
Abstract
Triple-negative breast cancer (TNBC) has poor prognostic outcome compared with other types of breast cancer. The molecular and cellular mechanisms underlying TNBC pathology are not fully understood. Here, we report that the transcription factor BCL11A is overexpressed in TNBC including basal-like breast cancer (BLBC) and that its genomic locus is amplified in up to 38% of BLBC tumours. Exogenous BCL11A overexpression promotes tumour formation, whereas its knockdown in TNBC cell lines suppresses their tumourigenic potential in xenograft models. In the DMBA-induced tumour model, Bcl11a deletion substantially decreases tumour formation, even in p53-null cells and inactivation of Bcl11a in established tumours causes their regression. At the cellular level, Bcl11a deletion causes a reduction in the number of mammary epithelial stem and progenitor cells. Thus, BCL11A has an important role in TNBC and normal mammary epithelial cells. This study highlights the importance of further investigation of BCL11A in TNBC-targeted therapies.
Similar content being viewed by others
Introduction
One of the major challenges in treating breast cancer is the heterogeneous nature of the disease1. TNBC accounts for around 15% of all breast cancer cases and in the absence of effective targeted therapies, TNBC patients tend to have a poor prognosis2,3,4. At the molecular level, several distinct subtypes of breast cancer have been identified based on the gene expression profiling3,5,6. The most commonly used classification describes six subtypes: luminal A, luminal B, Her2, claudin low, basal-like breast cancer (BLBC) and normal3,6. More recently, analysis of large numbers of tumour samples as part of the METABRIC study identified 10 pathologically distinct subtypes known as integrative cluster (IC) 1–10 (ref. 5). The majority of TNBC cases (80%) have a BLBC7 or IC10 (ref. 5) gene expression signatures. In addition, cancer sequencing studies have identified mutations of p53, PTEN and BRCA1 in TNBC2,4,8,9. However, driver oncogenic genomic aberrations in TNBC have not been comprehensively identified.
The developmental hierarchies of the mammary epithelium and hematopoietic lineages share many similarities10 in that stem cells progressively give rise to lineage-restricted progenitors, which ultimately differentiate and generate all functional cells. A number of key hematopoiesis transcription factors have important roles in mouse mammary gland development and are human breast cancer genes11,12,13,14,15. For example, the key regulator of T-helper-2 cell development, GATA3, is critical in luminal mammary cell development12,13 and is a luminal breast cancer marker gene16. In this study we interrogated cancer genomics data focusing on a subset of important hematopoiesis factors and identified BCL11A as a novel TNBC oncogene.
Results
BCL11A is highly expressed in triple-negative breast cancer
In an attempt to identify potential TNBC oncogenes, we selected a list of genes known to have important roles in hematopoiesis and investigated their expression across the major molecular subtypes of breast cancer3. We first re-analysed a publically available microarray data set6 and found that out of the examined genes, BCL11A was differentially and highly expressed in BLBC (Supplementary Fig. 1a). This is in sharp contrast to GATA3, which is highly expressed only in luminal subtypes (Supplementary Fig. 1a) and is a known prognostic marker for these tumours 16.
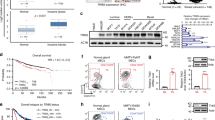
We then investigated the expression of BCL11A in other patient data sets including METABRIC5 and TCGA8, which between them have curated gene expression, copy number (CN) variation and clinical data from close to 3,000 patients5. Pathologically, we found that high BCL11A expression significantly correlated with TNBC pathology (Fig. 1a). At the molecular level, high BCL11A expression was also found to significantly correlate with the BLBC subtype in the METABRIC, TCGA and six other microarray data sets (Fig. 1b and Supplementary Fig. 1b). Quantitative reverse transcription PCR (qRT–PCR) analysis of BCL11A expression on a randomly selected subset of METABRIC tumours (all subtypes, n=230) validated the above expression data (Supplementary Fig. 2a). In addition, we also found that high BCL11A expression in METABRIC samples correlated with the recently described IC10 cluster of tumours (Fig. 1c), thus further supporting the concordance between the BLBC and IC10 classifications. Consistent with TNBC cases, high BCL11A expression was significantly correlated with a high histological grade (Supplementary Fig. 2b).

(a) Significant correlation between BCL11A expression and the TNBC type of breast cancer in both METABRIC (n=2,000) and TCGA (n=1,100) data sets—* indicates t-test P value<0.005. (b) BCL11A expression across the six molecular subtypes of breast cancer (‘Normal’ refers to the PAM50 subtype) in both METABRIC and TCGA data sets—* indicates t-test P value<0.005. (c) The METABRIC samples distributed according to the ICs 1–10, showing the correlation between BCL11A expression and IC10—* indicates t-test P value<0.005.
Furthermore, high BCL11A expression in BLBC cases was further validated by immunohistochemistry (IHC) on a subset of the METABRIC tumours (all subtypes, n=368. BLBC, n=24). Strong BCL11A immunostaining was predominantly found in BLBC (Fig. 2a). Out of 24 BLBC samples examined from this subset, 16 scored positive for BCL11A (Fig. 2a; details in Methods). In addition, samples stained positively in IHC also had higher RNA levels compared with those scored as negative (Supplementary Fig. 2c).

(a) Images and scoring of BCL11A IHC staining on a subset of tumours from the METABRIC study (see Methods for scoring)—* indicated χ2-test P<0.0001. (b–d) Bar chart depicting the percentage of samples that harbour BCL11A CN gains in each subtype in both METABRIC and TCGA data sets—* indicates χ2-test P<0.0001. (e) Scatter plots showing the methylation status of the BCL11A locus in all tumours or basal only from the TCGA data set. The colour-coded legend indicated the BCL11A CN status for each tumour. (f–g) Kaplan–Meir plots showing the survival rate comparison between patients who have normal or high levels of BCL11A expression, or between patients with CN gains or without (neutral).
