
Abstract
The palaeobiological record of 12 million to 7 million years ago (Ma) is crucial to the elucidation of African ape and human origins, but few fossil assemblages of this period have been reported from sub-Saharan Africa. Since the 1970s, the Chorora Formation, Ethiopia, has been widely considered to contain ~10.5 million year (Myr) old mammalian fossils1,2,3,4,5,6,7. More recently, Chororapithecus abyssinicus, a probable primitive member of the gorilla clade6, was discovered from the formation. Here we report new field observations and geochemical, magnetostratigraphic and radioisotopic results that securely place the Chorora Formation sediments to between ~9 and ~7 Ma. The C. abyssinicus fossils are ~8.0 Myr old, forming a revised age constraint of the human–gorilla split. Other Chorora fossils range in age from ~8.5 to 7 Ma and comprise the first sub-Saharan mammalian assemblage that spans this period. These fossils suggest indigenous African evolution of multiple mammalian lineages/groups between 10 and 7 Ma, including a possible ancestral-descendent relationship between the ~9.8 Myr old Nakalipithecus nakayamai8 and C. abyssinicus. The new chronology and fossils suggest that faunal provinciality between eastern Africa and Eurasia had intensified by ~9 Ma, with decreased faunal interchange thereafter9,10,11,12. The Chorora evidence supports the hypothesis of in situ African evolution of the Gorilla–Pan–human clade, and is concordant with the deeper divergence estimates of humans and great apes based on lower mutation rates of ~0.5 × 10−9 per site per year (refs 13, 14, 15).
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others

References
Sickenberg, O. & Schönfeld, M. in Afar Depression of Ethiopia (eds Pilger, A., Rössler, A. ) 277–284 (Schweizerbart, 1975)
Tiercelin, J. J., Michaux, J. & Bandet, Y. Le Miocène supérieur du sud de la dépression de l’Afar, Éthiopie; sédiments, faunes, âges isotopiques. Bull. Soc. Geol. Fr. Ser. 7 XXI, 255–258 (1979)
Geraads, D., Alemseged, Z. & Bellon, H. The late Miocene mammalian fauna of Chorora, Awash basin, Ethiopia: systematics, biochronology and the 40K-40Ar ages of the associated volcanics. Tertiary Res . 21, 113–127 (2002)
Bernor, R. L., Kaiser, T. M. & Nelson, S. V. The oldest Ethiopian hipparion (Equinae, Perissodactyla) from Chorora: systematics, paleodiet and paleoclimate. Cour. Forsch. Senckenberg 246, 213–226 (2004)
Cote, S. M. Origins of the African hominoids: an assessment of the palaeobiogeographical evidence. C. R. Palevol 3, 323–340 (2004)
Suwa, G., Kono, R. T., Katoh, S., Asfaw, B. & Beyene, Y. A new species of great ape from the late Miocene epoch in Ethiopia. Nature 448, 921–924 (2007)
Werdelin, L. & Sanders, W. J. Cenozoic Mammals of Africa (Univ. California Press, 2010)
Kunimatsu, Y. et al. A new Late Miocene great ape from Kenya and its implications for the origins of African great apes and humans. Proc. Natl Acad. Sci. USA 104, 19220–19225 (2007)
Douady, C. J., Catzeflis, F., Raman, J., Springer, M. S. & Stanhope, M. J. The Sahara as a vicariant agent, and the role of Miocene climatic events, in the diversification of the mammalian order Macroscelidea (elephant shrews). Proc. Natl Acad. Sci. USA 100, 8325–8330 (2003)
Boisserie, J.-R., Fisher, R. E., Lihoreau, F. & Weston, E. M. Evolving between land and water: key questions on the emergence and history of the Hippopotamidae (Hippopotamoidea, Cetancodonta, Cetartiodactyla). Biol. Rev. Camb. Phil. Soc . 86, 601–625 (2011)
Bibi, F. Mio-pliocene faunal exchanges and African biogeography: the record of fossil bovids. PLoS ONE 6, e16688 (2011)
Zhang, Z. et al. Aridification of the Sahara desert caused by Tethys Sea shrinkage during the Late Miocene. Nature 513, 401–404 (2014)
