
Abstract
Chemokine signalling drives monocyte recruitment in atherosclerosis and aortic aneurysms. The mechanisms that lead to retention and accumulation of macrophages in the vascular wall remain unclear. Regulator of G-Protein Signalling-1 (RGS1) deactivates G-protein signalling, reducing the response to sustained chemokine stimulation. Here we show that Rgs1 is upregulated in atherosclerotic plaque and aortic aneurysms. Rgs1 reduces macrophage chemotaxis and desensitizes chemokine receptor signalling. In early atherosclerotic lesions, Rgs1 regulates macrophage accumulation and is required for the formation and rupture of Angiotensin II-induced aortic aneurysms, through effects on leukocyte retention. Collectively, these data reveal a role for Rgs1 in leukocyte trafficking and vascular inflammation and identify Rgs1, and inhibition of chemokine receptor signalling as potential therapeutic targets in vascular disease.
Similar content being viewed by others
Introduction
Chemokine signalling plays a key role in leukocyte trafficking in the pathogenesis of vascular inflammation, the underlying cause of cardiovascular diseases such as atherosclerosis and abdominal aortic aneurysms (AAA). Leukocyte activation and chemotaxis is mediated by chemokines binding to multiple G-protein-coupled receptors (GPCRs). Genetic or pharmacological inhibition of chemokines or chemokine receptors results in reduced atherosclerotic plaque formation1,2,3,4 and inhibits AAA formation5,6. Deletion of either Ccr2 or Ccr5 decreases plaque formation in mouse models of atherosclerosis that is accompanied by reduced macrophages1,2, and deletion of Ccr2 inhibits aortic aneurysm formation6,7. Although the roles for chemokines and chemokine receptors are well defined in the recruitment of leukocytes to the vascular wall in driving disease progression8, it remains less clear how recruited leukocytes accumulate or emigrate in the presence of ongoing chemokine stimulation.
Elucidating these cellular mechanisms would provide strategies targeted at reducing macrophage accumulation or promoting macrophage emigration in the inflamed vasculature. Chemokine receptors couple to Gαi subunits that have an intrinsic GTPase activity, which can be enhanced by RGS proteins, leading to GPCR desensitization. Of the RGS proteins, Regulator of G-Protein Signalling-1 (RGS1) accelerates Gαi GTPase activity and acts to downregulate the response to sustained chemokine activation9,10. Genome-wide association studies have speculated a link between Rgs1 and polymorphisms associated with the risk of several chronic inflammatory diseases such as celiac disease, multiple sclerosis and type I diabetes11,12,13. Other studies have identified a role for RGS1 in the control of lymphocyte homeostasis10,14, but to date no studies have investigated the function of RGS1 in macrophages in vascular inflammation.
Here we report a previously unknown role for RGS1 in monocyte–macrophage trafficking in the development of vascular inflammation. We identified Rgs1 as a novel candidate gene in atherosclerosis and AAA, and tested the hypothesis that RGS1 is a key modulator of chemokine receptor activity, with critical roles in regulating the vascular inflammatory response by affecting macrophage function. We investigated the effects of Rgs1 deletion on macrophage recruitment and retention in the artery wall, and elucidate a new requirement for RGS1 in leukocyte accumulation in atherosclerosis and AAA rupture.
Results
Vascular inflammation increases Rgs1 expression
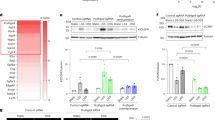
To identify genes that are specifically regulated with atherosclerosis progression, we used a whole mouse genome array to profile the gene expression in the thoracic aortas from ApoE−/− mice, comparing older atherosclerotic ApoE−/− mice with young littermate animals before plaque development (Supplementary Table 1). Among a number of genes already known to play major roles in the development of atherosclerosis such as Ccl2 (MCP-1), Rgs1 was identified as one of the novel candidate genes with higher expression in aortas from older ApoE−/− mice than in aortas from younger ApoE−/− mice. We confirmed that Rgs1 mRNA was upregulated in aortas from atherosclerotic ApoE−/− mice compared with younger ApoE−/− mice (8 weeks, male mice) or wild-type C57BL/6 mice of the same age (16 weeks, male mice) by quantitative reverse transcriptase-PCR (qRT–PCR) (Fig. 1a). The high expression of Rgs1 was associated with the high expression of the macrophage marker Cd68 in individual animals, suggesting that macrophages may be the source of RGS1 in atherosclerotic plaques (Fig. 1b). To test this hypothesis, we quantified Rgs1 expression in different primary cells isolated from ApoE−/− mice and found high Rgs1 mRNA levels in CD68 positive macrophages compared with B cells where Rgs1 is known to have a non-redundant role (Fig. 1c). In contrast, we did not detect Rgs1 mRNA in either vascular smooth muscle cells (VSMCs) or endothelial cells, which are also known to be involved in atherosclerotic plaque progression (Fig. 1c).

(a) Confirmation of Rgs1 mRNA changes in thoracic aortas of ApoE−/− mice and wild-type controls on a high-fat diet (n=5 per group) by qRT–PCR. (b) Rgs1 expression is correlated with the expression of the macrophage marker Cd68 in thoracic aortas of ApoE−/− mice and wild-type controls on a high-fat diet. Each symbol represents an individual mouse (n=5 per group) by qRT–PCR. (c) qRT–PCR analysis of Rgs1 mRNA in primary cells isolated from ApoE−/− mice (n=5–6; BC, B cells; BM, bone marrow cells; EC, endothelial cells; MΦ, macrophages; S, splenocytes). (d) qRT-PCR analysis of Rgs1 expression in human tissue and cells. (Mo; Blood monocytes, plaque macrophages from carotid endarterectomies (n=3), SV; Saphenous vein and IMA; internal mammary artery from CABGs (n=8), OA; omental artery and AAA; abdominal aortic aneurysm from AAA repair patients (n=8–11)). (e) SV and IMA are from CABGs. qRT–PCR analysis of Rgs1 mRNA in Ly6G-7/4hi BM monocytes and peritoneal monocytes isolated from zymosan-induced peritonitis (ZIP) in ApoE−/− mice (n=6–7 per group). (f) qRT–PCR analysis of Rgs1 mRNA in bone marrow-derived macrophages from ApoE−/− mice stimulated with IFN-γ and lipopolysaccharide (M1) and unstimulated (M0) over 24 h presented relative to mRNA in unstimulated cells, set as 1. *P<0.05, **P<0.01 calculated using the Student’s t-test (Data in a are expressed as mean±s.d. and data in c–f as mean±s.e.m.).
To extend these findings in the mouse atherosclerosis model to human disease, we first evaluated Rgs1 expression in macrophages isolated from human carotid artery plaques obtained at endarterectomy. Rgs1 mRNA levels were detectable in macrophages derived from atherosclerotic plaque (Fig. 1d), compared with blood monocytes. Because of the associations between aortic atherosclerosis, vascular inflammation and the pathogenesis of AAA15, we next sought to determine whether Rgs1 is related to human aneurysms. We analysed the tissue samples of AAA from patients undergoing surgical AAA repair, in comparison with non-diseased control samples of omental artery from the same patients, and to internal mammary artery and saphenous vein samples from patients undergoing coronary artery bypass graft surgery (CABG). We also measured Rgs1 in CD14-positive blood monocytes from the same AAA patients. Rgs1 was highly expressed in human AAA tissue compared with non-aneurysmal vascular tissues and blood monocytes (Fig. 1d), suggesting that RGS1 is specifically upregulated in recruited monocytes during vascular inflammation. To test the notion that induction of Rgs1 might occur during monocyte–macrophage activation and recruitment, we compared Rgs1 expression during the differentiation of murine M0 to M1 ‘inflammatory’ macrophages, and between bone marrow monocytes and peritoneal monocytes recruited in a model of sterile peritonitis. Rgs1 was significantly higher in recruited peritoneal monocytes than bone marrow monocytes (Fig. 1e), and upregulated in macrophages during early M1 macrophage differentiation (Fig. 1f). We also confirmed that Rgs1 was abundant in M1-polarized macrophages differentiated from healthy human peripheral blood mononuclear cells (Supplementary Fig. 1).
