
Abstract
The lifestyle of intracellular pathogens, such as malaria parasites, is intimately connected to that of their host, primarily for nutrient supply. Nutrients act not only as primary sources of energy but also as regulators of gene expression, metabolism and growth, through various signalling networks that enable cells to sense and adapt to varying environmental conditions1,2. Canonical nutrient-sensing pathways are presumed to be absent from the causative agent of malaria, Plasmodium3,4,5, thus raising the question of whether these parasites can sense and cope with fluctuations in host nutrient levels. Here we show that Plasmodium blood-stage parasites actively respond to host dietary calorie alterations through rearrangement of their transcriptome accompanied by substantial adjustment of their multiplication rate. A kinome analysis combined with chemical and genetic approaches identified KIN as a critical regulator that mediates sensing of nutrients and controls a transcriptional response to the host nutritional status. KIN shares homology with SNF1/AMPKα, and yeast complementation studies suggest that it is part of a functionally conserved cellular energy-sensing pathway. Overall, these findings reveal a key parasite nutrient-sensing mechanism that is critical for modulating parasite replication and virulence.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others

Accession codes
References
Efeyan, A., Comb, W. C. & Sabatini, D. M. Nutrient-sensing mechanisms and pathways. Nature 517, 302–310 (2015)
Conrad, M. et al. Nutrient sensing and signaling in the yeast Saccharomyces cerevisiae. FEMS Microbiol. Rev. 38, 254–299 (2014)
Babbitt, S. E. et al. Plasmodium falciparum responds to amino acid starvation by entering into a hibernatory state. Proc. Natl Acad. Sci. USA 109, E3278–E3287 (2012)
Miranda-Saavedra, D., Gabaldón, T., Barton, G. J., Langsley, G. & Doerig, C. The kinomes of apicomplexan parasites. Microbes Infect. 14, 796–810 (2012)
Serfontein, J., Nisbet, R. E., Howe, C. J. & de Vries, P. J. Evolution of the TSC1/TSC2-TOR signaling pathway. Sci. Signal. 3, ra49 (2010)
Collino, S. et al. Transcriptomics and metabonomics identify essential metabolic signatures in calorie restriction (CR) regulation across multiple mouse strains. Metabolites 3, 881–911 (2013)
Fontana, L., Meyer, T. E., Klein, S. & Holloszy, J. O. Long-term calorie restriction is highly effective in reducing the risk for atherosclerosis in humans. Proc. Natl Acad. Sci. USA 101, 6659–6663 (2004)
Fontana, L., Partridge, L. & Longo, V. D. Extending healthy life span—from yeast to humans. Science 328, 321–326 (2010)
Meidenbauer, J. J., Ta, N. & Seyfried, T. N. Influence of a ketogenic diet, fish-oil, and calorie restriction on plasma metabolites and lipids in C57BL/6J mice. Nutr. Metab. (Lond.) 11, 23 (2014)
Hunt, N. H., Manduci, N. & Thumwood, C. M. Amelioration of murine cerebral malaria by dietary restriction. Parasitology 107, 471–476 (1993)
Mejia, P. et al. Dietary restriction protects against experimental cerebral malaria via leptin modulation and T-cell mTORC1 suppression. Nat. Commun. 6, 6050 (2015)
Pasini, E. M. et al. Proteomic and genetic analyses demonstrate that Plasmodium berghei blood stages export a large and diverse repertoire of proteins. Mol. Cell. Proteomics 12, 426–448 (2013)
Gasch, A. P. in Yeast Stress Responses (eds Hohmann, S. & Mager, W. H. ) Ch. 2, (Springer, 2003)
Tewari, R. et al. The systematic functional analysis of Plasmodium protein kinases identifies essential regulators of mosquito transmission. Cell Host Microbe 8, 377–387 (2010)
Dorin-Semblat, D., Sicard, A., Doerig, C., Ranford-Cartwright, L. & Doerig, C. Disruption of the PfPK7 gene impairs schizogony and sporogony in the human malaria parasite Plasmodium falciparum. Eukaryot. Cell 7, 279–285 (2008)
Mony, B. M. et al. Genome-wide dissection of the quorum sensing signalling pathway in Trypanosoma brucei. Nature 505, 681–685 (2014)
Bracchi, V., Langsley, G., Thélu, J., Eling, W. & Ambroise-Thomas, P. PfKIN, an SNF1 type protein kinase of Plasmodium falciparum predominantly expressed in gametocytes. Mol. Biochem. Parasitol. 76, 299–303 (1996)
Hardie, D. G., Ross, F. A. & Hawley, S. A. AMP-activated protein kinase: a target for drugs both ancient and modern. Chem. Biol. 19, 1222–1236 (2012)
Crozet, P. et al. Mechanisms of regulation of SNF1/AMPK/SnRK1 protein kinases. Front. Plant Sci. 5, 190 (2014)