Scally, A. et al. Insights into hominid evolution from the gorilla genome sequence. Nature 483, 169–175 (2012)
Hara, Y., Imanishi, T. & Satta, Y. Reconstructing the demographic history of the human lineage using whole-genome sequences from human and three great apes. Genome Biol. Evol. 4, 1133–1145 (2012)
Langergraber, K. E. et al. Generation times in wild chimpanzees and gorillas suggest earlier divergence times in great ape and human evolution. Proc. Natl Acad. Sci. USA 109, 15716–15721 (2012)
Tsujikawa, H. T he updated late Miocene large mammal fauna from Samburu Hills, northern Kenya. Afr. Study Monogr . 32 (suppl.), 1–50 (2005)
Leakey, M. G. & Harris, J. M. Lothagam: The Dawn of Humanity in Eastern Africa. (Columbia Univ. Press, 2003)
Boisserie, J.-R. et al. A new species of Nyanzachoerus (Cetartiodactyla: Suidae) from the late Miocene Toros-Ménalla, Chad, central Africa. PLoS ONE 9, e103221 (2014)
Chernet, T., Hart, W., Aronson, J. & Walter, R. C. New age constraints on the timing of volcanism and tectonism in the northern Main Ethiopian Rift–southern Afar transition zone (Ethiopia). J. Volcanol. Geotherm. Res. 80, 267–280 (1998)
Wolfenden, E., Ebinger, C., Yirgu, G., Deino, A. & Ayalew, D. Evolution of the northern Main Ethiopian rift: birth of a triple junction. Earth Planet. Sci. Lett. 224, 213–228 (2004)
Kunz, K., Kreuzer, H. & Müller, P. in Afar Depression of Ethiopia (eds Pilger, A. & Rössler, A. ) 370–374 (Schweizerbart, 1975)
Suwa, G. et al. Newly discovered 7 to 9 million-year-old cercopithecid, equid and other mammalian fossils from the Chorora Formation, Ethiopia. Anthropol. Sci. 123, 19–39 (2015)
Benefit, B. R. & Pickford, M. Miocene fossil cercopithecoids from Kenya. Am. J. Phys. Anthropol. 69, 441–464 (1986)
Gilbert, C. C., Goble, E. D. & Hill, A. Miocene cercopithecoidea from the Tugen Hills, Kenya. J. Hum. Evol. 59, 465–483 (2010)
Bernor, R. L., Armour-Chelu, M. J., Gilbert, H., Kaiser, T. M. & Schulz, E. in: Cenozoic Mammals of Africa (eds Werdelin, L. & Sanders, W. J. ) 685–721 (Univ. California Press, 2010)
Uno, K. T. et al. Late Miocene to Pliocene carbon isotope record of differential diet change among East African herbivores. Proc. Natl Acad. Sci. USA 108, 6509–6514 (2011)
Eronen, J. T. et al. Distribution history and climatic controls of the Late Miocene Pikermian chronofauna. Proc. Natl Acad. Sci. USA 106, 11867–11871 (2009)
Flynn, L. J. et al. The Leporid Datum: a late Miocene biotic marker. Mammal Rev. 44, 164–176 (2013)
Hobolth, A., Dutheil, J. Y., Hawks, J., Schierup, M. H. & Mailund, T. Incomplete lineage sorting patterns among human, chimpanzee, and orangutan suggest recent orangutan speciation and widespread selection. Genome Res. 21, 349–356 (2011)
Begun, D. R., Nargolwalla, M. C. & Kordos, L. European Miocene hominids and the origin of the African ape and human clade. Evol. Anthropol. 21, 10–23 (2012)
Tiercelin, J. J. & Michaux, J. Un example de remplissage mio-pliocène de rift dans l’Est africain: la Formation de Ch’orora, Éthiopie: stratigraphie et mammifères fossils. In 5e Réun. ann. Sc. Terre, Renne, 449 (1977)
Katoh, S. et al. Chronostratigraphy and correlation of the Plio-Pleistocene tephra layers of the Konso Formation, southern main Ethiopia rift, Ethiopia. Quat. Sci. Rev. 19, 1305–1317 (2000)
WoldeGabriel, G., Hart, W. K., Katoh, S., Beyene, Y. & Suwa, G. Correlation of Plio-Pleistocene tephra in Ethiopian and Kenyan rift basins: temporal calibration of geological features and hominid fossil records. J. Volcanol. Geotherm. Res. 147, 81–108 (2005)
Borchardt, G. A., Harward, M. E. & Schmitt, R. A. Correlation of volcanic ash deposits by activation analysis of glass separates. Quat. Res. 1, 247–260 (1971)