RGS1 reduces macrophage chemotaxis and desensitization
Since Rgs1 expression in macrophages is high and upregulated with activation, we reasoned that RGS1 would inhibit the migration of macrophages to atherogenic chemokines. We compared the chemotactic responses between ApoE−/− and Rgs1−/−ApoE−/− peritoneal macrophages in vitro. Rgs1−/−ApoE−/− macrophage chemotaxis was significantly increased in response to CCL2, CCL3 and CCL5 (Fig. 2a–c) suggesting a broad specificity for RGS1 to Gαi-coupled chemokine receptors. We also confirmed the role of RGS1 in lymphocyte chemotaxis, by showing increased migration of Rgs1−/−ApoE−/− splenocytes to the homeostatic chemokine CXCL12 (Supplementary Fig. 2) at a similar magnitude to published studies10. Because RGS1 promotes the formation of the inactive G-protein heterotrimer and accelerates the termination of chemokine signalling, we tested the effect of RGS1 on chemokine receptor desensitization in macrophages. We observed that RGS1 reduced the migration to sustained CCL5 stimulation of macrophages. Pretreatment of peritoneal macrophages with increasing doses of CCL5 before chemotaxis to 1 nM CCL5 markedly impaired chemotaxis in ApoE−/− macrophages, whereas Rgs1-deficient macrophages continued to migrate, regardless of previous exposure to chemokine (Fig. 2d). To further address the role of RGS1 in myeloid cell chemotaxis, we used a chemokine-dependent model of sterile inflammation—zymosan-induced peritonitis16 to assess cellular recruitment in vivo. At an early time point after zymosan administration, coinciding with the phase of cellular recruitment to the peritoneum, we observed that the number of monocytes in the peritoneum was significantly increased in Rgs1−/−ApoE−/− mice (Fig. 2e). However, at 16 h after zymosan administration, when cellular recruitment has plateaued and the resolution phase is beginning, we observed a significant decrease in the numbers of monocytes in Rgs1−/−ApoE−/− mice (Fig. 2f) suggesting that an early increase in cell number in Rgs1−/−ApoE−/− mice is then followed by reduced accumulation. To address if there were any alterations in chemokine receptor signalling that may be crucial for trafficking, we assessed CCR5 and CCR2 surface expression on circulating monocytes in mice treated with zymosan. At 4 h, coinciding with the increase in monocytes in the peritoneum of Rgs1−/−ApoE−/− mice, there was an increase in CCR5 on the circulating monocytes compared with monocytes in ApoE−/− mice. In contrast, at 16 h, there was no difference in the cell surface level of CCR5 between Rgs1−/−ApoE−/− and ApoE−/− monocytes (Fig. 2g). However, CCR2 surface expression was not detectable on circulating monocytes from either Rgs1−/−ApoE−/− and ApoE−/− mice after 4 h of zymosan.

Migration of peritoneal macrophages from ApoE−/− and Rgs1−/− ApoE−/− mice through an 8-μm filter towards increasing concentrations of recombinant murine (a) CCL5, (b) CCL3 and (c) CCL2 placed in the lower chamber of a Boyden chamber. (d) Migration of peritoneal macrophages pretreated with 0, 0.1, 1 and 10 nM CCL5 and exposed to 1 nM CCL5. Quantification of migration is presented relative to results of untreated cells, set as 1. RPMI media was used as a negative control. Graphs indicate migration index±s.e.m. for each treatment group (triplicates; n=5–6 per group). In vivo chemotaxis was assessed by i.p. injection of 100 μg zymosan and recruited, peritoneal 7/4hiLy6G− monocytes quantified by flow cytometry at (e) 4 h and (f) 16 h after injection (n=2–4 for saline and n=6–11 for zymosan) (g) The expression of CCR5 on the surface of circulating monocytes after zymosan (n=6–7). Mean fluorescence intensity (MFI) is shown for CCR5 on monocytes at 4 h after zymosan above isotype control (grey). P<0.01 in a,b; P<0.05 in c,d calculated by two-way analysis of variance with significance at individual doses indicated by stars calculated by Bonferroni post-tests. P<0.05 in e–g calculated by Student’s t-test.
RGS1 modulates macrophage trafficking into the aortic wall
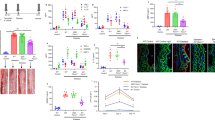
Since macrophage recruitment is a critical step in atherogenesis, and given that Rgs1-deficient macrophages showed an increased migratory response to atherogenic chemokines in vitro, we hypothesized that Rgs1−/−ApoE−/− mice would develop larger atherosclerotic lesions than ApoE−/− mice as a result of enhanced leukocyte recruitment. To test this hypothesis, we quantified atherosclerotic plaque in Rgs1−/−ApoE−/− and ApoE−/− mice at two anatomical sites—the aortic root and the descending aorta. However, in contrast to our hypothesis, the absence of Rgs1 significantly reduced both atherosclerotic plaque formation in the aortic root of ApoE−/− mice (Fig. 3a) and reduced plaque macrophage content, quantified by Galectin-3-positive macrophage immunostaining (Fig. 3b) in 9-week-old animals. Similar results were observed by en face analysis of the descending aorta, where plaques develop later in 16-week-old animals. Rgs1−/−ApoE−/− mice had smaller lesions in comparison with ApoE−/− littermates (Fig. 3c). No differences in lesion size or macrophage content were observed in the aortic root in mice fed a chow diet for 16 weeks or mice on a western-type diet (Supplementary Fig. 3). In addition, no CD3 T-lymphocyte infiltration in the aortic root was observed at any time point (Supplementary Fig. 4). Since Rgs1 has been reported to contribute to T-cell migration17, we also characterized Treg cells and antigen-specific cytokine responses of Th1 and Th17 cells in Rgs1−/−ApoE−/− and ApoE−/− mice after co-stimulation with anti-CD3/CD28, and found no significant difference between groups (Supplementary Figs 5 and 6). No difference in total serum cholesterol levels or circulating monocyte numbers were found between Rgs1−/−ApoE−/− and ApoE−/− mice, indicating that the observed effect was not due to a change in plasma lipids or monocyte numbers, respectively (Supplementary Table 2). In addition, no differences in in vitro foam cell formation were observed between Rgs1−/−ApoE−/− and ApoE−/− macrophages treated with acLDL over 24 h (Supplementary Fig. 7).