One mechanism for the induction of high BCL11A expression in BLBC cases could be CN aberrations. From ~2,000 breast cancer cases in METABRIC5, CN gains at the BCL11A genomic locus were identified in 62 patients (Supplementary Fig. 3a), which also correlates with high BCL11A expression (Supplementary Fig. 3b). Importantly, out of these 62 patients with CN gains, 39 were classified as BLBC, which account for 18.6% (39/210) of the total BLBC cases in METABRIC (Fig. 2b). Examination of the TCGA data set revealed that 38% (31/81) of BLBC samples have BCL11A CN gains, which is again significantly correlated with higher gene expression (Fig. 2c and Supplementary Fig. 3c). A similar result was also found when the METABRIC data was analysed using the integrative clustering, with 15.6% of IC10 samples having BCL11A CN gains (Fig. 2d).
Further analysis of the TGCA data set revealed that in BLBCs, the BCL11A locus is almost exclusively hypomethylated and this is correlated with high expression levels (Fig. 2e). There was also no correlation between BCL11A CNs and the methylation status. This result suggests that epigenetic changes at the BCL11A locus could be another mechanism that contributes to its high expression in BLBC. Given the strong correlation with TNBC, patients with either high expression or CN gains of BCL11A had poor survival rates compared with the rest of the cohort (Fig. 2f, g). A similar trend was also observed in four other patient data sets17,18,19,20 (Supplementary Fig. 3 f-i). In particular, patients with CN gains of BCL11A had a higher rate of relapse and metastasis and a lower rate of survival (Supplementary Fig. 3d-e). The utility of BCL11A expression/CN as a biomarker in the clinic thus warrants further investigation. Indeed, the future release of patient outcome for the complete TCGA cohort will aid in clarifying this finding.
High levels of BCL11A promote tumour development
Although BCL11A is involved in rare B-cell lymphomas and is able to transform fibroblast cells in vitro21,22, the cellular and molecular mechanisms of BCL11A-mediated tumourigenesis remains unclear. To address this, we first tested whether BCL11A overexpression could promote the colony formation or tumour development in mammary epithelial cells. We overexpressed BCL11A in immortalized non-tumourigenic mouse EpH4 (ref. 23) or human HMLE24,25 cells (Supplementary Fig. 4a) and performed Matrigel and suspension mammosphere assays. Forced BCL11A expression in both EpH4 and HMLE (EpH4-11A and HMLE-11A) cells resulted in double the number of spheres compared with their respective control cells (Fig. 3a-b). Furthermore, mouse EpH4-11A cells injected orthotopically in cleared mammary fat pads of immune-compromized NOD/SCID/IL2rγ−/− (NSG) mice26 formed larger and palpable tumours compared with control cells (n=6) (Fig. 3c and Supplementary Fig. 4b). Similarly, three out of four mice injected with HMLE-11A cells developed tumours within 8 weeks of injection (Fig. 3d and Supplementary Fig. 4c) suggesting that elevated levels of BCL11A promote tumour development. Moreover, gene expression analysis of these three tumours along with the 2,000 tumours from the METABRIC study clustered them with the BLBC subgroup (Fig. 3e).

(a) Comparison of colony numbers in matrigel from EpH4-11A cells or from the control cells. Data are presented as mean±s.d. (n=3). Image on the right are depicting EpH4-control and EpH4-11A mammospheres grown in Matrigel (scale bar, 100 μm). P value indicates student’s t-test. (b) Comparison of the number of floating mammospheres formed from human HMLE-11A cells or the control cells. Data are presented as mean±s.d. (n=3). Images on the right are of floating mammospheres formed by HMLE-control and HMLE-11A-expressing cell (scale bar, 200 μm). P value indicates student’s t-test. (c) Graph depicting the size difference between tumours at 6 weeks after injection of EpH4-control and EpH4-11A cells orthotopicaly into contralateral mammary fat pads. Data are presented as mean±s.d. (n=6). P value indicates student’s t-test. (d) Kaplan–Meier survival curve depicting the percentage of tumour-free mice injected with either HMLE-control or HMLE-11A cells (n=4). (e) Unsupervized clustering of the HMLE-11A tumours in the mouse with human tumours from the METABRIC study based on the PAM50 (ref. 3) gene expression. Nearest centroid correlation score is plotted against the various subtypes for all three tumours.
Knockdown of BCL11A reduces tumourigenicity of TNBC cells
Analysis of BCL11A expression in a panel of breast cancer cell lines revealed that BCL11A is highly expressed in TNBC lines but is undetectable in any of the luminal cell lines tested (Supplementary Fig. 5a). Next, we assessed if disrupting BCL11A expression could affect the clonogenic and oncogenic potential of the TNBC cell lines. To inactivate BCL11A in these cells, we performed shRNA knockdown experiments (Supplementary Fig. 5b) in the TNBC cell lines 4T1 (mouse), MDA231, SUM159 and HMLER (human). Knockdown of BCL11A had no significant impact on cell viability, cell cycle kinetics or cell death (Fig. 4a-c and Supplementary Fig. 5b,d). However, BCL11A knockdown significantly reduced the clonogenic capacity of all four cell lines (Fig. 4d-f and Supplementary Fig. 5c). To assess tumourigeneic potential, BCL11A knockdown cells were injected subcutaneously into NSG recipients. Robust tumours developed from the control 4T1, MDA231, SUM159 and HMLER cells within 25 days. In contrast, the BCL11A knockdown cells produced tumours of significantly reduced sizes (Fig. 4g-i and Supplementary Fig. 5c). Furthermore, primary and secondary limiting dilution transplantations of MDA231 control or shRNA1 cells revealed a reduction in the number of tumour-initiating cells during the secondary transplants from 1/123 to 1/667 (Supplementary Fig. 5e).

(a) Western blot showing BCL11A knockdown efficiency in 4T1, SUM159 and MDA231 cells transfected with control (scramble), shRNA1 or shRNA2 vectors. (b) MTS cell viability assay (see Methods) shows that BCL11A knockdown does not affect cell viability in culture. (c) EdU cell cycle analysis showing that Bcl11a knockdown does not affect cell cycle kinetics in all tested cell lines. Data are presented as mean±s.d. (n=3). (d–f) Comparison of colony numbers from control, shRNA1 and shRNA2 in 4T1, SUM159 and MDA231 cells. Data are presented as mean±s.d. (n=3). P value indicates student’s t-test. (g–i) Graph depicting the reduction in tumour size observed when shRNA1 or shRNA 2 transfected 4T1, SUM159 or MDA231 cells are injected subcutaneously into mice compared with control. Image under graph shows the actual tumours measured (scale bar, 5 mm). Unpaired T-test was performed on d–i and * indicates P<0.05 and ** indicates P<0.005.