Katoh, S., Danhara, T., Hart, W. K. & WoldeGabriel, G. Use of sodium polytungstate solution in the purification of volcanic glass shards for bulk chemical analysis. Nat. Hum. Activities 4, 45–54 (1999)
Itaya, T. et al. Argon analysis by a newly-developed mass spectrometric system for K-Ar dating. Mineral. J. 15, 203–221 (1991)
Ryu, S., Oka, M., Yagi, K. & Itaya, T. K-Ar ages of the Quaternary basalts in the Jeongok area, the central part of Korean peninsula. GeoJournal 15, 1–8 (2011)
Hyodo, H., Kim, S.-W., Itaya, T. & Matsuda, T. Homogeneity of neutron flux during irradiation for 40Ar/39Ar age dating in the research reactor at Kyoto University. J. Min. Petr. Econ. Geol. 94, 329–337 (1999)
Kawamura, M., Uchino, T., Gouzu, C. & Hyodo, H. 380 Ma 40Ar/39Ar ages of the high-P/T schists obtained from the Nedamo Terrane, Northeast Japan. J. Geol. Soc. Japan 113, 492–499 (2007)
Hyodo, M. et al. High-resolution record of the Matuyama-Brunhes transition constrains the age of Javanese Homo erectus in the Sangiran dome, Indonesia. Proc. Natl Acad. Sci. USA 108, 19563–19568 (2011)
Kirschvink, J. L. The least-squares line and plane and the analysis of palaeomagnetic data. Geophys. J. R. Astron. Soc . 62, 699–718 (1980)
Cande, S. C. & Kent, D. V. Revised calibration of the geomagnetic polarity time scale for the Late Cretaceous and Cenozoic. J. Geophys. Res. 100, 6093–6095 (1995)
White, T. D. et al. Macrovertebrate paleontology and the Pliocene habitat of Ardipithecus ramidus. Science 326, 67, 87–93 (2009)
Kohn, M. J. Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proc. Natl Acad. Sci. USA 107, 19691–19695 (2010)
Tipple, B. J., Meyers, S. R. & Pagani, M. Carbon isotope ratio of Cenozoic CO2: a comparative evaluation of available geochemical proxies. Paleoceanography 25, PA3202 (2010)
Feakins, J. J. et al. Northeast African vegetation change over 12 m.y. Geology 41, 295–298 (2013)
Diefendorf, A. F., Mueller, K. E., Wing, S. L., Koch, P. L. & Freeman, K. H. Global patterns in leaf 13C discrimination and implications for studies of past and future climate. Proc. Natl Acad. Sci. USA 107, 5738–5743 (2010)
Cerling, T. E., Harris, J. M., Leakey, M. G. & Mudida, N. in Lothagam: The Dawn of Humanity in Eastern Africa (eds Leakey, M. G., Harris, J. M. ) 584–603 (Columbia Univ. Press, 2003)
Boisserie, J.-R. et al. Diets of modern and late Miocene hippopotamids: evidence from carbon isotope composition and micro-wear of tooth enamel. Palaeogeogr. Palaeoclimatol. Palaeoecol. 221, 153–174 (2005)
Acknowledgements
We thank the Authority for Research and Conservation of Cultural Heritage, Ministry of Culture and Tourism of Ethiopia, for permissions and facilitation; we thank the Western Hararge Chiro Zone Culture and Tourism Office and the Mieso Woreda for fieldwork support; we thank all participants in the fieldwork, especially the Gololcha people, who were essential to the success of the project. Neutron irradiation for 40Ar–39Ar dating was done under the Visiting Researchers Program at the Kyoto University Research Reactor Institute. We thank J. Morton for assistance with major and trace element analyses, and the Janet and Elliott Banes Professorship and National Science Foundation EAR-1028789 for support to W.H. We thank H. Ishiguro for assistance with sampling enamel for the isotope analysis, and M. Nakatsukasa and Y. Kunimatsu for comparative materials. We thank D. Geraads for providing field information about previous geochronological sampling localities. This project was supported primarily by the Japan Society for the Promotion of Science (Kakenhi grant numbers 21255005 and 24000015).