Boehme, U. et al. Complete avian malaria parasite genomes reveal host-specific parasite evolution in birds and mammals. Preprint at bioRxivhttp://dx.doi.org/10.1101/086504 (2016)
Stein, S. C., Woods, A., Jones, N. A., Davison, M. D. & Carling, D. The regulation of AMP-activated protein kinase by phosphorylation. Biochem. J. 345, 437–443 (2000)
Hawley, S. A. et al. The ancient drug salicylate directly activates AMP-activated protein kinase. Science 336, 918–922 (2012)
Calabrese, M. F. et al. Structural basis for AMPK activation: natural and synthetic ligands regulate kinase activity from opposite poles by different molecular mechanisms. Structure 22, 1161–1172 (2014)
Xiao, B. et al. Structural basis of AMPK regulation by small molecule activators. Nat. Commun. 4, 3017 (2013)
Pankevich, D. E., Teegarden, S. L., Hedin, A. D., Jensen, C. L. & Bale, T. L. Caloric restriction experience reprograms stress and orexigenic pathways and promotes binge eating. J. Neurosci. 30, 16399–16407 (2010)
Murray, M. J., Murray, N. J., Murray, A. B. & Murray, M. B. Refeeding-malaria and hyperferraemia. Lancet 1, 653–654 (1975)
NCD Risk Factor Collaboration (NCD-RisC). Trends in adult body-mass index in 200 countries from 1975 to 2014: a pooled analysis of 1,698 population-based measurement studies with 19·2 million participants. Lancet 387, 1377–1396 (2016)
Bousema, T., Okell, L., Felger, I. & Drakeley, C. Asymptomatic malaria infections: detectability, transmissibility and public health relevance. Nat. Rev. Microbiol. 12, 833–840 (2014)
Janse, C. J., Ramesar, J. & Waters, A. P. High-efficiency transfection and drug selection of genetically transformed blood stages of the rodent malaria parasite Plasmodium berghei. Nat. Protocols 1, 346–356 (2006)
Franke-Fayard, B. et al. Murine malaria parasite sequestration: CD36 is the major receptor, but cerebral pathology is unlinked to sequestration. Proc. Natl Acad. Sci. USA 102, 11468–11473 (2005)
Liu, Y., Xu, X. & Carlson, M. Interaction of SNF1 protein kinase with its activating kinase Sak1. Eukaryot. Cell 10, 313–319 (2011)
Gietz, R. D. & Schiestl, R. H. Quick and easy yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protocols 2, 35–37 (2007)
Lemieux, J. E. et al. Statistical estimation of cell-cycle progression and lineage commitment in Plasmodium falciparum reveals a homogeneous pattern of transcription in ex vivo culture. Proc. Natl Acad. Sci. USA 106, 7559–7564 (2009)
Kafsack, B. F., Painter, H. J. & Llinás, M. New Agilent platform DNA microarrays for transcriptome analysis of Plasmodium falciparum and Plasmodium berghei for the malaria research community. Malar. J. 11, 187 (2012)
Kim, D. et al. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 14, R36 (2013)
Robinson, M. D., McCarthy, D. J. & Smyth, G. K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2010)
Ritchie, M. E. et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 43, e47 (2015)
McCarthy, D. J., Chen, Y. & Smyth, G. K. Differential expression analysis of multifactor RNA-seq experiments with respect to biological variation. Nucleic Acids Res. 40, 4288–4297 (2012)
Acknowledgements
We thank A. Pena and A. Pamplona for help with animal experiments; S. Marques for help in transfections; A. Parreira for the mosquito preparation; M. Rebelo for help with flow cytometry analysis; A. Waters, T. Hanscheid and F. Enguita for helpful discussions; R. Wall and D. Brady for technical assistance with Δkin clones; and Jacobus Pharmaceuticals for the WR99210 compound. The work was supported by European Research Council (311502) and Fundação para a Ciência e Tecnologia (FCT) grants (EXCL/IMI-MIC/0056/2012) and (PTDC/SAU-MET/118199/2010) to M.M.M. and L.M.S., respectively. L.M.S., A.R.Go. and M.M.M. were supported by the European Commission (FP7/2007-2013; EVIMALAR 242095). L.M.S. was supported by EMBO LTF (ALTF 960-2009). K.S. was sponsored by FCT fellowship (SFRH/BPD/111788/2015). I.M.V. received EMBO (LTF 712-2012) and NIH NRSA (5F32AI104252-03) fellowships. Work at the Sanger Institute was funded by Wellcome Trust (098051) and Medical Research Council (MRC, G0501670). Work at the Instituto Gulbenkian de Ciência was funded through FCT grants (SFRH-BPD79255-2011 and UID/Multi/04551/2013). R.T. was supported by MRC grant (G0900109, MR/K011782/1). M.L. was funded through the Burroughs Wellcome Fund and the NIH Director’s New Innovators award (1DP2OD001315-01).