(a) Atherosclerotic plaque in the aortic roots of 9-week-old mice on a chow diet. Microscopy of massons trichrome stained aortic root lesions. (b) Galectin-3-positive macrophage content in the aortic roots of 9-week-old mice on a chow diet. Microscopy of Galectin-3-stained aortic root lesions. (c) En face atherosclerotic plaque in the aortas of 16-week-old mice on a chow diet. Microscopy of en face Oil Red O staining of aortic arches of descending aortas. Each symbol represents an individual mouse (n=7–8 per group). Scale bars indicate 0.25 mm for aortic roots and 1 mm for aortas. Arrows indicate atherosclerotic lesions. Values are expressed as mean±s.e.m. ***P<0.001, **P<0.01, *P<0.05 calculated using the Student’s t-test.
To determine the effect of Rgs1 deficiency on the early influx of myeloid cells into the aortic wall during inflammation, we infused Angiotensin II (Ang II) at 0.8 mg kg−1 per day using subcutaneous mini pumps in ApoE−/− mice, a model known to induce acute aortic leukocyte recruitment and increase blood pressure. Leukocyte content in the thoracic aortas of Rgs1−/−ApoE−/− and ApoE−/− mice following Ang II infusion was quantified by enzymatic digestion and flow cytometry (Fig. 4a,b)18,19. In Rgs1-deficient mice, CD45+ leukocytes and CD11b+ myeloid cells in aortas were reduced more than 10-fold compared with ApoE−/− mice after 5 days of Ang II infusion (Fig. 4c–e). These cells were CD14+CCR2+, indicative of recruited monocyte–macrophages. Since Ang II mediates monocyte recruitment to the aorta, and AAA formation is largely driven by CCR2 in contrast to CCR5 (refs 6, 18, 19), we assessed CCR2 expression on these cells. We found that CCR2 surface expression on macrophages in the aortas was significantly decreased following Ang II treatment in ApoE−/− mice (Fig. 4f). However, no reduction in cell surface CCR2 was observed in macrophages from Rgs1−/−ApoE−/− mice, indicating a lack of receptor desensitization in Rgs1−/−ApoE−/− mice. Furthermore, between days 3 and 5 of Ang II infusion, two out of six ApoE−/− mice died from aneurysm rupture, but no deaths occurred in Rgs1−/−ApoE−/− mice, indicating that Rgs1 deficiency confers protection from Ang II-induced aortic aneurysm rupture.

Flow cytometric analysis of aortic leukocytes in ApoE−/− and Rgs1−/− ApoE−/− mice that received (a) saline or (b) Ang II infusion at 0.8 mg kg−1 per day for 5 days. Representative dot plots shown for gated aortic cells of each positive population from ApoE−/− and Rgs1−/− ApoE−/− mice with representative percentages. Labels on both axes are on a log scale. Quantification of the numbers of (c) CD45+ cells (d) CD45+CD14+CD11b+ cells and (e) CD45+CD14+CD11b+CCR2+ cells in saline-treated and Ang II-infused mice. Each symbol represents an individual mouse (n=3–4 for saline and n=4–6 for Ang II). There were two deaths from aneurysm rupture in the ApoE−/− Ang II group. (f) The expression of CCR2 on CD45+CD14+CD11b+ cells in the aorta of saline-treated and Ang II-infused mice (MFI, mean fluorescence intensity). Values are expressed as mean±s.e.m. *P<0.05 and **P<0.01 calculated using the Student’s t-test.
To further investigate the contribution of RGS1 to Ang II-induced aortic aneurysm formation, we performed 14-day Ang II infusions in Rgs1−/−ApoE−/− and ApoE−/− mice, since most aortic ruptures occur within the first 7 days20. ApoE−/− mice were significantly more susceptible to aortic aneurysm rupture in comparison with Rgs1−/−ApoE−/− mice, with 56% survival in ApoE−/− mice versus 94% in the Rgs1−/−ApoE−/−group (Fig. 5a,b). We also noted aneurysms at the study end point in surviving ApoE−/− mice, which were absent in Rgs1−/−ApoE−/− mice (Fig. 5c). No difference in circulating, bone marrow and spleen monocyte numbers were found between Rgs1−/−ApoE−/− and ApoE−/− mice, indicating that the observed effect was not due to a change in monocyte numbers elsewhere (Supplementary Fig. 8). Previous studies have demonstrated that Ang II infusion increases systolic blood pressure in mice7. Therefore, to determine whether Rgs1 deficiency affects Ang II-mediated increases in blood pressure, we measured the systolic blood pressure in both groups. Between days 2 and 10, Ang II treatment increased systolic blood pressure in Rgs1−/−ApoE−/− mice more than in ApoE−/− mice (Fig. 5d), demonstrating that protection from aneurysm formation in Rgs1−/−ApoE−/− mice occurs despite a greater rise in blood pressure and through mechanisms that are independent of Ang II-induced hypertension.

(a) ApoE−/− and Rgs1−/− ApoE−/− mice were infused with Ang II or saline for 14 days. Representative photographs showing features of aneurysms induced by Ang II in surviving mice. The arrows indicate typical aneurysms in ApoE−/− mice. There was no aneurysm formation in the control saline-treated group in both ApoE−/− and Rgs1−/− ApoE−/− mice (n=3–4). (b) Survival curve of ApoE−/− and Rgs1−/− ApoE−/− mice during Ang II (0.8 mg kg−1 per day) infusion. All deaths were due to aortic rupture (c) The incidence of Ang II-induced aortic aneurysms in ApoE−/− mice compared with Rgs1−/− ApoE−/− mice. (d) The systolic blood pressure (BP) of ApoE−/− and Rgs1−/− ApoE−/− mice that were infused with Ang II (0.8 mg kg−1 per day) over 14 days. (e) Survival curve of chimeric ApoE−/− mice during Ang II infusion (3 mg kg−1 per day). All deaths were due to aortic rupture. (f) The incidence of Ang II-induced aortic aneurysms in chimeric ApoE−/− mice. (g) The systolic blood pressure of chimeric ApoE−/− and Rgs1−/− ApoE−/− mice that were infused with Ang II (0.8 mg kg−1 per day) over 14 days. *P<0.05 in b,e calculated using the χ2-test (n=14–19). *P<0.05 in d,g calculated using one-way analysis of variance of area under the curve (n=5–6). Data in d,g are expressed as mean±s.e.m.
Leukocyte Rgs1 deficiency protects against aneurysm rupture
Evidence suggests that chemokines are involved in the modulation of Ang II-accelerated leukocyte recruitment to the vessel wall21. It is well known that macrophages are the predominant leukocyte in Ang II ascending aneurysms and AAAs19,22 and mediate extracellular matrix breakdown that leads to aneurysm formation. We hypothesized that RGS1 may regulate macrophage recruitment and retention via chemokine receptor desensitization and limit macrophage egress and thus augment Ang II-induced AAA development and rupture.
To test this hypothesis, we performed Rgs1−/−ApoE−/− and ApoE−/− bone marrow transplantation into irradiated ApoE−/− mice, to generate bone marrow chimeric animals. Four weeks after transplantation, we tested for Rgs1 and ApoE DNA by PCR in blood from irradiated mice, to confirm complete engraftment. At 6 weeks after engraftment, mice were infused with Ang II for 14 days at 3 mg kg−1 per day. We chose this dose as Ang II at 0.8 mg kg−1 per day was not sufficient to induce aortic aneurysms in irradiated ApoE−/− mice in comparison with non-irradiated mice receiving this dose, despite being sufficient to induce hypertension (Fig. 5g).