Bcl11a is required for the development of DMBA tumours
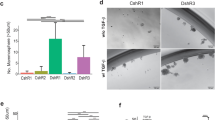
To examine the role of BCL11A in mammary tumour development in vivo, we generated Bcl11a conditional knockout (cko) mice (referred to as flox/flox; Supplementary Fig. 6a), as germline deletion of Bcl11a causes neonatal lethality27 and crossed them to the inducible Rosa26-CreERT2. As a tumour model, we used the potent carcinogen DMBA (7,12-dimethylbenz(a)anthracene) in combination with medroxyprogesterone acetate (MPA) to promote TNBC-like tumours in the mouse28,29. To minimize the effects of Bcl11a deletion on non-mammary tissues, we transplanted mammary tissue from 8- to 12-week-old control (wild type) or flox/flox virgin female mice into contralateral cleared fat pads of female NSG mice followed by DMBA mutagenesis as illustrated in Supplementary Fig. 6b. By week 15, after the last dose of DMBA was administered, palpable tumours were visible in the mammary glands engrafted with the control mammary cells, but not with the flox/flox cells (Fig. 5a). By week 22 post DMBA treatment, all control cell engraftments (8/8) developed tumours compared with only one from flox/flox mammary cells (1/8) (Fig. 5b). qRT–PCR analysis of this tumour revealed expression of Bcl11a probably owing to incomplete Cre-loxP recombination (Supplementary Fig. 6c, sample T1). Also, qRT–PCR and IHC results revealed that tumours upregulated Bcl11a expression in response to DMBA-induced carcinogenesis (Supplementary Fig. 6c-d). These data thus reveal a requirement for Bcl11a in DMBA-induced mammary tumourigenesis.

(a) Mouse images and fat pads whole-mount fat pads of either WT, flox/flox, p53flox/flox or Dflox/flox mammary cells. (scale bar, 500 μm) (b) Quantification of the tumours with DMBA-mediated tumour development from the four depicted groups of engrafted cells over a period of 26 weeks. A log-rank (Mantel–Cox) test was used to compare the two groups and calculate the P value. (c) Tumour size quantification of DMBA-mediated tumours in WT, +/flox and flox/flox mice. Mice were checked regularly for tumour development and once detected, Cre activation was induced using three injections of tamoxifen (first day of injection is indicated by black arrow). Tumours size was then monitored for up to 20 days or until they reach a critical size.
To investigate Bcl11a oncogenic activity in the DMBA model further, we performed the DMBA mutagenesis experiment using Trp53flox/flox30 (p53 single cko) or Bcl11aflox/flox/p53flox/flox (cko alleles for both p53 and Bcl11a or Dflox/flox) mammary tissues. In the recipients transplanted with Trp53flox/flox cells, palpable tumours were detectable as early as 4 weeks after the last injection of DMBA, and most tumours were detectable by week 10 (Fig. 5b; n=16). However, deletion of Bcl11a together with p53 in Dflox/flox mice severely delayed tumour development with only 4 out of 16 mice developeing tumours by week 17 (Fig. 5b). This result indicates that BCL11A is a potent oncogene and is required in concert with p53 for tumour development.
Bcl11a is required for the maintenance of DMBA tumours
Although Bcl11a is important for DMBA-induced mammary tumour formation, it is more clinically relevant if it has functions in mammary tumour progression and maintenance. We thus performed the DMBA mutagenesis on WT, flox/+ and flox/flox mammary epithelial cells before the induction of Bcl11a deletion. Only when mammary tumours were detected and measured, the mice were then injected with tamoxifen to induce Bcl11a deletion. As shown in Fig. 5c, deletion of Bcl11a caused a significant reduction in tumour size as soon as 5 days post deletion. On contrary, tumours from the control heterozygous donor cells continued to grow post tamoxifen injection (Fig. 5c). The requirement of Bcl11a in the established mouse mammary tumours is consistent with the decreased tumourigensis of BCL11A knockdown breast cancer cells and underscores its candidature for therapeutic development.
Bcl11a is required for mammary stem and progenitor cells
To understand the biological function of Bcl11a in healthy mammary epithelial cells, we generated a Bcl11a-lacZ knock-in mouse to determine the temporal and spatial expression of Bcl11a in the mammary gland (Supplementary Fig. 7a). X-gal staining of the reporter embryos revealed that Bcl11a was expressed in the mammary placodes from 12.5dpc (Fig. 6a). At puberty, Bcl11a was expressed in the cap cells of the terminal end buds, a region thought to harbour stem cells31 (Fig. 6b). During adult mammary gland development, Bcl11a exhibited a dynamic expression pattern with a marked increase at early gestation and a gradual decline towards lactation and involution (Supplementary Fig. 7b). qRT–PCR analysis of RNA samples from several mammary epithelial compartments32,33 detected higher levels of Bcl11a expression in the luminal progenitors (CD49b+/CD24hi), the basal cells (CD49Fhi/CD24+) and the mammary stem cell (MaSC) (CD49Fhi/CD24med)-enriched population (Fig. 6c).

(a,b) X-gal staining of a 12.5dpc embryo and whole mount of the mammary gland from a 5-week-old Bcl11a-lacZ/+ female virgin mouse. The dashed circles highlight the mammary placodes and the terminal end buds. LN: lymph node. (scale bar, 500 μm) (c) qRT–PCR for Bcl11a in different mammary epithelial cell compartments that were fluorescence-activated cell sorting-purified using antibodies for CD24, CD49f and CD49b. Data are presented as mean±s.d. (n=3) and t-test was performed and the P values are displayed on the plot. (d) Depletion of the MaSCs-enriched population (CD24medCD49fhi, dashed lines) in the Bcl11a-deficient mammary gland detected by flow cytometric analysis. (e) Limiting dilution transplant (fat pad) assay showing severely compromized engraftment of Bcl11a-deficient MaSCs. Stem cell frequency calculation is described in the Methods.