Author information
Authors and Affiliations
Contributions
L.M.S. and M.M.M. conceived and led the study. O.B., M.L., R.T. and E.B.G. contributed ideas and interpretation. Animal work was conducted by L.M.S., K.S., M.T.G.R., I.M.V. and J.S.D. Parasite transfections and analysis were performed by L.M.S., K.S., M.T.G.R. and A.R.Go. Transcriptomic analysis was carried out by A.R.Gr., K.K.M., C.R.M., L.M.S. and M.T.G.R. Yeast complementation studies were performed by K.S., M.T.G.R. and M.A. AMPK modelling was performed by P.C.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Additional information
Reviewer Information Nature thanks Z. Bozdech, D. Gilson, L. Miller and the other anonymous reviewer(s) for their contribution to the peer review of this work.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Figure 1 Effect of calorie restricted diet in rodent malaria models.
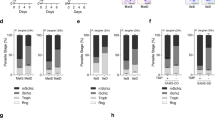
a, b, Body weight change (a) and blood glucose levels (b) of C57BL/6 male mice under long-term calorie restriction (CR) or ad libitum (AL). Body weight data (n = 4 per group) was normalized to the initial weight for each animal. The average daily food consumption for AL and CR mice was 2.93 ± 0.48 g and 1.76 ± 0.11 g, respectively (mean ± s.d.). c, Number of RBCs per ml in infected C57BL/6 male mice in AL and CR diet regimens. Cell density was determined in 1 μl of blood collected from the tail and counted on a haemocytometer chamber (n = 4 per group). d–f, Full course of parasitaemia from infected mice infected by mosquito bite (d) or by intraperitoneal (i.p.) injection of 1 × 106 iRBCs (e, f). Values (mean ± s.e.m.) represent one of two independent experiments. Parasitaemia of GFP-expressing P. berghei ANKA (mosquito bite, n = 5 per group; iRBCs, n = 4 per group) determined by flow cytometry analysis. Parasitaemia of P. berghei K173 (n = 4 per group) obtained by microscopy analysis of blood smears. g, h, Parasitaemia of male BALB/c mice infected i.p. with 1 × 105 P. yoelii 17XNL iRBCs (n = 4 per group) or 1 × 105 P. chabaudi AS iRBCs (n = 3 per group). Parasitaemia obtained by microscopy analysis represents 1 of 2 independent experiments. i, BALB/c mice i.p. injected with 1 × 105 P. berghei ANKA expressing luciferase under a constitutive promotor30 and imaged on day 4 after infection (AL n = 10, CR n = 5; 2 independent experiments pooled). j, k, Parasitaemia of P. berghei ANKA infection in BALB/c (j) and BALB/c scid (severe combined immune deficiency) mice (k). Mice were infected by i.p. injection of 1 × 105 GFP-expressing parasites and parasitaemia was assessed by flow cytometry (mean ± s.e.m.; BALB/c, n = 8 per group; BALB/c scid, AL n = 7, CR n = 8; 2 independent experiments pooled). l, BALB/c mice infected with P. berghei ANKA expressing luciferase under the ama1 schizont-specific promoter30, to allow imaging of the sequestering parasite stage. Mice were imaged 25 h after i.v. infection with purified mature schizonts. AL, top animals; CR, bottom animals. Spleen weight and parasite load measured 72 h after infection with mature schizonts. Parasite load was determined by qPCR analysis of P. berghei 18S rRNA. Spleen weight was normalized to body weight for each mouse (AL n = 3, CR n = 4). m, Relative abundance of circulating young and mature parasites in AL and CR C57BL/6 mice infected with 1 × 106 GFP-expressing P. berghei ANKA iRBCs. Tail blood from infected mice was analysed by flow cytometry. After excluding false GFP-positive events, the total GFP population was separated in low (GFP+) and high (GFP++) GFP-expressing cells, corresponding to young and mature parasites, respectively. The same gates were applied to AL and CR blood samples on days 4, 5, and 6 after infection. The observed marked reduction in the percentage of mature parasites overtime in both diet conditions is indicative of sequestration of infected RBCs. Bars are mean ± s.e.m. (7 mice per group; 2 independent experiments pooled).
