
Abstract
In mammals, taste buds typically contain 50–100 tightly packed taste-receptor cells (TRCs), representing all five basic qualities: sweet, sour, bitter, salty and umami1,2. Notably, mature taste cells have life spans of only 5–20 days and, consequently, are constantly replenished by differentiation of taste stem cells3. Given the importance of establishing and maintaining appropriate connectivity between TRCs and their partner ganglion neurons (that is, ensuring that a labelled line from sweet TRCs connects to sweet neurons, bitter TRCs to bitter neurons, sour to sour, and so on), we examined how new connections are specified to retain fidelity of signal transmission. Here we show that bitter and sweet TRCs provide instructive signals to bitter and sweet target neurons via different guidance molecules (SEMA3A and SEMA7A)4,5,6. We demonstrate that targeted expression of SEMA3A or SEMA7A in different classes of TRCs produces peripheral taste systems with miswired sweet or bitter cells. Indeed, we engineered mice with bitter neurons that now responded to sweet tastants, sweet neurons that responded to bitter or sweet neurons responding to sour stimuli. Together, these results uncover the basic logic of the wiring of the taste system at the periphery, and illustrate how a labelled-line sensory circuit preserves signalling integrity despite rapid and stochastic turnover of receptor cells.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


Accession codes
References
Lindemann, B. Receptors and transduction in taste. Nature 413, 219–225 (2001)
Yarmolinsky, D. A., Zuker, C. S. & Ryba, N. J. P. Common sense about taste: from mammals to insects. Cell 139, 234–244 (2009)
Barlow, L. A. & Klein, O. D. Developing and regenerating a sense of taste. Curr. Top. Dev. Biol. 111, 401–419 (2015)
Zipursky, S. L. & Sanes, J. R. Chemoaffinity revisited: dscams, protocadherins, and neural circuit assembly. Cell 143, 343–353 (2010)
Yoshida, Y. Semaphorin signaling in vertebrate neural circuit assembly. Front. Mol. Neurosci. 5, 71 (2012)
Pasterkamp, R. J. Getting neural circuits into shape with semaphorins. Nat. Rev. Neurosci. 13, 605–618 (2012)
Wang, F., Nemes, A., Mendelsohn, M. & Axel, R. Odorant receptors govern the formation of a precise topographic map. Cell 93, 47–60 (1998)
Komiyama, T. & Luo, L. Development of wiring specificity in the olfactory system. Curr. Opin. Neurobiol. 16, 67–73 (2006)
Zhao, G. Q. et al. The receptors for mammalian sweet and umami taste. Cell 115, 255–266 (2003)
Mueller, K. L. et al. The receptors and coding logic for bitter taste. Nature 434, 225–229 (2005)
Hayashi, S. & Takeichi, M. Emerging roles of protocadherins: from self-avoidance to enhancement of motility. J. Cell Sci. 128, 1455–1464 (2015)
Zipursky, S. L. & Grueber, W. B. The molecular basis of self-avoidance. Annu. Rev. Neurosci. 36, 547–568 (2013)
Koropouli, E. & Kolodkin, A. L. Semaphorins and the dynamic regulation of synapse assembly, refinement, and function. Curr. Opin. Neurobiol. 27, 1–7 (2014)
Coate, T. M., Spita, N. A., Zhang, K. D., Isgrig, K. T. & Kelley, M. W. Neuropilin-2/semaphorin-3F-mediated repulsion promotes inner hair cell innervation by spiral ganglion neurons. eLife 4, e07830 (2015)
Tran, T. S. et al. Secreted semaphorins control spine distribution and morphogenesis in the postnatal CNS. Nature 462, 1065–1069 (2009)
Luo, Y., Raible, D. & Raper, J. A. Collapsin: a protein in brain that induces the collapse and paralysis of neuronal growth cones. Cell 75, 217–227 (1993)
Messersmith, E. K. et al. Semaphorin III can function as a selective chemorepellent to pattern sensory projections in the spinal cord. Neuron 14, 949–959 (1995)
Song, H. et al. Conversion of neuronal growth cone responses from repulsion to attraction by cyclic nucleotides. Science 281, 1515–1518 (1998)
Polleux, F., Morrow, T. & Ghosh, A. Semaphorin 3A is a chemoattractant for cortical apical dendrites. Nature 404, 567–573 (2000)
Telley, L. et al. Dual function of NRP1 in axon guidance and subcellular target recognition in cerebellum. Neuron 91, 1276–1291 (2016)
Barretto, R. P. et al. The neural representation of taste quality at the periphery. Nature 517, 373–376 (2015)