We next induced Bcl11a deletion and analysed the mammary epithelial fluorescence-activated cell sorting profile 3 weeks post deletion. The basal mammary epithelial cells from the flox/flox mice appeared to be depleted, and in particular the MaSC fraction (Fig. 6d). In addition, Bcl11a deletion caused a significant decrease in the number of luminal colony-forming cells (CFCs) (Supplementary Fig. 7c). To functionally demonstrate loss of MaSC activities upon Bcl11a deletion and to determine that the defects are cell-autonomous, we transplanted control and flox/flox cells at limiting dilution into cleared fat pads of NSG mice (see Methods). We found approximately sixfold reduction in stem cell frequency from 1/483 to 1/2859, in the Bcl11a-deficient mammary gland (Fig. 6e). Reduction of MaSCs and progenitors in the Bcl11a-deficient mammary gland was also reflected in the altered expression of the MaSC gene expression signature34 (Supplementary Table 1) (Supplementary Fig. 7e).
Discussion
We have demonstrated here that the transcription regulator BCL11A is a novel breast cancer gene. By investigating cancer genomics data from ~3,000 patients (METABRIC and TCGA), BCL11A was significantly expressed at higher levels in TNBC and particularly in BLBC/IC10 tumours both at RNA and protein levels. Experimentally, we have shown that disrupting BCL11A expression in TNBC cell lines and in the mouse significantly reduced tumour development and maintenance. At the cellular level, Bcl11a is expressed and required in both MaSCs and luminal progenitor cells in the mammary gland. Lineage tracing experiments in the future will determine if Bcl11a is expressed in the recently identified lineage-restricted luminal and basal progenitor cells35 or in the bipotent MaSCs36. Importantly, given the recent implication of luminal progenitors as the ‘cell of origin’ of BLBC37,38, it will be important to ascertain if Bcl11a upregulation in luminal progenitor cells is one of the earliest steps in TNBC development.
In addition, it will be important to identify how BCL11A is transcriptionally regulated and what are its downstream targets in TNBC. In erythrocytes, KLF1 has been shown to affect BCL11A expression39, while in non-small cell lung cancer MIR30A has been suggested to regulate BCL11A expression40. We found no correlation between KLF1 or MIR30A and BCL11A expression in the TCGA data set (Supplementary Fig. 8), suggesting that BCL11A regulation could be context dependent. In terms of downstream targets, in leukaemia, it has been shown that BCL11A abrogates p21 transcription possibly via direct regulation of SIRT1 (refs 41, 42). Previous work from our lab also showed that in B cells, BCL11A induces MDM2 expression, which is a negative regulator of p53 (ref. 43). However, the TCGA data does not indicate a strong correlation between BCL11A and SIRT1 or MDM2 expression at least in the tumour context (Supplementary Fig. 8). Therefore, identifying the putative BCL11A regulators and its downstream targets in the breast epithelial cells should clarify its molecular and cellular roles in TNBC.
In conclusion, through cancer genomics, in vitro assays, experimental xenograft models and mouse genetics, we have demonstrated in this study that BCL11A is a new breast cancer gene and a critical regulator in normal mammary epithelial development. These results warrant further investigation of BCL11A as a potential candidate for TNBC-targeted therapy.
Methods
Mouse strains and breeding
All experimental animal work was performed in accordance to the Animals (Scientific Procedures) Act 1986, UK and approved by the Ethics Committee at the Sanger Institute. Bcl11a bacterial artificial chromosomes (BACs) were identified from the 129/SvJ mouse BAC library (Sanger Institute) and used to generate the Bcl11a-lacZ- and Bcl11a cko-targeting vectors. For Bcl11a-lacZ reporter, targeting construct (Supplementary Fig. 7a) was generated based on the recently published strategy44. For the Bcl11a cko mouse, targeting construct (Supplementary Fig. 6a) was generated based on the original recombineering strategy45. Gene targeting in embryonic stem (ES) cells and chimera production were performed according to the standard procedures. The Bcl11a cko line was then crossed to the Rosa26-Cre-ERT2 mouse line described previously46. The p53 cko line was described previously30. Homozygous p53 cko mice were crossed to the Bcl11a/Cre-ERT line described above and the F1 generation was mated to generate mice doubly conditional for Bcl11a and p53. Genotyping was confirmed using the primers listed in Supplementary Table 2. Cre activation was mediated by three injections of 1 mg tamoxifen per mouse over 3 days.
Mammary epithelial cell isolation and analysis
Mammary epithelial cells were dissociated using a mixture of collagenase (Roche) and hyaluronidase (Sigma), and cells were stained using the following primary antibodies: biotinylated anti-CD45 (clone 30-F11; eBioscience, 1:500), anti-Ter119 (clone Ter119; eBioscience, 1:500) and anti-CD31 (clone 390; eBioscience, 1:500); anti-CD24-R-phycoerythrin (PE; clone M1/69, eBioscience, 1:500), anti-CD49f-Alexa Fluor 647 (AF647; clone GoH3, eBioscience, 1:100), anti-CD49b-Alexa Fluor 488 (AF488; clone HMa2; eBioscience, 1:500) and Sca1-Alexa Fluor 647 (AF647; clone D7, eBioscience, 1:500). Secondary antibodies used: Strepavidin-PE-Texas Red (PE-TR; Molecular Probes, 1:500). Apoptotic cells were excluded by elimination of propidium iodide-positive cells. Flow cytometric analysis was done using CyAN ADP (DakoCytomation) and all sorts were performed using MoFlo (DakoCytomation) and gates were set to exclude >99.9% of cells labelled with isoform-matched control antibodies conjugated with the corresponding fluorochromes. For whole-mount analysis, abdominal glands (no. 4) were spread out using forceps on a glass slide and incubated in Carnoy’s fixative overnight. The slide was washed in water and placed in carmine alum (Sigma) stain overnight. The slide was again washed with ethanol and cleared in Xylene for 1 day before documentation. For histological analysis, abdominal glands were fixed in 4% formaldehyde in PBS for 24 h at room temperature. The glands were transferred to 70% ethanol and stored at −20 °C until embedding and sectioning. All tissues were embedded in wax and sectioned at 5 μm before being stained with haematoxylin and eosin.