We observed that recipient mice transplanted with ApoE−/− bone marrow were more prone to aortic aneurysm rupture in comparison with mice receiving Rgs1−/−ApoE−/− bone marrow (Fig. 5e,f). Over 14 days of Ang II infusion, we found similar survival of ApoE−/− recipient mice with ApoE−/− donor marrow when compared with that in non-chimeric ApoE−/− mice from our original study. In contrast, ApoE−/− recipient mice receiving Rgs1−/−ApoE−/− donor marrow were protected from Ang II-induced AAA formation. Consistent with our earlier findings, there was a significant difference in systolic blood pressure between ApoE−/− mice transplanted with ApoE−/− bone marrow and Rgs1−/−ApoE−/− mice transplanted with ApoE−/− bone marrow (Fig. 5g), revealing a role for vascular wall RGS1 in the control of Ang II-induced blood pressure. Taken together, these results suggest that Rgs1 expression in bone marrow-derived cells, rather than vascular cells, is crucial for aortic aneurysm rupture.
RGS1 promotes leukocyte accumulation in aneurysms
To specifically address the role of Rgs1 in the accumulation or emigration of monocyte-derived cells in the aortic wall during aneurysm development, we used a pulse-chase approach to track bead-labelled monocytes in aortas following Ang II infusion. Inflammatory 7/4hi monocytes were labelled in vivo with fluorescent latex microbeads and administered intravenously (i.v.) at the time of osmotic mini pump implantation23,24. Aortic cell numbers were quantified by flow cytometry at days 3 and 5 after bead injection (Fig. 6a). At 3 days post Ang II infusion, the time of peak monocyte recruitment from the bloodstream, bead-positive leukocyte content in aortas was similar between ApoE−/− and Rgs1−/−ApoE−/− mice, implying that monocyte recruitment was similar between the groups (Fig. 6b). However, by day 5 after the initiation of Ang II infusion, bead-positive CD45+ cells were significantly higher in ApoE−/− aortas compared with Rgs1−/−ApoE−/− mice, suggesting that RGS1 promotes the accumulation of ApoE−/− monocytes in aortic tissue, rather than emigration. Importantly, these findings likely underestimate the magnitude of accumulation of bead-labelled monocytes, since we found several ApoE−/− mice with aneurysms at the time of harvest, which were excluded from the analysis because of the confounding effect of blood cells trapped in aneurysms, to the flow cytometric analysis of aortic cells. Bead-labelled monocytes localized to areas of the subintimal space of the vessel wall and were 7/4 and Ly6C positive (Fig. 6c). In contrast to the difference in bead-labelled cells in the aortic wall, there was no difference in the number of circulating bead-labelled inflammatory monocytes between ApoE−/− and Rgs1−/−ApoE−/− mice at day 3 or 5 (Supplementary Fig. 9). Together these data suggest that RGS1 is a mediator of inflammatory monocyte accumulation in aortic aneurysms.

Flow cytometric analysis of bead-labelled aortic leukocytes in ApoE−/− and Rgs1−/− ApoE−/− mice that received Ang II infusion at 0.8 mg kg−1 per day for 5 days following fluorescent bead labelling of circulating inflammatory monocytes. (a) Representative dot plots shown for gated aortic cells of each positive population from ApoE−/− and Rgs1−/− ApoE−/− mice with representative percentages. Labels on both axes are on a log scale. (b) Quantification of the number of bead-labelled CD45+ cells in aortas of Ang II-infused mice at days 3 and 5. (c) Immunofluorescence microscopy of abdominal aortas from ApoE−/− mice at day 5 after Ang II infusion and bead labelling stained for Ly6C and 7/4 (red), 4′,6-diamidino-2-phenylindole (blue). Arrows indicate the presence of cells containing fluorescent beads (green) on the luminal side (L) of the aorta or within the internal elastic laminar (green autofluorescence). Adv, adventitia. **P<0.01 calculated using the Student’s t-test (n=5–7 per group. Data in b are expressed as mean±s.e.m.).
Discussion
Leukocyte recruitment and accumulation leading to vascular inflammation are critical components of major vascular diseases such as atherosclerosis and AAA. Thus, the signals that regulate these processes are crucial for understanding the underlying causes of inflammatory diseases and to identifying novel therapies. Chemokines and their receptors are rational therapeutic targets in vascular inflammation as indicated by several gene targeting6,25 and inhibition studies4,5. However, functional redundancy within chemokines and chemokine receptors confers limited potential as therapeutic targets. Targeting downstream pathways that modulate chemokine receptor signalling is an alternative strategy but the mechanisms behind this regulation are not fully elucidated.
For the first time, we provide new evidence for a role for RGS1, a downstream mediator of GPCR signalling, in the recruitment and accumulation of leukocytes to the aorta during vascular inflammation in atherosclerosis, aortic aneurysm formation and aneurysm rupture. The major findings of this study are, first, that Rgs1 is upregulated in atherosclerotic vessels and in AAA, is low in circulating monocytes but is greatly upregulated in response to activation and macrophage differentiation (Fig. 7); second, that Rgs1 deficiency increases macrophage chemotaxis and reduces chemokine receptor homologous desensitization; and third, Rgs1 deficiency protects against early atherosclerotic plaque and aortic aneurysm rupture in ApoE−/− mice, due to reduced accumulation of leukocytes in the artery wall. Thus, RGS1 contributes to the persistence of macrophages in the initial stages of atherosclerosis and promotes aortic aneurysm formation and rupture.

The expression of Rgs1 in non-activated circulating monocytes is low and upregulated with monocyte activation by pro-inflammatory stimuli during the recruitment phase. Monocytes differentiate into inflammatory macrophages, whereupon they increase their expression of Rgs1, when they are required to generate an inflammatory response. RGS1 terminates chemokine signalling and thereby reduces the capacity for cell migration, which results in the accumulation of macrophages in the subintimal space. RGS1 might be overactivated in the local environment and perpetuate inflammation in the vessel wall. Thus, the differential expression of Rgs1 in monocytes and macrophages may affect early atherosclerotic lesion and aortic aneurysm progression in vivo. This may also be applicable in other inflammatory diseases where dysregulation of monocyte–macrophage trafficking can influence the result of an inflammatory response.
Our identification of Rgs1 in a gene expression analysis in atherosclerotic vessels from older ApoE−/− mice compared with younger ApoE−/− mice, and in macrophages from human plaques, is in keeping with other studies in human atherosclerotic arteries. Such studies have reported Rgs1 upregulation in advanced calcified aortic valve stenosis26, atherosclerotic coronary arteries27 and unstable carotid artery plaques28. Plaque instability can lead to rupture, the underlying cause of myocardial infarction. Our study now reveals that Rgs1 is specifically upregulated in monocytes and macrophages by inflammatory stimuli, indicating that Rgs1 expression is high in the recruited cells that contribute to plaque formation and/or is further upregulated in the cells within the fatty-streak lesions or aneurysms of ApoE−/− mice, and in human aneurysms. The more complex plaque lesions characteristic of more advanced atherosclerosis are influenced by multiple processes such as cholesterol cleft formation, VSMC proliferation, cellular apoptosis and non-cellular lipid accumulation, rather than the more dominant effect of monocyte recruitment and macrophage accumulation that dominate early lesion formation. Our finding supports the notion that RGS1 expression acts to reduce ongoing chemokine signalling in recruited cells, leading to accumulation at sites of inflammation, such that in Rgs1−/−ApoE−/− mice, inflammatory cell accumulation is significantly reduced.
