKBP-5 and response to antidepressants has never, to our knowledge, been described before; but previous studies have demonstrated that polymorphisms in the gene encoding this co-chaperone are associated with antidepressant response (Binder, 2009; Horstmann et al, 2010; Lekman et al, 2008). Moreover, Menke et al (2012) have recently shown that the pattern of RNA levels of GR-stimulated genes, including FKBP-5, discriminate between patients and controls (Menke et al, 2012). Taken together, our data (see also the changes in GR discussed below) suggest that depression is characterized by the coexistence of higher FKBP-5 and lower GR, leading to GR resistance, and that successful antidepressant treatment requires normalization of GR function. Finally, treatment response was associated with an increase in BDNF and VGF. The evidence for a role of BDNF in depression is abundant (Bocchio-Chiavetto et al, 2010). It is also of note that the recent study from Rojas et al (2011), describing patients who responded to an open-labeled treatment with venlafaxine, also showed an increase in serum BDNF by week 3 of treatment. Also of interest is a large cross-sectional study (Molendjik et al, 2011), again showing that depressed patients had lower levels of serum BDNF, whereas remitted patients (a different group) had normal levels. There are less data on the role of VGF in depression, but previous studies have shown that VGF modulates hippocampal function and behavior through an effect that is BDNF-dependent and that it is involved both in the pathogenesis of depression and in the effects of antidepressants (Cattaneo et al, 2010a, 2010b; Thakker-Varia et al, 2010).
We also find that GR and p11 levels are lower in depressed patients compared with controls, and that their levels increase after antidepressants, but not in relationship with treatment response. Both lower levels of GR and lower levels of p11, which is considered a GR-target gene via two specific glucocorticoid response elements in its promoter (Zhang et al, 2008), have been described before in depressed patients (Anacker et al, 2011; Snyder et al, 2011). To our knowledge, this is the first report describing prospective changes in p11 levels; and only one previous study has assessed GR expression prospectively using radioactive dexamethasone binding in peripheral blood mononuclear cells, and, again, found a reduction of GR density in depressed patients (compared with controls) and an increase after antidepressant treatment (Calfa et al, 2003). Of note, we have consistently shown that antidepressants regulate the function and the expression of the GR (Jurena et al, 2010; Pariante et al, 1997, 2003, 2004, 2012; Pariante and Miller, 2001). Moreover, a recent study in rats has shown that escitalopram increases brain p11 levels concomitantly to a decrease in the p11 promoter methylation (Melas et al, 2011). Indeed, our own research in human neuronal stem cells has recently shown that antidepressants directly increase the function of the GR and, via a GR-dependent mechanism, the expression of p11: an action that is required for the effects of antidepressants on neurogenesis (Anacker et al, 2011).
It is interesting to note that, except for IL-6, we have found that all the examined genes are modulated in the same way by both drugs, even if escitalopram and nortryptiline have different modes of action. We believe this is due to the fact that we have examined genes that belong to pathways that are in common to both drugs (Uher et al, 2009), rather than genes that are modulated differently by the two drugs, that is, belonging to the serotonergic and the noradrenergic pathways, respectively. This is also in line with the results of our previous genetic study in the same GENDEP sample (Uher et al, 2009).
The main limitation of this study is that our finding cannot yet be used to change clinical practice. Notwithstanding the strengths of the randomized, longitudinal design, these findings need to be replicated in an independent sample. Moreover, the predictive biomarkers that we have identified are not specific to one or the other of the antidepressants, and as such could not be used to guide the choice of antidepressants but rather to identify patients that potentially may be helped by early access to adjuvant therapies. Moreover, it is possible that these gene expression changes are not causally involved in the treatment response, and indeed we have not investigated potential biological mechanisms that are changed by antidepressants and may underlie the gene expression changes that we have described, such as changes in immune cells composition or in cortisol levels.
In conclusion, our findings identify for the first time that the baseline levels of MIF, IL1-β, and TNF-α are ‘predictors’ of antidepressant treatment response. Moreover, we show that an enhancement of GR function and an improvement in neuroplasticity are needed to observe a response to antidepressant therapies, suggesting that future antidepressant strategies should specifically target these pathways.
References
Anacker C, Zunszain PA, Carvalho LA, Pariante CM (2011). The glucocorticoid receptor: pivot of depression and of antidepressant treatment? Psychoneuroendocrinology 36: 415–425.