Mammary CFC assay
For colony-forming assays, the medium used was (human) NeuroCult NS-A Proliferation Medium (StemCell) supplemented with 5% fetal bovine serum, 10 ηg ml−1 epidermal growth factor (Sigma), 10 ng ml−1 basic fibroblast growth factor (Peprotech) and N2 Supplement (Invitrogen); the cultures were maintained at 37 °C/5% CO2 for 7 days; then fixed using ice-cold acetone/methanol (1:1) and visualized using Giemsa staining (Merck). Lin−CD24hiCD49b+ luminal progenitors from the flox/flox mammary gland were sorted and plated with irradiated feeders in colony-forming assay medium for 6 days before the number of mammary CFCs was enumerated.
Transplantation of mammary epithelium
Mammary epithelial cells (basal fraction) from tamoxifen-injected and non-injected flox/flox or flox/+ mice were sorted based on CD24/CD49f and transplanted in limiting doses (500/750/1,000/2,000 cells) into cleared fat pads of 3-week-old NSG females. In each case, non-injected and tamoxifen-injected epithelial cells were engrafted into contralateral glands of the same recipient mice. The recipient mice were impregnated 3–6 weeks after transplant and outgrowths produced were dissected, stained with carmine and scored. Stem cell frequency was calculated using L-Calc (StemCell Technologies).
DMBA/MPA tumourigenesis protocol
Mammary fragments were transplanted into cleared fat pads of 3-week-old NSG mice. At the time of surgery, the MPA slow release pellet (Innovative Research of America) was also implanted subcutaneously. The mice were allowed to recover for 2 weeks and then Bcl11a deletion was induced using three injections of tamoxifen. One week after deletion of Bcl11a, 1 mg of DMBA (Sigma) was administered orally; this was followed by three further doses of 1 mg of DMBA over 3 weeks. Mice were then examined weekly for tumour incidence and killed when tumours reached the legal limit.
Transfection and mammosphere assays
EpH4 (gift from Professor Christine Watson) and MDA231 (ATCC) cells were cultured to confluence in 1:1 DMEM:F12 (Invitrogen) media containing 10% fetal calf serum (FCS; FetalcloneIII, Clonetech). 4T1 (ATCC) cells were cultured in Roswell Park Memorial Institute (RPMI) media (Invitrogen) containing 10% FCS, and SUM159 cells (gift from Dr Charlotte Kuperwasser) were cultured in Ham's F12 (Sigma), 5% FCS, insulin (5 μg ml−1, Sigma) hydrocortisone (1 μg ml−1, Sigma) and 1 × Penicillin Streptomycin Glutamine (PSG) (Gibco). HMLE and HMLER cells (gift from Professor Robert Weinberg) were cultured in complete HuMEC media (Invitrogen). The control or the Bcl11a overexpression piggyBac vectors were delivered into cells using the Amaxa Basic Nucleofactor Kit for primary mammalian epithelial cells (Lonza) according to the manufacturer’s recommendations. Transfected cells were maintained at 37 °C/5% CO2 for 48 h. Cells were then cultured in puromycin (1–5 μg ml−1) for 48 h to allow for selection. To induce BCL11A expression in EpH4 and HMLE cells, doxycycline (Clonetech) was used at a final concentration of 1.0 μg ml−1. Floating or Matrigel-embedded mammosphere were cultured and passaged as previously described47 in ultra-low attachment plates (Corning).
RNA knockdown
BCL11A shRNA sequences were obtained from the TRC consortium48 (TRCN0000033449 and TRCN0000033453) were cloned into piggyBac transposon vector (PB-H1-shRNA-GFP). 4T1, SUM159, MDA231 and HMLER cells were transfected with 4.0 μg of piggyBac vector using Amaxa Basic Nucleofactor Kit for primary mammalian epithelial cells (Lonza) and GFP+ cells were sorted/analysed 24–48 h later.
RNA extraction and real-time PCR analysis
RNA from sorted cells was extracted using PicoPure RNA isolation kit (Molecular Devices) according to the manufacturer’s instructions. RNA from mammary tissue and cell lines was extracted using Tri-Reagent (Invitrogen) according to the manufacturer’s instructions. Complementary DNA (cDNA) was synthesized from 1 to 2 μg of total RNA using the Transcriptor Reverse Transcription cDNA Synthesis Kit (Roche). RT–PCR was performed using Hi-Fidelity Extensor mix (Thermo) using primers listed in Supplementary Table 2. Quantitative real-time PCR detection of cDNA was performed using SYBR Green Master Mix (Sigma, ABI and Invitrogen) according to supplier’s recommendations. The real-time PCR reactions were run in ABI-7900HT (Applied Biosystems) in triplicate. Primers used for real-time PCR on mouse samples were designed using PrimerBank49 website ( http://pga.mgh.harvard.edu/primerbank/) and listed in Supplementary Table 2. All primers were purchased from Sigma-Aldrich. For real-time PCR on human samples Taqman gene expression probes (Life Technologies) were used.
Cell cycle analysis
A total of 150,000 control or BCL11A knockdown cells were seeded in six-well plates and allowed to recover for 48 h. Cells were then incubated with 5 μM Edu (Invitrogen) for 1 h. Cells were fixed and assayed using the EdU flow cytometery detection kit (Invitrogen) following the manufacturer’s instructions.
Annexin v assays
A total of 100,000 control or BCL11A knockdown cells (in triplicates) were seeded in six-well plates and allowed to recover for 48 h. Cell were then collected and stained using the Annexin-V-AF647 (BioLegend) following the manufacturer’s instructions, and cells were then quantified using fluorescence-activated cell sorting.
Cell viability assay
A total of 1,000 control or BCL11A knockdown cells (in triplicates) were seeded in 96-well plates and allowed to recover for 48 h. Cells were then incubated with CellTiter Aqueous One Solution (Promega) for 4 h following the manufacturer’s instructions. Absorbance was then measured at 490 nm using a plate reader (Bio-Rad).
Western blotting and IHC
Protein samples were prepared as described previously14 and probed using anti-Bcl11a (Abcam, Clone 14B5, 1:1000) and Actin (Cell Signalling, 1:10000). For IHC analysis, BCL11A (Abcam (14B5, 1:50); CK14 (Abcam; 1:100) and ERα (SCBT; 1:50) were used. Staining was detected using AF488- or Cy3-conjugated secondary (Sigma) and bisbenzimide-Hoechst 33342 (Sigma). Fluorescence microscopy was carried out using a Zeiss Axiophot microscope equipped with a Hamamatsu Orca 285 camera, with images visualized, captured and manipulated using Simple PCI 6 (C Imaging Systems). The hematoxylin- and eosin-stained samples were visualized on a LEICA light microscope, while the mouse mammary gland whole mounts were visualized using the LEICA MZ75 light microscope.