The functional role of RGS1 has previously been limited to lymphocytes and the control of lymphocyte migration to lymphoid homing chemokines. Several studies have suggested both atheroprotective29,30,31 and proatherogenic32,33,34 roles for the different B-cell and T-cell subsets. Hypercholesterolemic Rag1−/− mice have reduced atherosclerosis at 16 weeks on a chow diet, but no differences on a western-type diet35. The functional roles of lymphocytes in AAA are less clear. Both B cells and T cells have been detected within AAA36 but are thought to have minor roles given that Rag1−/− mice show modest protection from AAA formation37. Rgs1−/− B and T cells show increased migration to CXCL12 (refs 10, 14) and Rgs1−/− B cells still retain this exaggerated response after pre-exposure due to impaired desensitization9,10. In addition, when Rgs1−/− and wild-type T cells are transferred in the inflammatory colitis model in Rag2-deficient mice, wild-type mice were more susceptible to colitis, presumably through RGS1 repressing T-cell egress from the gut14 associating RGS1 in having a key role in leukocyte accumulation. We examined both the T-cell and B-cell phenotype in of Rgs1−/−ApoE−/− mice and found no significant alterations, suggesting RGS1 in lymphocytes in the context of atherosclerosis and AAA has a less significant role to that in myeloid cells in vascular inflammation.
Although the findings of this study implicate RGS1 in the formation of fatty-streak lesions and aortic aneurysm rupture, identification of the downstream signalling pathways by which macrophages accumulate remains to be determined. Monocyte–macrophage trafficking to and from the inflamed vasculature is regulated by a number of different mechanisms, such as adhesion, differentiation, retention, proliferation, apoptosis and egress which are also regulated by chemokines. We observed that RGS1 deletion increased macrophage migration to CCL2 and CCL5. Both of these chemokines are involved in monocyte recruitment to atherosclerotic lesions1,23,25 and their respective receptors are high on inflammatory Ly6Chi monocytes23, which were the cells recruited in our bead-tracking studies in aortic aneurysms, suggesting RGS1 acts as a ‘stop’ signal downstream of these receptors to reduce signalling and migration. A minor role for RGS1 in reducing integrin-dependent adhesion through the N-formyl-methionine-leucine-phenylalanine receptor has been reported in a transfected B-cell line L1.2, although the response was less sensitive to RGS1 action on chemotaxis38. Atherosclerotic plaque regression and pro-resolution pathways are expected to act via inhibiting macrophage accumulation, by promoting macrophage egression or by recruiting patrolling Ly6Clo monocytes in reparative processes. Studies in models of atherosclerosis regression suggest that macrophages exhibit a dendritic cell-like state and emigrate from lesions to lymph nodes in a CCR7-dependent manner39. RGS1 has been reported to regulate CCR7-mediated T-cell chemotaxis to CCL19 (ref. 14), a chemokine receptor pairing not explored in this study, but a process that could bring about resolution of inflammation. Neuronal guidance molecules such as Netrin-1 and Semaphorin 3E inhibit macrophage chemotaxis and promote the persistence of inflammation by retaining macrophages in the plaque40,41. These molecules like RGS1, have immunomodulatory functions in the migration and activation of macrophages, indicating the significance of negative regulators in chronic inflammatory diseases.
Our study identifies therapeutic targeting of RGS1 to reduce local vascular inflammation as a new rational strategy for the treatment of cardiovascular diseases. RGS proteins have previously been identified as drug targets, but further understanding on their regulation is needed42,43. Current strategies have focused on altering RGS protein interactions with Gα protein subunits or the localization or expression of a particular RGS protein in a defined cell44. The structure of RGS1 and its binding sites to Gαi protein is known45 and so it is logical to propose that an agent to prevent RGS1 binding to its Gαi protein in macrophages, in chronic inflammation may have therapeutic potential. In summary, we have demonstrated a new requirement for RGS1 as a key regulator of chemokine receptor signalling and leukocyte trafficking in vascular inflammation. This provides new insights into the mechanisms and importance underlying the recruitment and retention of leukocytes in vascular inflammation and presents new strategies in targeting RGS1 in limiting atherogenesis and AAA formation.
Methods
Mice
The generation of Rgs1−/− mice has been previously described10. Rgs1−/− mice were crossed onto an ApoE−/− background (Charles River, UK) to generate matched litters of Rgs1−/−ApoE−/− and ApoE−/− mice. Mice were housed in individually ventilated cages with 12-h light/dark cycle and controlled temperature (20–22 °C). Standard chow (B & K Universal Ltd, UK) and water were available ad libitum. All animal studies were conducted with ethical approval from the Local Ethical Review Committee and in accordance with the UK Home Office Animals (Scientific Procedures) Act 1986.
Human blood and tissue sampling
Subjects undergoing open AAA repair were prospectively recruited from the Oxford Abdominal Aortic Aneurysm study. Baseline characteristics of each participant were recorded. For each subject, peripheral venous blood was collected after overnight fasting, before commencement of the surgery. During the surgery, a wedge of abdominal omentum containing a segment of omental artery was identified and biopsied en bloc. Isolation of omental artery was performed immediately in the operating theatre. The omental artery segment was cleared of perivascular tissue and snapped frozen. Before incision of the aortic aneurysm, a marker pen was used to denote the cross-section of maximal dilatation according to visual inspection. A longitudinal strip of the aneurysm wall along the incision was then excised. The aneurysm tissue was stripped off perivascular tissue and mural thrombus. The tissue at the maximal dilatation was isolated, divided into smaller segments and snap frozen for subsequent analysis. The study was approved by the Oxford regional ethics committee (Ethics Reference: 13/SC/0250). All subjects gave written informed consent before the study procedure. Human carotid artery plaque macrophages from endarterectomy specimens were isolated following enzymatic digestion of intimal artery segments and ex vivo culture46. Human saphenous vein and internal mammary artery samples from coronary artery bypass graft surgery patients were dissected, excess adventitia removed and the lumen flushed gently using an insulin syringe to remove blood, before snap freezing for RNA47.
Gene expression profiling
Thoracic aortas from either 8- or 16-week-old ApoE−/− mice fed a high-fat diet were homogenized in TRIzol reagent (Sigma-Aldrich, UK) and total RNA isolated using RNeasy kits (Qiagen, UK) and reverse transcribed to complementary DNA (cDNA) using SuperScript II Reverse Transcriptase (Invitrogen, UK). Gene expression was measured using a custom-built whole mouse gene array (12,000 genes) with 70mer probes, analysed using the Lucidea system using the Wellcome Trust Centre for Human Genetics Core Genomics Facility. Each sample was arrayed across six slides with two-colour analysis, with reference samples and included a dye-swap. Data analysis was performed using GeneSpring software. The microarray data have been deposited in the NCBI Gene Expression Omnibus under the accession code GSE65494.
Quantitative real-time RT–PCR
Total RNA was isolated using RNeasy kits (Qiagen) and reverse transcribed to cDNA using SuperScript II Reverse Transcriptase (Invitrogen). Quantitative real-time PCR was performed with 10–50 ng of cDNA on an iCycler IQ real-time detection system (Bio-Rad Laboratories, UK). Gene expression was determined using TaqMan Gene Expression Assays (Applied Biosystems, UK; Supplementary Table 3) relative to the level of the house keeping genes β-actin for mouse, and GAPDH for human using real-time RT–PCR. Relative quantitation of gene expression was performed using the comparative Ct method (ΔΔCt).