Author information
Authors and Affiliations
Contributions
G.S., Y.B., B.A., S.K. designed the research; G.S., Y.B., B.A., S.K., T.S., K.S. conducted field work; S.K., T.I., H.H., M.H., K.Y., C.G., G.W, W.H. did the geochronological and geochemical analysis; G.S., H.N., R.B., J.-R.B., F.B., H.S., S.A. did the faunal and isotopic analysis; and G.S., Y.B., B.A., S.K. wrote the manuscript with contributions from all co-authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Additional information
The Chorora Formation vertebrate fossils have been deposited in the Paleontology and Paleoanthropology laboratories of the Authority for Research and Conservation of Cultural Heritage, Ministry of Culture and Tourism, Addis Ababa, Ethiopia.
Extended data figures and tables
Extended Data Figure 1 Cladistic relationships of C. abyssinicus and N. nakayamai.
C. abyssinicus is known from only nine teeth (or fragments)6 and N. nakayamai from a mandibular corpus with M1–M3, and ten other isolated teeth (excluding a possible antimere)8. The only informative dental elements shared by the two reported samples are the lower M3 and the lower M1 (the latter damaged in C. abyssinicus and considerably worn in N. nakayamai). This fragmentary evidence makes cladistic evaluations difficult. C. abyssinicus was considered to share the following derived combination of features with the modern gorilla: large postcanine size, upper molars buccolingually narrow and mesiodistally elongate, with a relatively long and mesiobuccally extending mesial protocone crest, reduced protoconule, and lower molars with a prominent anterior transverse crest. Following ref. 6, we consider Chororapithecus to be a basal member of the gorilla clade. When compared with the Middle Miocene examples of Kenyapthecini and Equatorini, C. abyssinicus and N. nakayamai share reduced cingula in both upper and lower molars, although ref. 8 pointed out that the lower M3 cingulum appears slightly better developed in N. nakayamai. The available N. nakayamai teeth include elements (not reported in C. abyssinicus) that are morphologically more derived than in Kenyapthecini/Equatorini, such as upper premolars that are relatively elongate mesiodistally (narrow buccolingually) and a lower P3 that is not as obliquely elongate and transversly compressed as in the Middle Miocene forms. On the basis of these observations, ref. 8 preferred a stem modern African ape–human cladistic position of N. nakayamai (option 1). However, N. nakayamai also shares two possibly derived features with C. abyssinicus: large postcanine size (although slightly less so) and prominent transverse crest in the lower molars. The latter is inferred from the characteristic wear pattern of the holotype mandible lower M1, the dentine exposures of the lingual cusps that are buccolingually linear (figure 1 of ref. 8). These observations suggest that N. nakayamai may also be a stem member of the gorilla clade but more primitive than C. abyssinicus (option 2).