Extended Data Figure 2 Effect of CR in vitro.
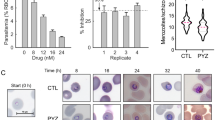
a, Workflow representation of the in vitro parasite maturation assay. Experiments were conducted in a glucose-free RPMI medium supplemented with HEPES, antibiotic, 10–25% rodent sera and glucose at the concentration indicated in the corresponding figure and/or legend. b, c, Box plots showing microscopic quantification of the number of P. berghei ANKA (b) or P. yoelii 17XNL (c) merozoites per segmented schizont after in vitro culturing in the presence of 25% AL or CR mouse sera (glucose, 4 mM). Culture for 30 h showed similarly reduced merozoite numbers in the CR condition, suggesting that parasite development is not delayed in CR. Only segmented schizonts with a single pigmented digestive vacuole were imaged and scored (Mann–Whitney test). Total numbers of schizonts analysed in two independent experiments are as follows: P. berghei AL 22 h, 111; CR 22 h, 78; AL 30 h, 74; CR 30 h, 94; P. yoelii AL, 58; CR, 107. d, Flow cytometry analysis of P. berghei ANKA schizonts prepared as in b and stained with SYBR Green to quantify the DNA content (two independent experiments). e, Representative flow cytometry plots and gating strategy for analysis of GFP-expressing P. berghei ANKA parasites after 24 h in vitro culture with 25% AL or CR rat sera (glucose, 4 mM). Cells were selected on FSC and SSC and then on FITC (green) and PE (red) channels. As shown in left and middle panels, mature schizonts express strong GFP signal detected in the FITC channel. Histogram plot show fluorescent intensity comparison between AL and CR. Data represents 1 of 3 independent experiments. f, Luminescence analysis of schizont-specific luciferase-expressing parasites after in vitro maturation (mean ± s.e.m.; n = 5; Mann–Whitney). g, Comparing parasitaemia levels (left axis, black) and the estimated parasite numbers (right axis, blue) using a geometric progression (y = mx) in which the basis is the mean merozoite number for AL and CR (9 and 6, respectively) for the first days of parasite linear growth. Parasitaemia data was obtained from Fig. 2c (mean ± s.e.m.; n = 11 per group; AL, closed circles, CR open circles). Mathematical modelling of parasitaemia taking into account only the number of merozoites appears to be sufficient to predict the observed growth difference of P. berghei during early infection. h, i, Box plot of merozoite numbers (h) and parasitaemia (i) of wild-type P. berghei ANKA and Δpk7 parasites. Number of schizonts analysed after in vitro maturation in AL conditions in 2 independent experiments are as follows: wild type, 36; Δpk7, 85. Parasitaemia determined by microscopic examination of blood smears from C57BL/6 mice infected with 1 × 106 iRBCs of P. berghei ANKA Δpk7 (n = 4) and the parental wild type (AL n = 3; CR n = 4). The data show that parasites producing fewer merozoites lead to similar low parasitaemia to that of parasites under CR.
Extended Data Figure 3 Microarray analysis of P. berghei parasites under CR.