Primary cell isolation and culture
Primary cells and whole thoracic aortas were obtained from 8–16-week-old ApoE−/− and Rgs1−/−ApoE−/− mice. Mouse peritoneal macrophages were isolated by peritoneal lavage 4 days after an intraperitoneal (i.p.) injection of 2% BioGel polyacrylamide beads (Bio-Rad Laboratories). Cells were washed in PBS and used for chemotaxis assays or adhered for 2 h in OptiMEM (0.2% bovine serum albumin, BSA) before washing and cell lysis. Mouse splenocytes were isolated by passing spleen pieces through a 70-μm cell strainer. The cell suspension was resuspended in hypotonic lysis buffer (one part Tris-HCl 0.17 M, pH 7.2 to nine parts NH4Cl) to lyse red blood cells and washed before lysis for RNA. Purification of B cells was performed by magnetic cell sorting (Miltenyi Biotec, UK) of splenocytes and confirmed by flow cytometry. B cells (>90% pure) were isolated by positive selection using anti-B220 microbeads (Miltenyi Biotec). Bone marrow cells were isolated by flushing the femur and tibia with PBS and a single-cell suspension prepared by passing through a 70-μm cell strainer before lysis. For bone marrow-derived macrophages, bone marrow cells were isolated and the cell suspension plated into Petri dishes with DMEM-F12 (supplemented with 1% penicillin–streptomycin, 1% L-glutamine, 10% fetal bovine serum and 15% L929-cell-conditioned media). Cells were cultured for 7 days in a humidified atmosphere at 37 °C with 5% CO2 and then harvested and polarized to M1 inflammatory macrophages with lipopolysaccharide (100 ng ml−1; Sigma-Aldrich) and interferon-γ (20 ng ml−1; Peprotech EC) for 24 h, compared with untreated (M0).
Primary endothelial cells were isolated from PBS-perfused lung tissue. Lungs were finely minced and digested in an enzyme solution of DMEM containing 0.18 U ml−1 Liberase Blendzyme 3 (130 μl; Roche, UK) and 0.1 mg ml−1 DNase I (100 μl; Roche) for 1 h at 37 °C with gentle agitation. Purification of endothelial cells was performed by magnetic cell sorting (Miltenyi Biotec) by positive selection using anti-CD31 microbeads (Miltenyi Biotec). Primary VSMCs were isolated by the aortic explant method. Aortas were cut into small segments following endothelial cell denudation and adhered onto 2% gelatin-coated wells and cultured in DMEM complete growth media for 2 weeks. Outgrowing VSMCs from explants of aortic tissue were harvested using Trypsin/EDTA and pelleted for RNA. Mouse peritoneal monocytes were isolated by peritoneal lavage 24 h after i.p. injection of 10 μg Zymosan A (Sigma-Aldrich) and then purified by magnetic cell sorting (Miltenyi Biotec) and confirmed by flow cytometry. Monocytes (>90% pure) were isolated by positive selection using anti-7/4 microbeads (Miltenyi Biotec). Peripheral blood human mononuclear cells were isolated by density-gradient centrifugation (MP Biochemicals, UK). Purification of monocytes was performed by magnetic cell sorting (Miltenyi Biotec) and confirmed by flow cytometry. Monocytes (>90% pure) were isolated by positive selection using anti-CD14 microbeads (Miltenyi Biotec).
Chemotaxis assays
Cells were resuspended in RPMI (25 mM HEPES and 0.1% BSA) to a cell density of 5 × 106 cells ml−1. Eighty μl of the cell suspension was placed on top of 96-well Neuroprobe ChemoTx membranes (5.7 mm diameter, 8 μm pore size; Receptor Technologies, UK) and allowed to migrate towards recombinant murine CCL2, CCL3 or CCL5 (Peprotech EC) or RPMI in lower chambers (320 μl per well) for 4 h at 37 °C, 5% CO2. To assess desensitization, macrophages were prestimulated with 0, 0.1, 1 or 10 nM CCL5 for 10 min followed by washing to remove chemokine and then allowed to migrate towards 1 nM CCL5. Cell migration was quantified from fluorescent microscopic images of cells on the underside of the membranes, with a minimum of three replicate wells per treatment.
Zymosan-induced peritonitis
Mice were injected i.p. with 100 μg of zymosan A (Sigma-Aldrich) diluted in 0.5 ml of PBS or PBS alone. Four or 16 h later, the mice were killed and the peritoneal cavity was lavaged with 5 ml of PBS+2 mM EDTA. Peritoneal exudate cells were stained with antibodies against Ly6G (BD Biosciences) and 7/4 (AbD Serotec, UK) and analysed by flow cytometry to assess monocyte recruitment.
Atherosclerosis analysis
Mice were fed a normal mouse chow (B & K Universal, UK) and harvested at 9 or 16 weeks of age. For the high-fat diet cohort, 8-week-old mice were fed a western-type diet for 8 weeks and harvested at 16 weeks of age. Atherosclerotic lesion size was assessed in paraffin-embedded aortic root sections stained with Masson-Goldner trichrome (Merck, Germany). The average lesion size was calculated from three sections taken at 100-μm intervals starting from the section showing all three aortic cusps. The infiltration of macrophages into aortic lesions was analysed using anti-Galectin-3 (BD Pharmingen, UK) immunostaining (Supplementary Table 4). Aortic lipid deposition was assessed in fixed aortas stained with Oil red O (Sigma-Aldrich) from mice fed a chow diet for 16 weeks. Aortic roots were visualized and imaged (coolSNAP-pro camera, Roper Scientific, Leica DMRBE microscope and the lesion area and Galectin-3-positive areas were quantified from digitized microscopic images using Image-Pro Plus (Media Cybernetics, USA).
Ang II infusion and blood pressure recordings
Eight- to 16-week-old male mice were anaesthetized with isoflurane by inhalation and osmotic mini pumps (Alza Corp, USA) delivering saline or Ang II (0.8 or 3 mg kg−1 per day; Sigma-Aldrich) for 3, 5 or 14 days were implanted subcutaneously. Systolic blood pressure was measured using a non-invasive computerized tail-cuff system in 16-week-old conscious mice following a 1 week training period (Visitech BP2000, Visitech Systems Inc., USA).
Flow cytometry
Descending aortas from the aortic arch to femoral bifurcations were microdissected and digested in an enzyme solution containing 60 U ml−1 DNase I, 60 U ml−1 Hyalronidase, 450 U ml−1 Collagenase I and 125 U ml−1 Collagenase XI (all enzymes from Sigma-Aldrich) at 37 °C (ref. 18). A single-cell suspension was prepared by passing aortic pieces through a strainer for subsequent flow cytometry staining. Isolated aortic cells were antibody stained for the surface markers PE-Cy7-conjugated CD45 (BD Pharmingen), total PerCP-conjugated CD11b (BD Pharmingen), PE-conjugated CD14 (eBioscience, UK), APC-conjugated CCR2 (R&D Systems, UK) with appropriate isotype controls19 (Supplementary Table 5). Absolute cell counts were performed by ratio to a known quantity of calibration beads added to each sample (CaliBrite, BD Pharmingen). Data were acquired using a CyAn Analyser flow cytometer (Beckman Coulter, UK) and then analysed using Summit (Dako, UK) and FlowJo (Tree Star Inc, USA) software.