Microarray analysis
The intensity value for each probe set was calculated and the average of each gene was computed before the data analysis. For the quality control (QC) step, a set of intensity value of control genes were examined. All data were normalized and scaled by Partek Genomic Suite 6.4. Principal components analysis was performed to show the distribution of samples, eliminating outliers. Differentially expressed genes were selected by one-way analysis of variance by the factor of KO versus wild type, P value<0.08. Hierarchical clustering of selected genes was performed to show the expression pattern. The resulting genes then underwent a pathway analysis (GeneGO: http://www.genego.com) to determine the biological significance of the data.
Xenograft tumourigenesis assays
One hundred thousand EpH4, HMLE, 4T1, MDA231, SUM159 or HMLER cells were suspended in 25% Matrigel (BD Biosciences) and HBSS, and injected into either cleared contralateral number 4 mammary fat pads of 3-week-old female mice or subcutaneously in 6–12-week-old female NSG mice. For secondary transplants, tumours were dissociated using collagenase/hyloronase mix (Roche) for 16 h and viable cells were counted and injected into NSG recipient mice at the indicated doses.
METABRIC analysis
Matched DNA and RNA were extracted for tumours. CN analysis was performed using the Affymetrix SNP 6.0 platform. The arrays were pre-processed and normalized using CRMAv2 (ref. 50) method from aroma.affymetrix. Allelic-crosstalk calibration, probe sequence effects normalization, probe-level summarization and PCR fragment length normalization were performed for each array. The intensities obtained were normalized against a pool of 473 normals for the samples with no matched pair or against their matched normal when available (258 samples). The log-ratios were then segmented using the circular binary segmentation algorithm51 in the DNAcopy Bioconductor package52. Then, callings into five groups (homozygous deletion, heterozygous deletion, neutral CN, gain (>2) and amplification >3) were made using thresholds based on the variability of each sample and their proportion of tumoural cells. RNA analysis was performed using Illumina HT-12 v3 platform and analysed using beadarray package53. BASH54 was used to correct for spatial artifacts. The bead-level data were summarized and a selection of suitable probes based on their quality was done using the re-annotation of the Illumina HT-12v3 platform55. The samples were classified into the five breast cancer subtypes using PAM50 (ref. 56), but only those genes with a probe with perfect annotation on the chip were considered. A mixture model was used to classify BCL11A expression into low and high values57,58.
TCGA data analysis
All TCGA data and figures were accessed, analysed and generated using the cBio Cancer Genomics Portal59. All data included in this manuscript is in agreement with the TCGA publication guidelines.
METABRIC IHC analysis
A subset of patients enroled in the METABRIC study with tumour samples represented in tissue microarrays (TMAs) were included for the detection of BCL11A protein expression by IHC. TMAs were constructed from formalin-fixed paraffin-embedded tumour blocks as previously described60. Each tumour was represented by a single 0.6-mm tissue core. A total of 439 tumours were included arising from 436 patients (three were synchronous tumours arising in the contralateral breast). CN and gene expression data was available for 368 of these tumours for correlative analyses. Three micrometre TMA sections were dewaxed in xylene and rehydrated through graded alcohols. IHC was conducted using a BondMaX Autoimmunostainer (Leica, Bucks, UK). Antigen retrieval was achieved by heating TMA sections in pH 6 citrate buffer for 20 min. Primary mouse monoclonal (clone 14B5) antibody bound to BCL11A (ab19487, AbCam) was diluted to 1:200 and detected using a BOND Polymer detection kit (Leica) and signal developed with 3,3′-diaminobenzidine (DAB). Stained TMA sections were digitized using the Ariol (Genetix Ltd, Hampshire, UK) platform for scoring by a pathologist (H.R.A.). The ordinal Allred scoring system was used for assessing the amount of staining present in tumour cells accounting for the intensity (0=no staining, 1=weak, 2=moderate and 3=strong) and proportion (0=0%, 1=<1%, 2=1–10%, 3=11–33%, 4=34–66% and 5=>66%) of stained cells, finally producing a summed score (intensity+proportion=Allred score) between zero and eight. Analogous to clinical practice for estrogen receptor (ER), tumours with an Allred score of >2 were deemed positive for BCL11A expression and comparison with molecular subtypes was made using Pearson’s χ2 test.
Statistical significance
All P values were calculated using Student’s t-test unless otherwise indicated in the figure legends.
Additional information
How to cite this article: Khaled, W.T. et al. BCL11A is a triple-negative breast cancer gene with critical functions in stem and progenitor cells. Nat. Commun. 6:5987 doi: 10.1038/ncomms6987 (2015).
Accession codes: Microarray data have been deposited in the Gene Expression Omnibus database with the accession number GSE63386.
Accession codes
References
Blows, F. M. et al. Subtyping of breast cancer by immunohistochemistry to investigate a relationship between subtype and short and long term survival: a collaborative analysis of data for 10,159 cases from 12 studies. PLoS Med. 7, e1000279 (2010).
Foulkes, W. D., Smith, I. E. & Reis-Filho, J. S. Triple-negative breast cancer. N. Engl. J. Med. 363, 1938–1948 (2011).
Perou, C. M. et al. Molecular portraits of human breast tumours. Nature 406, 747–752 (2000).
Carey, L., Winer, E., Viale, G., Cameron, D. & Gianni, L. Triple-negative breast cancer: disease entity or title of convenience? Nat. Rev. Clin. Oncol. 7, 683–692 (2010).
Curtis, C. et al. The genomic and transcriptomic architecture of 2,000 breast tumours reveals novel subgroups. Nature 486, 346–352 (2012).
Prat, A. et al. Phenotypic and molecular characterization of the claudin-low intrinsic subtype of breast cancer. Breast Cancer Res. 12, R68 (2010).
Weigelt, B., Baehner, F. L. & Reis-Filho, J. S. The contribution of gene expression profiling to breast cancer classification, prognostication and prediction: a retrospective of the last decade. J. Pathol. 220, 263–280 (2010).