Anacker C, Zunszain PA, Cattaneo A, Carvalho LA, Garabedian MJ, Thuret S et al (2011). Antidepressants increase human hippocampal neurogenesis by activating the glucocorticoid receptor. Mol Psychiatry 16: 738–750.
Bauer ME, Papadopoulos A, Poon L, Perks P, Lightman SL, Checkley S et al (2003). Altered glucocorticoid immunoregulation in treatment resistant depression. Psychoneuroendocrinology 28: 49–65.
Bebbington P, Nayani T (1995). The psychosis screening questionnaire. Int J Methods Psychiatr Res 5: 11–19.
Beck AT, Ward CH, Mendelson M, Mock J, Erbaugh J (1961). An inventory for measuring depression. Arch Gen Psychiatry 4: 561–571.
Benedetti F, Lucca A, Brambilla F, Colombo C, Smeraldi E (2002). Interleukine-6 serum levels correlate with response to antidepressant sleep deprivation and sleep phase advance. Prog Neuropsychopharmacol Biol Psychiatry 26: 1167–1170.
Binder EB (2009). The role of FKBP5, a co-chaperone of the glucocorticoid receptor in the pathogenesis and therapy of affective and anxiety disorders. Psychoneuroendocrinology 1: S186–S195.
Bocchio-Chiavetto L, Bagnardi V, Zanardini R, Molteni R, Nielsen MG, Placentino A et al (2010). Serum and plasma BDNF levels in major depression: a replication study and meta-analyses. World J Biol Psychiatry 11: 763–773.
Calfa G, Kademian S, Ceschin D, Vega G, Rabinovich GA, Volosin M (2003). Characterization and functional significance of glucocorticoid receptors in patients with major depression: modulation by antidepressant treatment. Psychoneuroendocrinology 28: 687–701.
Capuron L, Neurauter G, Musselman DL, Lawson DH, Nemeroff CB, Fuchs D et al (2003). Interferon-alpha-induced changes in tryptophan metabolism. relationship to depression and paroxetine treatment. Biol Psychiatry 54: 906–914.
Cattaneo A, Bocchio-Chiavetto L, Zanardini R, Milanesi E, Placentino A, Gennarelli M (2010a). Reduced peripheral brain-derived neurotrophic factor mRNA levels are normalized by antidepressant treatment. Int J Neuropsychopharmacol 13: 103–108.
Cattaneo A, Sesta A, Calabrese F, Nielsen G, Riva MA, Gennarelli M (2010b). The expression of VGF is reduced in leukocytes of depressed patients and it is restored by effective antidepressant treatment. Neuropsychopharmacology 35: 1423–1428.
Chopra K, Kumar B, Kuhad A (2011). Pathobiological targets of depression. Expert Opin Ther Targets 15: 379–400.
Danese A, Moffitt TE, Pariante CM, Ambler A, Poulton R, Caspi A (2008). Elevated inflammation levels in depressed adults with a history of childhood maltreatment. Arch Gen Psychiatry 65: 409–415.
Dowlati Y, Herrmann N, Swardfager W, Liu H, Sham L, Reim EK et al (2010). A meta-analysis of cytokines in major depression. Biol Psychiatry 67: 446–457.
Gustavsson A, Svensson M, Jacobi F, Allgulander C, Alonso J, Beghi E et al (2011). Cost of disorders of the brain in Europe. Eur Neuropsychopharmacol 21: 718–779.
Hamilton M (1967). Development of a rating scale for primary depressive illness. Brit J Clin Psychol 6: 278–296.
Hannestad J, DellaGioia N, Bloch M (2011). The effect of antidepressant medication treatment on serum levels of inflammatory cytokines: a meta-analysis. Neuropsychopharmacol 36: 2452–2459.
Haroon E, Raison CL, Miller AH (2012). Psychoneuroimmunology meets neuropsychopharmacology: translational implications of the impact of inflammation on behaviour. Neuropsychopharmacology 37: 137–162.
Hiles SA, Baker AL, de Malmanche T, Attia J (2012). Interleukin-6, C-reactive protein and interleukin-10 after antidepressant treatment in people with depression: a meta-analysis. Psychol Med 16: 1–12.
Horstmann S, Lucae S, Menke A, Hennings JM, Ising M, Roeske D et al (2010). Polymorphisms in GRIK4, HTR2A, and FKBP5 show interactive effects in predicting remission to antidepressant treatment. Neuropsychopharmacology 35: 727–740.