Bone marrow transplantation
Ten-week-old male mice received a lethal dose of whole-body irradiation (2 × 5 Gy) followed by an i.v. injection of 5 × 106 bone marrow cells from male ApoE−/− and Rgs1−/−ApoE−/− mice. As a control for the efficiency of the irradiation procedure and bone marrow transfer, bone marrow from CD45.1 antigen-expressing mice was transplanted to CD45.2 antigen-expressing mice and blood samples taken 4 weeks after engraftment and then antibody stained for the presence of the donor CD45.1 or recipient CD45.2 allele by flow cytometry. More than 95% blood cells expressed the donor CD45.1 antigen confirming successful reconstitution. Bone marrow transplantation resulted in a significant reduction in the incidence of aneurysms at 0.8 mg kg−1 per day Ang II. To achieve a similar degree of aneurysm to that of non-bone marrow-transplanted mice, we increased the dose of Ang II to 3 mg kg−1 per day after dosing studies.
Inflammatory blood monocyte tracking
Inflammatory 7/4hi monocytes were labelled in vivo by i.v. injection of 1 μm Fluoresbrite green fluorescent plain microspheres (Polysciences Inc., Germany) diluted 1:4 in saline, 18 h post i.v. injection of 200 μl clodronate liposomes (Clodronate Liposomes Org, Amsterdam)23,24. Labelling efficiency was confirmed by flow cytometry. Mice were then implanted with Ang II osmotic mini pumps (0.8 mg kg−1 per day). Flow cytometry was performed after 3 days (during the recruitment and clearance phase from the blood) and 5 days (during influx into aortic tissue) for quantification of bead-labelled cells in the aorta and blood.
Immunofluorescence staining
Abdominal aortas from bead-injected mice were sectioned (10 μm) and blocked for 2 h in 5% goat serum, 1% BSA, 0.2% gelatin, 0.2% Triton-X-100 in PBS. Thereafter, the sections were incubated for 2 h with primary antibodies against Ly6C (BD Pharmingen) and 7/4 (AbD Serotec) followed by incubation with the secondary antibody Alexa Fluor 568 (Invitrogen; Supplementary Table 4). Sections were washed and mounted with mowial mounting medium with 4′,6-diamidino-2-phenylindole. Images were obtained with a Zeiss 510 MetaHead confocal fluorescence microscope.
Statistical analysis
Between-group comparisons of normally distributed measurements were assessed by Student’s t-test. One-way analysis of variance was used to compare more than two data groups and Dunnett’s post-test was used to compare each group with a control (untreated) group. Two-way analysis of variance was used to compare multiple data groups affected by two independent variables, with a Bonferroni correction to compare groups with each other. Differences were considered statistically significant at P<0.05.
Additional information
Accession codes: The microarray data has been deposited in the NCBI Gene Expression Omnibus under the accession code GSE65494.
How to cite this article: Patel, J. et al. RGS1 regulates myeloid cell accumulation in atherosclerosis and aortic aneurysm rupture through altered chemokine signalling. Nat. Commun. 6:6614 doi: 10.1038/ncomms7614 (2015).
Accession codes
References
Boring, L., Gosling, J., Cleary, M. & Charo, I. F. Decreased lesion formation in CCR2−/− mice reveals a role for chemokines in the initiation of atherosclerosis. Nature 394, 894–897 (1998) .
Braunersreuther, V. et al. Ccr5 but not Ccr1 deficiency reduces development of diet-induced atherosclerosis in mice. Arterioscler. Thromb. Vasc. Biol. 27, 373–379 (2007) .
Veillard, N. R. et al. Antagonism of RANTES receptors reduces atherosclerotic plaque formation in mice. Circ. Res. 94, 253–261 (2003) .
Bursill, C. A., Choudhury, R. P., Ali, Z., Greaves, D. R. & Channon, K. M. Broad-spectrum CC-chemokine blockade by gene transfer inhibits macrophage recruitment and atherosclerotic plaque formation in apolipoprotein E-knockout mice. Circulation 110, 2460–2466 (2004) .
Iida, Y. et al. Peptide inhibitor of CXCL4-CCL5 heterodimer formation, MKEY, inhibits experimental aortic aneurysm initiation and progression. Arterioscler. Thromb. Vasc. Biol. 33, 718–726 (2013) .
MacTaggart, J. N., Xiong, W., Knispel, R. & Baxter, B. T. Deletion of CCR2 but not CCR5 or CXCR3 inhibits aortic aneurysm formation. Surgery 142, 284–288 (2007) .
Daugherty, A. et al. Angiotensin II infusion promotes ascending aortic aneurysms: attenuation by CCR2 deficiency in apoE−/− mice. Clin. Sci. (Lond) 118, 681–689 (2010) .
Zernecke, A. & Weber, C. Chemokines in atherosclerosis: proceedings resumed. Arterioscler. Thromb. Vasc. Biol. 34, 742–750 (2014) .
Han, S. B. et al. Rgs1 and Gnai2 regulate the entrance of B lymphocytes into lymph nodes and B cell motility within lymph node follicles. Immunity 22, 343–354 (2005) .
Moratz, C., Hayman, J. R., Gu, H. & Kehrl, J. H. Abnormal B-cell responses to chemokines, disturbed plasma cell localization, and distorted immune tissue architecture in Rgs1−/− mice. Mol. Cell. Biol. 24, 5767–5775 (2004) .
Smyth, D. J. et al. Shared and distinct genetic variants in type 1 diabetes and celiac disease. N. Engl. J. Med. 359, 2767–2777 (2008) .
Johnson, B. A. et al. Multiple sclerosis susceptibility alleles in African Americans. Genes Immun. 11, 343–350 (2010) .
Hunt, K. A. et al. Newly identified genetic risk variants for celiac disease related to the immune response. Nat. Genet. 40, 395–402 (2008) .
Gibbons, D. L. et al. Cutting Edge: regulator of G protein signaling-1 selectively regulates gut T cell trafficking and colitic potential. J. Immunol. 187, 2067–2071 (2011) .
Golledge, J. & Norman, P. E. Atherosclerosis and abdominal aortic aneurysm: cause, response, or common risk factors? Arterioscler. Thromb. Vasc. Biol. 30, 1075–1077 (2010) .
Cash, J. L., White, G. E. & Greaves, D. R. Chapter 17. Zymosan-induced peritonitis as a simple experimental system for the study of inflammation. Methods Enzymol. 461, 379–396 (2009) .
Agenes, F., Bosco, N., Mascarell, L., Fritah, S. & Ceredig, R. Differential expression of regulator of G-protein signalling transcripts and in vivo migration of CD4+ naive and regulatory T cells. Immunology 115, 179–188 (2005) .
Galkina, E. et al. Lymphocyte recruitment into the aortic wall before and during development of atherosclerosis is partially L-selectin dependent. J. Exp. Med. 203, 1273–1282 (2006) .
Tieu, B. C. et al. An adventitial IL-6/MCP1 amplification loop accelerates macrophage-mediated vascular inflammation leading to aortic dissection in mice. J. Clin. Invest. 119, 3637–3651 (2009) .
Cao, R. Y., Amand, T., Ford, M. D., Piomelli, U. & Funk, C. D. The murine angiotensin II-induced abdominal aortic aneurysm model: rupture risk and inflammatory progression patterns. Front. Pharmacol. 1, 9 (2010) .
Mateo, T. et al. Angiotensin II-induced mononuclear leukocyte interactions with arteriolar and venular endothelium are mediated by the release of different CC chemokines. J. Immunol. 176, 5577–5586 (2006) .