TCGA. Comprehensive molecular portraits of human breast tumours. Nature 490, 61–70 (2012).
Shah, S. P. et al. The clonal and mutational evolution spectrum of primary triple-negative breast cancers. Nature 486, 395–399 (2012).
Stingl, J. & Caldas, C. Molecular heterogeneity of breast carcinomas and the cancer stem cell hypothesis. Nat. Rev. Cancer 7, 791–799 (2007).
Bouras, T. et al. Notch signaling regulates mammary stem cell function and luminal cell-fate commitment. Cell Stem Cell 3, 429–441 (2008).
Kouros-Mehr, H., Slorach, E. M., Sternlicht, M. D. & Werb, Z. GATA-3 maintains the differentiation of the luminal cell fate in the mammary gland. Cell 127, 1041–1055 (2006).
Asselin-Labat, M. L. et al. Gata-3 is an essential regulator of mammary-gland morphogenesis and luminal-cell differentiation. Nat. Cell Biol. 9, 201–209 (2007).
Khaled, W. T. et al. The IL-4/IL-13/Stat6 signalling pathway promotes luminal mammary epithelial cell development. Development 134, 2739–2750 (2007).
Zuo, T. et al. FOXP3 is an X-linked breast cancer suppressor gene and an important repressor of the HER-2/ErbB2 oncogene. Cell 129, 1275–1286 (2007).
Mehra, R. et al. Identification of GATA3 as a breast cancer prognostic marker by global gene expression meta-analysis. Cancer Res. 65, 11259–11264 (2005).
Desmedt, C. et al. Strong time dependence of the 76-gene prognostic signature for node-negative breast cancer patients in the TRANSBIG multicenter independent validation series. Clin. Cancer Res. 13, 3207–3214 (2007).
Hatzis, C. et al. A genomic predictor of response and survival following taxane-anthracycline chemotherapy for invasive breast cancer. JAMA 305, 1873–1881 (2011).
Ma, X. J. et al. A two-gene expression ratio predicts clinical outcome in breast cancer patients treated with tamoxifen. Cancer Cell 5, 607–616 (2004).
Schmidt, M. et al. The humoral immune system has a key prognostic impact in node-negative breast cancer. Cancer Res. 68, 5405–5413 (2008).
Weniger, M. A. et al. Gains of the proto-oncogene BCL11A and nuclear accumulation of BCL11A(XL) protein are frequent in primary mediastinal B-cell lymphoma. Leukemia. 20, 1880–1882 (2006).
Nakamura, T. et al. Evi9 encodes a novel zinc finger protein that physically interacts with BCL6, a known human B-cell proto-oncogene product. Mol. Cell Biol. 20, 3178–3186 (2000).
Reichmann, E., Ball, R., Groner, B. & Friis, R. R. New mammary epithelial and fibroblastic cell clones in coculture form structures competent to differentiate functionally. J. Cell Biol. 108, 1127–1138 (1989).
Ince, T. A. et al. Transformation of different human breast epithelial cell types leads to distinct tumor phenotypes. Cancer Cell 12, 160–170 (2007).
Elenbaas, B. et al. Human breast cancer cells generated by oncogenic transformation of primary mammary epithelial cells. Genes Dev. 15, 50–65 (2001).
Ishikawa, F. et al. Development of functional human blood and immune systems in NOD/SCID/IL2 receptor {gamma} chain(null) mice. Blood 106, 1565–1573 (2005).
Liu, P. et al. Bcl11a is essential for normal lymphoid development. Nat. Immunol. 4, 525–532 (2003).
Herschkowitz, J. I. et al. Identification of conserved gene expression features between murine mammary carcinoma models and human breast tumors. Genome Biol. 8, R76 (2007).
Herschkowitz, J. I. et al. Comparative oncogenomics identifies breast tumors enriched in functional tumor-initiating cells. Proc. Natl Acad. Sci. USA 109, 2778–2783 (2012).
Jonkers, J. et al. Synergistic tumor suppressor activity of BRCA2 and p53 in a conditional mouse model for breast cancer. Nat. Genet. 29, 418–425 (2001).
Hennighausen, L. & Robinson, G. W. Information networks in the mammary gland. Nat. Rev. Mol. Cell Biol. 6, 715–725 (2005).
Stingl, J. et al. Purification and unique properties of mammary epithelial stem cells. Nature 439, 993–997 (2006).
Shackleton, M. et al. Generation of a functional mammary gland from a single stem cell. Nature 439, 84–88 (2006).
Lim, E. et al. Transcriptome analyses of mouse and human mammary cell subpopulations reveal multiple conserved genes and pathways. Breast Cancer Res. 12, R21 (2010).
Van Keymeulen, A. et al. Distinct stem cells contribute to mammary gland development and maintenance. Nature 479, 189–193 (2011).
Rios, A. C., Fu, N. Y., Lindeman, G. J. & Visvader, J. E. In situ identification of bipotent stem cells in the mammary gland. Nature 506, 322–327 (2014).
Lim, E. et al. Aberrant luminal progenitors as the candidate target population for basal tumor development in BRCA1 mutation carriers. Nat. Med. 15, 907–913 (2009).
Molyneux, G. et al. BRCA1 basal-like breast cancers originate from luminal epithelial progenitors and not from basal stem cells. Cell Stem Cell 7, 403–417 (2010).
Zhou, D., Liu, K., Sun, C. W., Pawlik, K. M. & Townes, T. M. KLF1 regulates BCL11A expression and gamma- to beta-globin gene switching. Nat. Genet. 42, 742–744 (2010).
Jiang, B. Y. et al. BCL11A overexpression predicts survival and relapse in non-small cell lung cancer and is modulated by microRNA-30a and gene amplification. Mol. Cancer 12, 61 (2013).
Senawong, T., Peterson, V. J. & Leid, M. BCL11A-dependent recruitment of SIRT1 to a promoter template in mammalian cells results in histone deacetylation and transcriptional repression. Arch. Biochem. Biophys. 434, 316–325 (2005).
Yin, B. et al. A retroviral mutagenesis screen reveals strong cooperation between Bcl11a overexpression and loss of the Nf1 tumor suppressor gene. Blood 113, 1075–1085 (2008).
Yu, Y. et al. Bcl11a is essential for lymphoid development and negatively regulates p53. J. Exp. Med. 209, 2467–2483 (2012).