Extended Data Figure 2 Chorora Formation type locality.
a, View north-northwest from the southeastern margin of the type locality. Yellow and red arrows show the rhyolitic ignimbrite units considered by previous workers to overlie the Chorora Formation sediments1,2,3. The ignimbrite unit (yellow arrows) adjacent to the sediment exposures has been interpreted to directly overlie the Chorora Formation sediments including the gravel unit that forms the resistant ledge along the western margin of the whitish lacustrine exposures. A second ridge top unit (red arrows) was considered to cap the entire sequence. RhyUn, RhyOv1 and RhyOv2 indicate sampling localities of the present study for K–Ar dating. The RhyUn unit dips westwards and stratigraphically underlies the Chorora Formation sediments. The horizontally bedded light brown sediments are Middle Pleistocene in age and unconformably cap both RhyUn and Chorora Formation sediments. The yellow and red dotted lines indicate approximate positions of the step faults that resulted in uplifted RhyOv1 and RhyOv2 exposures. b, View northeast at the RhyOv1 sampling spot. Yellow arrows point to the RhyOv1 rhyolitic ignimbrite unit forming a planar terrace. c, View west of the b RhyOv1 section (yellow arrows). Note that the rhyolitic ignimbrite unit terminates abruptly (dotted line) in fault contact with sediments previously considered to underlie the ignimbrite2,3.
Extended Data Figure 3 Electron-probe microanalysis (Or–Ab–An diagrams) of the K–Ar-dated samples.
Results of the electron-probe microanalysis of feldspar separates of the mesh size fraction that was used in the K–Ar dating of samples listed in Extended Data Table 1. The full data are shown in Supplementary Table 3.
Extended Data Figure 4 Single crystal step-heating 40Ar–39Ar analysis.
Results of individual crystals with stable ages (indicated by arrows) are shown. Errors are 1σ. Samples 10TC-1 and 10TC-58 are from the same consolidated tuff unit collected <5 m apart. Samples 10TC-52 and 10TC-53 are from the same pumiceous tuff collected ~30 m apart. The weighted means of these are shown in Fig. 3 and Extended Data Table 1.
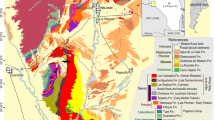
Extended Data Figure 5 Chorora Formation stratigraphic columns of the type locality, Beticha and nearby localities.
The sections were taken at exposures adjacent to or continuous with the sampling locations of the dated volcanic samples (coordinates tabulated in Extended Data Table 1). Radioisotopic dates (K–Ar ages) and samples correspond to those shown in Fig. 3 and Extended Data Table 1. Eight tephra units are considered to occur at multiple localities, and shown as marker tuffs CT-1 to CT-8. The analytical details of the tuffs are presented in Supplementary Tables 1 and 2. CT-6 is identified at Adadi from lithologic, petrographic, glass morphology and refractive index analyses. Results of the remanent magnetism analysis are given in Supplementary Table 4. See Extended Data Fig. 6 for explanation of the lithological codes.
Extended Data Figure 6 Chorora Formation stratigraphic columns of the western localities.
The sections were taken at exposures adjacent to or continuous with the sampling locations of the dated volcanic samples (coordinates tabulated in Extended Data Table 1). Radioisotopic dates (K–Ar ages) and samples correspond to those shown in Fig. 3 and Extended Data Table 1. Tephra unit CT-4 occurs at multiple localities. The analytical details of the tuffs are presented in Supplementary Tables 1 and 2. Analytical results of the remanent magnetism analysis are given in Supplementary Table 4.
Supplementary information
Supplementary Tables
This file contains Supplementary Tables 1-4. (PDF 299 kb)
Rights and permissions
About this article

Cite this article
Katoh, S., Beyene, Y., Itaya, T. et al. New geological and palaeontological age constraint for the gorilla–human lineage split. Nature 530, 215–218 (2016). https://doi.org/10.1038/nature16510
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature16510
This article is cited by
-
Postcranial evidence of late Miocene hominin bipedalism in Chad
Nature (2022)
-
Divergence-time estimates for hominins provide insight into encephalization and body mass trends in human evolution
Nature Ecology & Evolution (2021)
-
Enamel proteome shows that Gigantopithecus was an early diverging pongine
Nature (2019)
-
The rise and fall of the Old World savannah fauna and the origins of the African savannah biome
Nature Ecology & Evolution (2018)
-
From four hands to two feet: human evolution in the context of primate evolution
Primates (2016)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