Rateri, D. L. et al. Endothelial cell-specific deficiency of Ang II type 1a receptors attenuates Ang II-induced ascending aortic aneurysms in LDL receptor−/− mice. Circ. Res. 108, 574–581 (2011) .
Tacke, F. et al. Monocyte subsets differentially employ CCR2, CCR5, and CX3CR1 to accumulate within atherosclerotic plaques. J. Clin. Invest. 117, 185–194 (2007) .
Potteaux, S. et al. Suppressed monocyte recruitment drives macrophage removal from atherosclerotic plaques of Apoe−/− mice during disease regression. J. Clin. Invest. 121, 2025–2036 (2011) .
Combadiere, C. et al. Combined inhibition of CCL2, CX3CR1, and CCR5 abrogates Ly6C(hi) and Ly6C(lo) monocytosis and almost abolishes atherosclerosis in hypercholesterolemic mice. Circulation 117, 1649–1657 (2008) .
Anger, T. et al. Statins stimulate RGS-regulated ERK 1/2 activation in human calcified and stenotic aortic valves. Exp. Mol. Pathol. 85, 101–111 (2008) .
Archacki, S. R. et al. Identification of new genes differentially expressed in coronary artery disease by expression profiling. Physiol. Genomics 15, 65–74 (2003) .
Papaspyridonos, M. et al. Novel candidate genes in unstable areas of human atherosclerotic plaques. Arterioscler. Thromb. Vasc. Biol. 26, 1837–1844 (2006) .
Major, A. S., Fazio, S. & Linton, M. F. B-lymphocyte deficiency increases atherosclerosis in LDL receptor-null mice. Arterioscler. Thromb. Vasc. Biol. 22, 1892–1898 (2002) .
Caligiuri, G., Nicoletti, A., Poirier, B. & Hansson, G. K. Protective immunity against atherosclerosis carried by B cells of hypercholesterolemic mice. J. Clin. Invest. 109, 745–753 (2002) .
Ait-Oufella, H. et al. Natural regulatory T cells control the development of atherosclerosis in mice. Nat. Med. 12, 178–180 (2006) .
Ait-Oufella, H. et al. B cell depletion reduces the development of atherosclerosis in mice. J. Exp. Med. 207, 1579–1587 (2010) .
Kyaw, T. et al. Conventional B2 B cell depletion ameliorates whereas its adoptive transfer aggravates atherosclerosis. J. Immunol. 185, 4410–4419 (2010) .
Lutgens, E., Lievens, D., Beckers, L., Donners, M. & Daemen, M. CD40 and its ligand in atherosclerosis. Trends Cardiovasc. Med. 17, 118–123 (2007) .
Dansky, H. M., Charlton, S. A., Harper, M. M. & Smith, J. D. T and B lymphocytes play a minor role in atherosclerotic plaque formation in the apolipoprotein E-deficient mouse. Proc. Natl Acad. Sci. USA 94, 4642–4646 (1997) .
Ocana, E., Bohorquez, J. C., Perez-Requena, J., Brieva, J. A. & Rodriguez, C. Characterisation of T and B lymphocytes infiltrating abdominal aortic aneurysms. Atherosclerosis 170, 39–48 (2003) .
Wang, Y. et al. TGF-beta activity protects against inflammatory aortic aneurysm progression and complications in angiotensin II-infused mice. J. Clin. Invest. 120, 422–432 (2010) .
Bowman, E. P. et al. Regulation of chemotactic and proadhesive responses to chemoattractant receptors by RGS (regulator of G-protein signaling) family members. J. Biol. Chem. 273, 28040–28048 (1998) .
Trogan, E. et al. Gene expression changes in foam cells and the role of chemokine receptor CCR7 during atherosclerosis regression in ApoE-deficient mice. Proc. Natl Acad. Sci. USA 103, 3781–3786 (2006) .
Wanschel, A. et al. Neuroimmune guidance cue Semaphorin 3E is expressed in atherosclerotic plaques and regulates macrophage retention. Arterioscler. Thromb. Vasc. Biol. 33, 886–893 (2013) .
van Gils, J. M. et al. The neuroimmune guidance cue netrin-1 promotes atherosclerosis by inhibiting the emigration of macrophages from plaques. Nat. Immunol. 13, 136–143 (2012) .
Neubig, R. R. & Siderovski, D. P. Regulators of G-protein signalling as new central nervous system drug targets. Nat. Rev. Drug Discov. 1, 187–197 (2002) .
Riddle, E. L., Schwartzman, R. A., Bond, M. & Insel, P. A. Multi-tasking RGS proteins in the heart: the next therapeutic target? Circ. Res. 96, 401–411 (2005) .
Roman, D. L. & Traynor, J. R. Regulators of G protein signaling (RGS) proteins as drug targets: modulating G-protein-coupled receptor (GPCR) signal transduction. J. Med. Chem. 54, 7433–7440 (2011) .
Soundararajan, M. et al. Structural diversity in the RGS domain and its interaction with heterotrimeric G protein alpha-subunits. Proc. Natl Acad. Sci. USA 105, 6457–6462 (2008) .
Monaco, C. et al. Canonical pathway of nuclear factor kappa B activation selectively regulates proinflammatory and prothrombotic responses in human atherosclerosis. Proc. Natl Acad. Sci. USA 101, 5634–5639 (2004) .
Antoniades, C. et al. Rapid, direct effects of statin treatment on arterial redox state and nitric oxide bioavailability in human atherosclerosis via tetrahydrobiopterin-mediated endothelial nitric oxide synthase coupling. Circulation 124, 335–345 (2011) .
Acknowledgements
We thank A. Handa and the Vascular Surgery Unit (John Radcliffe Hospital, Oxford University Hospital NHS Trust) for human blood and tissue sampling; C. Monaco and C. Antoniades (Oxford University) for human cDNA samples. We thank H. Lockstone (Oxford University) for microarray interpretation. This work was supported by the British Heart Foundation (RG/10/15/28578 and RG/12/5/29576) and the Wellcome Trust (091504/Z/10/Z and 090532/Z/09/Z), and the National Institute for Health Research (NIHR) Oxford Biomedical Research Centre.
Author information
Authors and Affiliations
Contributions
J.P., E.M., G.D., A.B.H., J.d.B., R.L., A.J.I., D.R.-K. and E.S. performed the experiments, analysed and interpreted the data. D.R.G. provided discussions; J.P, E.M, J.d.B. and K.M.C. designed the experiments and J.P. and K.M.C. wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Information
Supplementary Figures 1-9 and Supplementary Tables 1-5 (PDF 399 kb)
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Patel, J., McNeill, E., Douglas, G. et al. RGS1 regulates myeloid cell accumulation in atherosclerosis and aortic aneurysm rupture through altered chemokine signalling. Nat Commun 6, 6614 (2015). https://doi.org/10.1038/ncomms7614
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/ncomms7614
This article is cited by
-
RGS2 and female common diseases: a guard of women’s health
Journal of Translational Medicine (2023)
-
Regulator of G protein signaling protein 6 alleviates acute lung injury by inhibiting inflammation and promoting cell self-renewal in mice
Cellular & Molecular Biology Letters (2023)
-
“Mind the GAP”: RGS1 hinders antitumor lymphocytes
Nature Immunology (2021)
-
Targeting regulator of G protein signaling 1 in tumor-specific T cells enhances their trafficking to breast cancer
Nature Immunology (2021)
-
Monocytes and macrophages in abdominal aortic aneurysm
Nature Reviews Cardiology (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