a, Schematic representation of parasite sample preparation for microarray time-course analysis. P. berghei ANKA parasites collected with 4 h intervals 30 h after intravenous injection of purified schizonts into AL and CR mice. b, Microscopy analysis of parasite size (as proxy for parasite age) in the samples used in the microarrays show no apparent morphological differences in parasite development under AL and CR across the different time-points. The parasite area (a.u., arbitrary units) is defined by the Giemsa staining on thin blood smears33 and was scored using ImageJ. Histograms of parasite size distribution (3 mice per group per time point). The total number of parasites analysed are as follows: 6 h, AL n = 150, CR n = 143; 10 h, AL n = 158, CR n = 159; 14 h, AL n = 158, CR n = 157; 18 h, AL n = 180, CR n = 121). The indicated time-points correspond to the parasite developing time after RBC re-invasion, in the second cycle. c. Scatter plots of log2 fold change (y axis) and mean expression levels (x axis) of parasites in CR versus AL (CR/AL) of 3 mice per group at the indicated time-points. Genes differentially induced or repressed in CR (with log2(fold change) of 1 and FDR adjusted P < 0.01) are highlighted in red and blue, respectively. The number (and relative percentage) of genes altered for each time-point is given in the graphs in red (induced) and blue (repressed). d, Correlation plot between microarray and qPCR analysis for the 14-h samples. 20 genes were selected on the basis of the highest fold-changes, analysed by qPCR and compared to the values obtained in the microarray analysis. Validation rate was 80%. Values are mean of 3 mice per group. The list of genes analysed and their fold-changes in qPCR and microarray are provided in the Source Data. e, qPCR analysis of repressed genes in independent biological samples collected at 14 h (AL n = 4, CR n = 3). Each circle represents one mouse. Gene IDs are shown without the ‘PBANKA_’ prefix. Microarray hybridization was performed one time and confirmed by qPCR for a subset of genes in the same RNA samples (d), as well as independently collected samples (e). f, Gene ontology enrichment analysis (Molecular Function) of the genes showing significant alterations for each time point using PlasmoDB (http://www.plasmodb.org) tools and considering Benjamini–Hochberg adjusted P <0.05. The graph highlights the top four of terms with highest significance for each time point and/or terms that appear more than once. Red, induced; blue, repressed. The full list of terms (including biological process analysis) for each time point are provided in the Source Data.
Extended Data Figure 4 Screen of P. berghei kinase mutants and characterization of Δkin and complemented parasite lines.
a, Screening of kinase mutants using the CR in vitro assay. Screen performed in media supplemented with 25% of AL or CR sera (glucose, 4 mM). The graph shows the relative reduction of merozoite formation in CR (unfilled bars) in comparison to AL (filled bars). Values are mean ± s.d. of 3 independent experiments for wild type, Δnek4, Δpk7, Δkin, Δcdkl, Δcdpk3, Δgak, Δtkl5 and 2 independent experiments for other knockout lines. The total number of schizonts analysed is as follows: wild type, AL n = 150, CR n = 167; Δkin, AL n = 80, CR n = 86; Δpk1, AL n = 108, CR n = 89; Δpk7, AL n = 134, CR n = 154; Δnek2, AL n = 99, CR n = 86; Δnek4, AL n = 117, CR n = 79; Δcdpk3, AL n = 46, CR n = 81; Δcdpk4, AL n = 55, CR n = 63; Δgak, AL n = 46, CR n = 54; Δcdkl, AL n = 79, CR n = 62; Δtkl5, AL n = 99, CR n = 93; Δsrpk, AL n = 59, CR n = 86; Δeik1, AL n = 110, CR n = 71; Δeik2, AL n = 116, CR n = 84; Δmap1, AL n = 90, CR n = 59; Δmap2, AL n = 50, CR n = 42. b, Box plot of microscopic analysis of merozoite numbers for P. berghei ANKA wild-type (AL n = 49, CR n = 77) and a second independent clone of Δkin (AL n = 54, CR n = 73) in the same conditions as in a (Mann–Whitney test). c, Schematic of Δkin complementation strategy. Double crossover recombination at kin 5′ and 3′ UTR was used to genetically delete the previously introduced Toxoplasma gondii dhfr (tgdhfr) and complement with codon-altered kin gene and human dhfr. Transgenic parasites were selected by WR99210 treatment of mice (4 subcutaneous daily injections, 16 mg per kg per day). Annealing sites for genotyping primers are illustrated (left) and primer sequences are given in Supplementary Table 1. Agarose gel image (representative of 3) showing diagnostic PCR products from Δkin and Δkin+kin extracted genomic DNA, after dilution cloning of the complemented parasite line (right). d, Flow cytometry analysis of GFP-expressing P. berghei ANKA wild type, Δkin and complemented Δkin parasites after in vitro maturation to schizonts with medium supplemented with AL and CR sera as in a and analysed as in Extended Data Fig. 2e. Histograms represent 2 independent experiments. e, Full course parasitaemia (mean ± s.e.m.) and survival of C57BL/6 mice AL and CR infected by i.p. injection of 1 × 106 iRBCs of P. berghei wild type (AL n = 7, CR n = 7), Δkin (AL n = 9, CR n = 8) and Δkin+kin (AL n = 10, CR n = 6). AL, closed circles; CR, open circles. f, Analysis of