Ising M, Horstmann S, Kloiber S, Lucae S, Binder EB, Kern N et al (2007). Combined dexamethasone/corticotropin releasing hormone test predicts treatment response in major depression-a potential biomarker? Biol Psychiatry 62: 47–54.
Janssen DG, Caniato RN, Verster JC, Baune BT (2010). A psychoneuroimmunological review on cytokines involved in antidepressant treatment response. Hum Psychopharmacol 25: 201–215.
Juruena MF, Cleare AJ, Papadopoulos AS, Poon L, Lightman S, Pariante CM (2010). The prednisolone suppression test in depression: dose-response and changes with antidepressant treatment. Psychoneuroendocrinology 35: 1486–1491.
Juruena MF, Pariante CM, Papadopoulos AS, Poon L, Lightman S, Cleare AJ (2009). Prednisolone suppression test in depression: prospective study of the role of HPA axis dysfunction in treatment resistance. Br J Psychiatry 194: 342–349.
Keers R, Uher R, Gupta B, Rietschel M, Schulze TG, Hauser J et al (2010). Stressful life events, cognitive symptoms of depression and response to antidepressants in GENDEP. J Affect Disord 127: 337–342.
Kitaichi N, Kotake S, Mizue Y, Sasamoto Y, Goda C, Iwabuchi K et al (2000). High-dose corticosteroid administration induces increase of serum macrophage migration inhibitory factor in patients with Vogt-Koyanagi-Harada’s disease. Microbiol Immunol 44: 1075–1077.
Lanquillon S, Krieg JC, Bening-Abu-Shach U, Vedder H (2000). Cytokine production and treatment response in major depressive disorder. Neuropsychopharmacol 22: 370–379.
Lee BH, Kim YK (2010). The roles of BDNF in the pathophysiology of major depression and in antidepressant treatment. Psychiatry Investig 7: 231–235.
Lekman M, Laje G, Charney D, Rush AJ, Wilson AF, Sorant AJ et al (2008). The FKBP5-gene in depression and treatment response—an association study in the Sequenced Treatment Alternatives to Relieve Depression (STAR*D) Cohort. Biol Psychiatry 63: 1103–1110.
Melas PA, Rogdaki M, Lennartsson A, Björk K, Qi H, Witasp A et al (2011). Antidepressant treatment is associated with epigenetic alterations in the promoter of P11 in a genetic model of depression. Int J Neuropsychopharmacol 20: 1–11.
Menke A, Arloth J, Pütz B, Weber P, Klengel T, Mehta D et al (2012). Dexamethasone stimulated gene expression in peripheral blood is a sensitive marker for glucocorticoid receptor resistance in depressed patients. Neuropsychopharmacology 37: 1455–1464.
Molendijk ML, Bus BA, Spinhoven P, Penninx BW, Kenis G, Prickaerts J et al (2011). Serum levels of brain-derived neurotrophic factor in major depressive disorder: state-trait issues, clinical features and pharmacological treatment. Mol Psychiatry 16: 1088–1095.
Montgomery SA, Asberg M (1979). A new depression scale designed to be sensitive to change. Br J Psychiatry 134: 382–389.
Nanni V, Uher R, Danese A (2012). Childhood maltreatment predicts unfavorable course of illness and treatment outcome in depression: a meta-analysis. Am J Psychiatry 169: 141–151.
Nemeroff CB, Heim CM, Thase ME, Klein DN, Rush AJ, Schatzberg AF et al (2003). Differential responses to psychotherapy versus pharmacotherapy in patients with chronic forms of major depression and childhood trauma. Proc Natl Acad Sci USA 100: 14293–14296.
O’Connor JC, André C, Wang Y, Lawson MA, Szegedi SS, Lestage J et al (2009). Interferon-gamma and tumor necrosis factor-alpha mediate the upregulation of indoleamine 2,3-dioxygenase and the induction of depressive-like behavior in mice in response to bacillus Calmette-Guerin. J Neurosci 29: 4200–4209.
Pandey GN, Dwivedi Y, Rizavi HS, Ren X, Zhang H, Pavuluri MN (2010). Brain-derived neurotrophic factor gene and protein expression in pediatric and adult depressed subjects. Prog Neuropsychopharmacol Biol Psychiatry 34: 645–651.