Wu, A., Dvoryanchikov, G., Pereira, E., Chaudhari, N. & Roper, S. D. Breadth of tuning in taste afferent neurons varies with stimulus strength. Nat. Commun. 6, 8171 (2015)
Taniguchi, M. et al. Disruption of semaphorin III/D gene causes severe abnormality in peripheral nerve projection. Neuron 19, 519–530 (1997)
Merte, J. et al. A forward genetic screen in mice identifies Sema3AK108N, which binds to neuropilin-1 but cannot signal. J. Neurosci. 30, 5767–5775 (2010)
Carcea, I. et al. Maturation of cortical circuits requires semaphorin 7A. Proc. Natl Acad. Sci. USA 111, 13978–13983 (2014)
Pasterkamp, R. J., Peschon, J. J., Spriggs, M. K. & Kolodkin, A. L. Semaphorin 7A promotes axon outgrowth through integrins and MAPKs. Nature 424, 398–405 (2003)
Langley, J. N. Note on regeneration of prae-ganglionic fibres of the sympathetic. J. Physiol. (Lond.) 18, 280–284 (1895)
Sperry, R. W. Chemoaffinity in the orderly growth of nerve fiber patterns and connections. Proc. Natl Acad. Sci. USA 50, 703–710 (1963)
Sanes, J. R. & Yamagata, M. Many paths to synaptic specificity. Annu. Rev. Cell Dev. Biol. 25, 161–195 (2009)
Zaidi, F. N. & Whitehead, M. C. Discrete innervation of murine taste buds by peripheral taste neurons. J. Neurosci. 26, 8243–8253 (2006)
Oka, Y., Butnaru, M., von Buchholtz, L., Ryba, N. J. P. & Zuker, C. S. High salt recruits aversive taste pathways. Nature 494, 472–475 (2013)
Zhang, Y. et al. Coding of sweet, bitter, and umami tastes: different receptor cells sharing similar signaling pathways. Cell 112, 293–301 (2003)
Yarmolinsky, D. A. et al. Coding and plasticity in the mammalian thermosensory system. Neuron 92, 1079–1092 (2016)
Chandrashekar, J. et al. The taste of carbonation. Science 326, 443–445 (2009)
Vandenbeuch, A. et al. Postsynaptic P2X3-containing receptors in gustatory nerve fibres mediate responses to all taste qualities in mice. J. Physiol. (Lond.) 593, 1113–1125 (2015)
Acknowledgements
We thank T. Yagi for providing the floxed Sema3a mice, R. Barretto and D. Yarmolinsky for help with in vivo imaging and analysis, and Z. Wu, I. Schieren and S. O’Keeffe for advice and expert technical support. We also thank N. Balaskas and the members of the Zuker laboratory for helpful comments and suggestions. Research reported in this publication was supported by the National Institute On Drug Abuse of the National Institutes of Health under Award Number R01DA035025 (C.S.Z.) and in part by the Intramural Research Program of the NIH, NIDCR (N.J.P.R.). L.J.M. was an HHMI Fellow of the Jane Coffin Childs Memorial Fund. C.S.Z. is an investigator of the Howard Hughes Medical Institute and a Senior Fellow at Janelia Farm Research Campus.
Author information
Authors and Affiliations
Contributions
H.L. and L.J.M. designed the study, carried out the experiments, analysed data and wrote the paper, C.A.P. performed experiments, C.S.Z. and N.J.P.R. designed the study, analysed data and wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Figure 1 Expression profiling of fluorescently labelled TRC populations identifies candidate signalling molecules enriched in bitter TRCs.
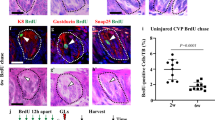
a, Confocal microscopy images of circumvallate papilla taste buds from a TRPM5–BFP (labelling sweet and bitter TRCs32, pseudo-coloured red); T2R32–GFP (labelling bitter TRCs) double-labelled transgenic mouse. These are the animals used in the profiling of sweet/umami and bitter TRCs. Note the expected co-expression of bitter taste receptors and a subset of TRPM5+ cells (right; TRPM5+, but T2R− TRCs are indicated by asterisks). b, Taste bud from a TRPM5–BFP transgenic mouse showing co-expression of the BFP reporter (pseudo-coloured red) and PLCβ2 (a marker of sweet, umami and bitter TRCs32, green). c, Taste bud from a PKD2L1–TdTomato transgenic animal demonstrating expression of the TdTomato reporter (red) in sour TRCs (marked by CAR4 (ref. 34), green). d, qRT–PCR illustrating the relative expression of several candidate connectivity molecules in bitter, sweet and sour TRCs. Data are presented as mean + s.e.m. relative to the expression levels in the whole circumvallate papilla (referred to as taste tissue); n = 3.