parasite area (arbitrary units, a.u.) on Giemsa-stained smears of the samples used for RNA sequencing, as in Extended Data Fig. 3b. Histograms of parasite size distribution (3 mice per group). The total number of parasites analysed as follows: wild type, AL n = 172, CR n = 148; Δkin, AL n = 112, CR n = 129. g, Correlation plot between microarray and RNA-seq analysis for the wild-type samples at 10 h. Analysis of top 500 genes with P < 0.01 in CR versus AL and expression levels higher than the first quartile in both platforms are shown in the graph. Despite the use of different platforms to analyse gene expression, there is 0.45 correlation between microarray and RNA-seq data from the two independently obtained wild-type samples (P < 0.001). h, Comparison of GO term enrichment analysis of CR-altered genes between RNA-seq and microarray platforms for the 10 h time-point. The GO ‘Molecular Function’ graph highlights the location and relation of significantly enriched terms. As indicated in the key, node size refers to the level of significance of each GO term, while the colour of the node represents if a particular term was detected in one or both platforms. The graph is split into two halves; where the top half represents the enrichment of terms from upregulated genes and the bottom half that of downregulated genes. Although the overlap between different transcriptomic methods was, as expected incomplete, this GO term enrichment analysis of the data sets revealed consistency in the functions of the genes that responded to CR. i, qPCR analysis of wild-type P. berghei and Δkin in independent biological samples (wild type, n = 3 per group; Δkin, n = 5 per group). Data normalized to AL of the correspondent genotype. Each circle represents 1 mouse. Gene IDs are given without the ‘PBANKA_’ prefix. The genes analysed were experimentally validated in Extended Data Fig. 3d, e and encode proteins related to lipid metabolism, members of transcriptional regulators (ApiAP2), and several transporters.
Extended Data Figure 5 KIN and KIN(T616D) phosphomimetic mutation.
a, Schematic diagram of yeast SNF1 and human AMPKα showing the conservation of P. falciparum and P. berghei KIN kinase catalytic domain (red). This kinase domain is flanked by an unusually long N-terminal region and a poorly conserved C terminus, both with no obvious domains. Amino acid sequence alignment of the activation loop reveals a high degree of similarity and the conservation of the T-loop threonine (red line) whose phosphorylation is essential for kinase activity. AID, autoinhibitory domain; KA1, kinase associated domain 1. b, Model of KIN on AMPKα (top) and KIN catalytic domain (bottom). The predicted amino acid sequence of KIN kinase domain (455–712) was used to generate a model by Phyre (http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index) using the human AMPK structure (PDB: 4RER). The model is shown in cartoon representation and depicts the small lobe (455–539) in green, the large lobe (540–712) in blue and the T-loop (601-626) in red with the T616 in sticks. An ATP molecule (stick representation) was docked to illustrate the catalytic site with Lysine 489 (stick representation). Both the AID and the KA1 domain were also modelled but it appeared not to be conserved. Figures were generated with PyMol (http://www.pymol.org). c, Schematic for the generation of P. berghei KIN and KIN(T616D) mutant. Double crossover recombination at 5′ and 3′ UTR was used to genetically delete endogenous kin and complement with codon-altered kin gene encoding wild-type (KIN(T616)) or phosphomimetic mutation (KIN(T616D)). Codon-altered sequence of kin was obtained from GenScript, which was then used in the site-directed mutagenesis reaction to introduce the T616D mutation. Presence of codon-altered kin and kinT616D was confirmed by sequencing of the locus of the transgenic parasites. Annealing sites for genotyping primers are illustrated and primer sequences are given in Supplementary Table 1. d, Agarose gel image (representative of 2) showing diagnostic PCR products from P. berghei KIN(T616), KIN(T616D) and parental wild-type. e, Survival of C57BL/6 mice infected by i.p. injection (1 × 106 iRBCs per mouse) of P. berghei KIN(T616) (n = 9) and KIN(T616D) (n = 9). f, Complementation of the Δsnf1 yeast mutant with GFP-fused yeast-optimized sequences of N-terminal truncated P. berghei kin, kinT616D or yeast snf1 without GFP tag. Truncation is indicated with ‘t.’. Growth of transformed Δsnf1 cells in culture, inoculated at a density of 0.05 OD600 in s.d. medium supplemented with glucose or raffinose as a carbon source and grown for 42 h. Shown are OD values obtained in the raffinose condition, normalized to those obtained in glucose condition for each cell line (snf1 n = 6, t.kin, n = 6, t.kinT616D n = 6; mean ± s.e.m.; Mann–Whitney test). g, Western blot analysis of Δsnf1 expressing full length (left) or N-terminal truncated (right) KIN(T616)–GFP or KIN(T616D)–GFP. Predicted size of full-length KIN–GFP is 147 kDa and of N-terminal truncated version is 93 kDa. Membranes were probed with anti-GFP antibody. A representative blot from 2 independent lysates is shown.