Pariante CM, Alhaj HA, Arulnathan VE, Gallagher P, Hanson A, Massey E et al (2012). Central glucocorticoid receptor-mediated effects of the antidepressant, citalopram, in humans: a study using EEG and cognitive testing. Psychoneuroendocrinology 37: 618–628.
Pariante CM, Hye A, Williamson R, Makoff A, Lovestone S, Kerwin RW (2003). The antidepressant clomipramine regulates cortisol intracellular concentrations and glucocorticoid receptor expression in fibroblasts and rat primary neurones. Neuropsychopharmacol 28: 1553–1561.
Pariante CM, Lightman SL (2008). The HPA axis in major depression: classical theories and new developments. Trends Neurosci 31: 464–468.
Pariante CM, Miller AH (2001). Glucocorticoid receptors in major depression: relevance to pathophysiology and treatment. Biol Psychiatry 49: 391–404.
Pariante CM, Pearce BD, Pisell TL, Owens MJ, Miller AH (1997). Steroid-independent translocation of the glucocorticoid receptor by the antidepressant desipramine. Mol Pharmacol 52: 571–581.
Pariante CM, Thomas SA, Lovestone S, Makoff A, Kerwin RW (2004). Do antidepressants regulate how cortisol affects the brain? Psychoneuroendocrinol 29: 423–447.
Raison CL, Borisov AS, Woolwine BJ, Massung B, Vogt G, Miller AH (2010). Interferon-alpha effects on diurnal hypothalamic-pituitary-adrenal axis activity: relationship with proinflammatory cytokines and behavior. Mol Psychiatry 15: 535–547.
Raison CL, Dantzer R, Kelley KW, Lawson MA, Woolwine BJ, Vogt G et al (2010). CSF concentrations of brain tryptophan and kynurenines during immune stimulation with IFN-alpha: relationship to CNS immune responses and depression. Mol Psychiatry 15: 393–403.
Ribeiro SC, Tandon R, Grunhaus L, Greden JF (1993). The DST as a predictor of outcome in depression: a meta-analysis. Am J Psychiatry 150: 1618–1629.
Rojas PS, Fritsch R, Rojas RA, Jara P, Fiedler JL (2011). Serum brain-derived neurotrophic factor and glucocorticoid receptor levels in lymphocytes as markers of antidepressant response in major depressive patients: a pilot study. Psychiatry Res 189: 239–245.
Sluzewska A, Sobieska M, Rybakowski JK (1997). Changes in acute-phase proteins during lithium potentiation of antidepressants in refractory depression. Neuropsychobiology 35: 123–127.
Snyder JS, Soumier A, Brewer M, Pickel J, Cameron HA (2011). Adult hippocampal neurogenesis buffers stress responses and depressive behaviour. Nature 476: 458–461.
Spijker AT, van Rossum EF (2012). Glucocorticoid sensitivity in mood disorders. Neuroendocrinology 95: 179–186.
Taler M, Gil-Ad I, Lomnitski L, Korov I, Baharav E, Bar M et al (2007). Immunomodulatory effect of selective serotonin reuptake inhibitors (SSRIs) on human T lymphocyte function and gene expression. Eur Neuropsychopharmacol 17: 774–780.
Thakker-Varia S, Jean YY, Parikh P, Sizer CF, Jernstedt Ayer J, Parikh A et al (2010). The neuropeptide VGF is reduced in human bipolar postmortem brain and contributes to some of the behavioral and molecular effects of lithium. J Neurosci 30: 9368–9380.
Thase ME (2006). Preventing relapse and recurrence of depression: a brief review of therapeutic options. CNS Spectr 11: 12–21.
Uher R (2011). Genes, environments and individual differences in response to treatment. Harv Rev Psychiatry 19: 109–124.
Uher R, Farmer A, Maier W, Rietschel M, Hauser J, Marusic A et al (2008). Measuring depression: comparison and integration of three scales in the GENDEP study. Psychol Med 38: 289–300.
Uher R, Huezo-Diaz P, Perroud N, Smith R, Rietschel M, Mors O et al (2009). Genetic predictors of response to antidepressants in the GENDEP project. Pharmacogenomics J 9: 225–233.
Uher R, Perroud N, Ng MY, Hauser J, Henigsberg N, Maier W et al (2010). Genome-wide pharmacogenetics of antidepressant response in the GENDEP project. Am J Psychiatry 167: 555–564.