Extended Data Figure 2 Expression of semaphorin receptors in geniculate ganglion neurons.
a, qRT–PCR analysis showing expression of candidate SEMA3A receptors5,6 (Nrp1, Plxna3 and Plxna4) and SEMA7A receptors5,6 (Itgb1 and Plxnc1) in the geniculate ganglia. P2x3 (also known as P2rx3) and T1r3 were used as controls for geniculate ganglia35 and TRC9 tissue integrity, respectively. Nrp1, neuropilin 1; Plxna3, plexin A3; Plxna4, plexin A4; Itgb1, integrin β1; Plxnc1, plexin C1. Data are presented for each gene as its relative abundance in the ganglia compared to TRCs. Values are mean + s.e.m. (n = 3). b–d, Confocal microscopy images of immunostaining illustrating the expression of NRP1 (c) and PLXNC1 (d) in subsets of geniculate ganglion neurons. b, The anatomy of the ganglion highlighting the location of fibre tracks.
Extended Data Figure 3 Targeted misexpression of SEMA3A and SEMA7A.
a, Confocal microscopy images of circumvallate papilla taste buds from a T1r3::SEMA3A mouse demonstrating transgene expression (marked by Flag tag; green) in T1R3-expressing TRCs (red). b, The SEMA7A transgene of T2r::SEMA7A animals (green) is expressed in a subset of TRCs not overlapping with T1R3 (red). c, SEMA7A (green) in Pkd2l1::SEMA7A animals is expressed in sour TRCs (marked by CAR4, red).
Extended Data Figure 4 Responses of ganglion neurons in T1r3::SEMA3A mice.
a, Dose responses to sweet and bitter in control and T1r3::SEMA3A mice. The number of cells that respond at each stimulus concentration is shown, cells are classified by their response profile (that is, sweet-responding, bitter-responding or sweet-bitter responding). Note that the tuning profiles are maintained at all three sweet and bitter concentrations, including extremely high concentrations of bitter10 and sweet9. Control, n = 58 cells; T1r3::SEMA3A, n = 34 cells. b, Responses are similar for different tastants within a modality. Tuning properties of bitter- and sweet-responding neurons in the T1r3::SEMA3A animals to two structurally different bitter- and sweet-tasting chemicals are shown. Quinine (5 mM), cycloheximide (Cyx, 100 μM), AceK (30 mM) and sucrose (300 mM). Control, n = 178 bitter-responding cells, n = 227 sweet-responding cells (seven mice). T1r3::SEMA3A, n = 130 bitter-responding cells, n = 165 sweet-responding cells (five mice).
Extended Data Figure 5 Aversion to bitter tastants is impaired in T1r3::SEMA3A;Sema3aM/M mice.
a, Dose–response to the bitter quinine in control and Sema3aM/M-mutant mice. Animals were tested using a brief-access lick assay as previously described10. The relative fraction of licks to each concentration of quinine is shown (0.25 mM, 0.5 mM, 1 mM, 2 mM, 4 mM). Control, n = 4; Sema3aM/M, n = 5; mean ± s.d. b, c, Targeting bitter neurons to sweet TRCs results in a significant loss of bitter taste sensitivity (see also Fig. 3f). b, The graphs show the relative fraction of licks to water and to two different concentrations of PROP in control and T1r3::SEMA3A;Sema3aM/M animals. c, The graphs show the relative fraction of licks to water and to two different concentrations of denatonium in control and T1r3::SEMA3A;Sema3aM/M animals. n = 3, mean + s.d., **P < 0.01, Student’s two-tailed, unpaired t-test. d, Chorda tympani whole-nerve recording32 in control and T1r3::SEMA3A;Sema3aM/M-mutant mice. Responses were normalized to 60 mM NaCl (n = 3, mean + s.d.). Note that whole-nerve responses to bitter and sweet are unchanged, probably because nerve recording measures bulk neural activity, further highlighting the importance of recording single-neuron activity; GCaMP-based imaging of ganglion activity is markedly more informative than whole-nerve, or even single-fibre physiological recordings, because it allows simultaneous sampling of large numbers of neurons with single-cell selectivity.
Supplementary information
Supplementary Data
This file contains Supplementary Table 1.
Source data
Rights and permissions
About this article

Cite this article
Lee, H., Macpherson, L., Parada, C. et al. Rewiring the taste system. Nature 548, 330–333 (2017). https://doi.org/10.1038/nature23299
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature23299
This article is cited by
-
Neuroscience of taste: unlocking the human taste code
BMC Neuroscience (2024)
-
The role of the oral microbiome in obesity and metabolic disease: potential systemic implications and effects on taste perception
Nutrition Journal (2023)
-
A brainstem map for visceral sensations
Nature (2022)
-
Gut–brain circuits for fat preference
Nature (2022)
-
Phase 1 study of single-agent WNT974, a first-in-class Porcupine inhibitor, in patients with advanced solid tumours
British Journal of Cancer (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