Chan, W. et al. A recombineering based approach for high-throughput conditional knockout targeting vector construction. Nucleic Acids Res. 35, e64 (2007).
Liu, P., Jenkins, N. A. & Copeland, N. G. A highly efficient recombineering-based method for generating conditional knockout mutations. Genome Res. 13, 476–484 (2003).
Hameyer, D. et al. Toxicity of ligand-dependent Cre recombinases and generation of a conditional Cre deleter mouse allowing mosaic recombination in peripheral tissues. Physiol. Genomics 31, 32–41 (2007).
Dontu, G. et al. In vitro propagation and transcriptional profiling of human mammary stem/progenitor cells. Genes Dev. 17, 1253–1270 (2003).
Moffat, J. et al. A lentiviral RNAi library for human and mouse genes applied to an arrayed viral high-content screen. Cell 124, 1283–1298 (2006).
Wang, X. & Seed, B. A PCR primer bank for quantitative gene expression analysis. Nucleic Acids Res. 31, e154 (2003).
Bengtsson, H., Wirapati, P. & Speed, T. P. A single-array preprocessing method for estimating full-resolution raw copy numbers from all Affymetrix genotyping arrays including GenomeWideSNP 5 & 6. Bioinformatics 25, 2149–2156 (2009).
Venkatraman, E. S. & Olshen, A. B. A faster circular binary segmentation algorithm for the analysis of array CGH data. Bioinformatics 23, 657–663 (2007).
Seshan VE, O. A. DNAcopy: DNA copy number data analysis R package http://www.bioconductor.org/packages/2.3/bioc/html/DNAcopy.html (2010).
Dunning, M. J., Smith, M. L., Ritchie, M. E. & Tavare, S. beadarray: R classes and methods for Illumina bead-based data. Bioinformatics 23, 2183–2184 (2007).
Cairns, J. M., Dunning, M. J., Ritchie, M. E., Russell, R. & Lynch, A. G. BASH: a tool for managing BeadArray spatial artefacts. Bioinformatics 24, 2921–2922 (2008).
Barbosa-Morais, N. L. et al. A re-annotation pipeline for Illumina BeadArrays: improving the interpretation of gene expression data. Nucleic Acids Res. 38, e17.
Parker, J. S. et al. Supervised risk predictor of breast cancer based on intrinsic subtypes. J. Clin. Oncol. 27, 1160–1167 (2009).
Yeung, K. Y., Fraley, C., Murua, A., Raftery, A. E. & Ruzzo, W. L. Model-based clustering and data transformations for gene expression data. Bioinformatics 17, 977–987 (2001).
Fraley, C. & Raftery, A. E. MCLUST Version 3 for R: Normal Mixture Modeling and Model-based Clustering.Technical Report No. 504, Department of Statistics, University of Washington (2006).
Cerami, E. et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2, 401–404 (2012).
Kononen, J. et al. Tissue microarrays for high-throughput molecular profiling of tumor specimens. Nat. Med. 4, 844–847 (1998).
Acknowledgements
W.T.K., S.C.L. and J.W. were funded by the Sanger Institute. W.T.K. was also funded by a BBSRC project grant and a Junior Research Fellowship, King’s College, Cambridge and a Cancer Research UK Career establishment award. J.S. is funded by Cancer Research UK, The University of Cambridge and Hutchison Whampoa Limited. C.J.W. is funded by BBSRC, MRC and BCC. We would like to thank Dr Floris Foijer and Professor Allan Bradley for providing the p53 cko mice. We thank Dr Shannon Burke, Dr Sara Pensa and Lauma Skruzmane for comments on the manuscript. We thank Miss Malgorzata Gawedzka from the W.T.K. lab for the technical assistance. We thank the Sanger Institute RSF, the flow cytometry core and microarray facilities for the technical assistance. This work is supported by Wellcome Trust (Grant number—098051) (P.L.).
Author information
Authors and Affiliations
Contributions
W.T.K. performed the mouse tumour studies, the cell culture experiments and analysed the Bcl11a cko line. S.C.L. generated and analysed the Bcl11a-LacZ mouse line and the Bcl11a cko line. J.S. developed the flow-sorting strategies, performed some of the colony assays, designed and performed some of the fat pad transplantation experiments. X.C. performed the microarray analysis. O.M.R., H.R.A., S.-F.C., S.A. and C.C. analysed and provided all the data (CN, expression, qRT–PCR and IHC) from the METABRIC cohort. All enquiries relating to the METABRIC data set should be made directly to carlos.caldas@cancer.org.uk. J.W. assisted with mouse experiments. Y.Y. assisted with qRT–PCR and cloning of shRNA constructs. A.F. and M.S. provided the reagents. The Bcl11a cko allele was made by P.L. in laboratory of N.G.C. and N.A.J. C.J.W. provided the reagents and contributed to the design of the study. W.T.K., S.C.L. and P.L. designed the studies and wrote the manuscript. The overall research project and manuscript writing are supervized by P.L.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Information
Supplementary Figures 1-9 and Supplementary Tables 1-2 (PDF 1396 kb)
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Khaled, W., Choon Lee, S., Stingl, J. et al. BCL11A is a triple-negative breast cancer gene with critical functions in stem and progenitor cells. Nat Commun 6, 5987 (2015). https://doi.org/10.1038/ncomms6987
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/ncomms6987
This article is cited by
-
Silencing Notch4 promotes tumorigenesis and inhibits metastasis of triple-negative breast cancer via Nanog and Cdc42
Cell Death Discovery (2023)
-
RNA-seq analysis reveals candidate genes associated with proliferation, invasion, and migration in BCL11A knockdown B-NHL cell lines
Annals of Hematology (2023)
-
Deciphering clinical significance of BCL11A isoforms and protein expression roles in triple-negative breast cancer subtype
Journal of Cancer Research and Clinical Oncology (2023)
-
A phase I/II study of preoperative letrozole, everolimus, and carotuximab in stage 2 and 3 hormone receptor-positive and Her2-negative breast cancer
Breast Cancer Research and Treatment (2023)
-
Conjugated polymer nanoparticles and their nanohybrids as smart photoluminescent and photoresponsive material for biosensing, imaging, and theranostics
Microchimica Acta (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