Extended Data Figure 6 Effect of AMPK agonists.
a, Dose-dependent effect of AMPK activators (salicylate and A769662) on P. berghei ANKA that express luciferase (under ama1 schizont-specific promoter). Parasites were cultured for 24 h with increasing concentrations of the compounds (media supplemented with 20% FBS and 5 mM glucose). Analysis of schizont development was performed by measuring luminescence. Values are mean ± s.e.m. (salicylate, n = 5; A769662, n = 6). EC50 values determined by using GraphPad Prism nonlinear regression variable slope (normalized) analysis. The calculated EC50 values are as follows: salicylate, 2.4 ± 0.9 mM; A769662, 256.5 ± 60.6 μM. b, Dose-dependent effect of A769662 on P. falciparum, P. berghei ANKA wild-type, Δkin and complemented parasites analysed by microscopy after Giemsa staining (Mann–Whitney test). Box plots show the data for the following number of schizonts (vehicle, A769662 62.5 and 125 μM): P. falciparum, 59, 18, 48; wild-type P. berghei, 28, 68, 61; Δkin, 44, 47, 40; Δkin+kin, 34, 57, 37. c, Dose-dependent effect of A769662 and salicylate treatments on P. falciparum for two developmental cycles. Synchronized cultures were set at 0.1% initial parasitaemia (rings) and analysed at 48 h and 96 h by flow cytometry after SYBR Green labelling of parasite DNA. A new generation of rings was observed at 48 and 96 h in the treated conditions, suggesting no growth delay. Data (mean ± s.e.m.) was normalized to the untreated control on each experiment at 48 h or 96 h. The EC50 values (determined as in a) are as follows: 133.1 ± 3.4 μM at 48 h (n = 6) and 70.1 ± 18.9 μM at 96 h (n = 5) for A769662; 2.2 ± 0.2 mM at 48 h (n = 6) and 1.25 ± 0.2 mM at 96 h (n = 6) for salicylate. d, qPCR analysis of P. falciparum parasites treated with salicylate for 72 h (n = 2 per condition). Data normalized to the untreated control. The genes analysed correspond to the P. berghei homologues experimentally validated in Extended Data Fig. 3. Gene IDs are shown in the figure without the ‘PF3D7_’ prefix. e, Dose-dependent effect of salicylate on other P. berghei ANKA kinase mutants. Box plot of parasites treated and analysed as in b (Mann–Whitney test). The number of schizonts analysed for vehicle, 0.6 mM and 1.25 mM are as follows: Δnek2, 43, 33, 37; Δcdpk3, 35, 44, 30. f, g, Salicylate effect in vivo. Mice were treated daily with 250 mg kg−1 salicylate (sal) or 0.9% NaCl (vehicle, veh) starting at day 1 after infection. Parasitaemia (mean ± s.e.m.; two-way ANOVA test) and survival (log-rank Mantel–Cox test) of C57BL/6 mice infected by i.p. injection of 1 × 106 wild-type (veh n = 15; sal n = 16), Δkin (veh n = 8; sal n = 8) and complemented Δkin+kin iRBCs (veh n = 10; sal n = 10).
Extended Data Figure 7 Supplementation studies in vitro and in vivo.