Uher R, Tansey KE, Malki K, Perlis RH (2012). Biomarkers predicting treatment outcome in depression: what is clinically significant? Pharmacogenomics 13: 233–240.
Wong ML, Dong C, Maestre-Mesa J, Licinio J (2008). Polymorphisms in inflammation-related genes are associated with susceptibility to major depression and antidepressant response. Mol Psychiatry 13: 800–812.
Yoshimura R, Hori H, Ikenouchi-Sugita A, Umene-Nakano W, Ueda N, Nakamura J (2009). Higher plasma interleukin-6 (IL-6) level is associated with SSRI- or SNRI-refractory depression. Prog Neuropsychopharmacol Biol Psychiatry 33: 722–726.
Yu YW, Chen TJ, Hong CJ, Chen HM, Tsai SJ (2003). Association study of the interleukin-1 beta (C-511T) genetic polymorphism with major depressive disorder, associated symptomatology, and antidepressant response. Neuropsychopharmacol 28: 1182–1185.
Zhang L, Li H, Su TP, Barker JL, Maric D, Fullerton CS et al (2008). Traumatic Stress Brain Study Group, Ursano R.p11 is up-regulated in the forebrain of stressed rats by glucocorticoid acting via two specific glucocorticoid response elements in the p11 promoter. Neuroscience 153: 1126–1134.
Zunszain PA, Anacker C, Cattaneo A, Carvalho LA, Pariante CM (2011). Glucocorticoids, cytokines and brain abnormalities in depression. Prog Neuropsychopharmacol Biol Psychiatry 35: 722–729.
Zunszain PA, Anacker C, Cattaneo A, Choudhury S, Musaelyan K, Myint AM et al (2012). Interleukin-1β: a new regulator of the kynurenine pathway affecting human hippocampal neurogenesis. Neuropsychopharmacology 37: 939–949.
Acknowledgements
The GENDEP project was supported by a European Commission Framework 6 grant (contract reference: LSHB-CT-2003-503428). This specific project has been supported by a grant from the Commission of European Communities Seventh Framework Programme (Collaborative Project Grant Agreement no. 22963, Mood Inflame) and a Clinician Scientist Fellowship from the Medical Research Council, UK (G108/603), to CMP; by a NARSAD young investigator award to PAZ; by a grant from the Psychiatry Research Trust, UK (McGregor 97) to CMP and AC; by grants from the Italian Ministry of Health (Ricerca Corrente) and Regione Lombardia (ID:17387Sal-13) to MG; and by salary support to CMP, GB, and PZ, and a Studentship to CA, from the National Institute for Health Research Mental Health Biomedical Research Centre at South London and Maudsley NHS Foundation Trust and King’s College London.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
All funding regarding this project is described in the acknowledgment section. Dr Pariante in the last 3 years has received fees as a speaker or as a member of advisory board, as well as research funding, from pharmaceutical companies that commercialize or are developing antidepressants, such as Lilly, Servier, and Janssen. All other authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on the Neuropsychopharmacology website
Supplementary information
Rights and permissions
This work is licensed under the Creative Commons Attribution-NonCommercial-No Derivative Works 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Cattaneo, A., Gennarelli, M., Uher, R. et al. Candidate Genes Expression Profile Associated with Antidepressants Response in the GENDEP Study: Differentiating between Baseline ‘Predictors’ and Longitudinal ‘Targets’. Neuropsychopharmacol 38, 377–385 (2013). https://doi.org/10.1038/npp.2012.191
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2012.191
Keywords
This article is cited by
-
Prediction and Validation of Hub Genes Related to Major Depressive Disorder Based on Co-expression Network Analysis
Journal of Molecular Neuroscience (2024)
-
L’asse dello stress: regolazione neuro-endocrino-immuno-metabolica e ruolo nello sviluppo della sindrome metabolica e della depressione
L'Endocrinologo (2023)
-
Higher immune-related gene expression in major depression is independent of CRP levels: results from the BIODEP study
Translational Psychiatry (2023)
-
Effect of the prosthetic index on stress distribution in Morse taper connection implant system and peri-implant bone: a 3D finite element analysis
BMC Oral Health (2022)
-
Immune targets for therapeutic development in depression: towards precision medicine
Nature Reviews Drug Discovery (2022)