a, Effect of supplementation with extra glucose, vitamins, essential amino acids (EAA), non-essential (NEAA), leptin and iron (FeSO4) on salicylate- or A769662-treated parasites. P. berghei expressing luciferase at schizont stage were cultured in 20% FBS supplemented medium (5 mM glucose, n = 5) with increasing concentrations of salicylate and extra glucose (50 mM, n = 5), vitamins (1×, n = 3; 5×, n = 2), EAA (100 μM, n = 3; 500 μM, n = 2), NEAA (100 μM, n = 3; 500 μM, n = 2), and leptin (50 ng ml−1, n = 2; 150 ng ml−1 n = 2). FeSO4 was added in the presence of equal amount of ascorbic acid (AA), an iron absorption enhancer. A769662 treatments were conducted in 20% FBS supplemented medium (5 mM glucose) and leptin (50 ng ml−1, n = 2; 150 ng ml−1 n = 2). Analysis of schizont development was performed by measuring luciferase activity as in Extended Data Fig. 2f. Values normalized to the vehicle control. Replicates are shown as individual data points for all supplementations except 50 mM glucose, AA and salicylate alone, which are shown as mean ± s.e.m. Salicylate EC50 values in these experiments were 3.4 ± 0.3 mM and 5.7 ± 1.3 mM for 5 and 50 mM glucose, respectively (P = 0.0081, two-way ANOVA test). b, Box plot of microscopic analysis of P. berghei ANKA wild-type segmented schizonts obtained in vitro after 24 h maturation in the presence of AL or CR sera. Recombinant leptin was added to the culture medium at the indicated concentration. Total number of schizonts analysed in two independent experiments as follows: AL, 87; CR, 85; AL+Leptin, 69; CR, 86 (Mann–Whitney test). c, Dose-dependent effect of salicylate and A769662 on P. berghei parasites in the presence of AL or CR sera. Parasites were incubated for 24 h with increasing concentrations of the compounds in media supplemented with 10% of AL or CR sera (glucose 5 mM). Analysis of schizont development was performed by measuring luminescence (A769662, n = 3; salicylate, n = 5). These compounds appear to have additive effects, suggesting that under CR conditions, A769662/salicylate might activate KIN through distinct and/or complementary mechanisms. This could be related to different binding sites on the kinase, as previously demonstrated for other AMPKs23,24. d, Effect of glucose supplementation on salicylate-treated P. falciparum parasites. Ring-stage synchronized cultures were set at 0.1% initial parasitaemia in a glucose free medium supplemented with 5 and 25 mM glucose. Values in the graph are parasitaemia (mean ± s.e.m.; n = 5 per condition) determined by flow cytometry after staining with SYBR Green at 48 h. Salicylate EC50 values were 2.0 ± 0.1 mM and 2.7 ± 0.2 mM for 5 and 25 mM glucose, respectively. P value in the figure calculated with two-way ANOVA test. e–g, Effect of glucose supplementation on C57BL/6 male mice in CR or AL. 0.2 g ml−1 of glucose was provided in the drinking water starting on the day of infection. e, Daily food consumption and water intake for the same group of mice. Blood glucose levels after 20 h of glucose supplementation. Representative of two independent experiments (AL n = 5, CR n = 4, AL+glucose n = 5, CR+glucose n = 5). f, Parasitaemia of C57BL/6 mice AL or CR and glucose-supplemented groups, infected by i.p. injection of 1 × 106 iRBCs of GFP-expressing P. berghei wild-type parasites. Data represent 1 of 2 independent experiments (n = 3 per group). Each mouse is plotted as an individual data point. g, qPCR analysis of P. berghei wild-type parasites in AL- or CR-fed C57BL/6 mice supplemented (diamonds; n = 5) or not (circles; n = 4) with glucose in drinking water. Shown is parasite relative gene expression in CR normalized to the mean of the correspondent AL condition at day 5 after i.p. infection. Each mouse is plotted as an individual data point. Representative of two experiments performed independently. Gene IDs are given without the ‘PBANKA_’ prefix as in Extended Data Fig. 3e.
Extended Data Figure 8 Schematic representation of the observed effect of dietary nutrients or AMPK agonists on Plasmodium intraerythrocytic replication.
The data supports the idea that parasites can replicate in higher or fewer numbers depending on host nutrient availability. This active parasite response mediated by an AMPKα-related kinase, KIN, which is expected to become active by an increase of the AMP/ATP ratio in parasites facing nutrient deficiency. KIN upstream regulators and downstream targets remain to be determined, as well as other potential molecular factors that might also contribute to this nutrient sensing mechanism.
Supplementary information
Supplementary information
This file contains the uncropped source data for Figure 2g, Extended Data Figure 4c, and Extended Data Figures 5d and g and Supplementary Table 1. (PDF 1067 kb)
Source data
Rights and permissions
About this article

Cite this article
Mancio-Silva, L., Slavic, K., Grilo Ruivo, M. et al. Nutrient sensing modulates malaria parasite virulence. Nature 547, 213–216 (2017). https://doi.org/10.1038/nature23009
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature23009
This article is cited by
-
Plasmodium yoelii iron transporter PyDMT1 interacts with host ferritin and is required in full activity for malarial pathogenesis
BMC Biology (2023)
-
A non-canonical sensing pathway mediates Plasmodium adaptation to amino acid deficiency
Communications Biology (2023)
-
Rapid metabolic reprogramming mediated by the AMP-activated protein kinase during the lytic cycle of Toxoplasma gondii
Nature Communications (2023)
-
Cytoprotective autophagy as a pro-survival strategy in ART-resistant malaria parasites
Cell Death Discovery (2023)
-
Stochastic expression of invasion genes in Plasmodium falciparum schizonts
Nature Communications (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
