
Abstract
High carbon dioxide (CO2) concentrations in sea-water (ocean hypercapnia) can induce neurological, physiological and behavioural deficiencies in marine animals1,2,3,4,5,6,7,8,9,10. Prediction of the onset and evolution of hypercapnia in the ocean requires a good understanding of annual variations in oceanic CO2 concentration, but there is a lack of relevant global observational data. Here we identify global ocean patterns of monthly variability in carbon concentration using observations that allow us to examine the evolution of surface-ocean CO2 levels over the entire annual cycle under increasing atmospheric CO2 concentrations. We predict that the present-day amplitude of the natural oscillations in oceanic CO2 concentration will be amplified by up to tenfold in some regions by 2100, if atmospheric CO2 concentrations continue to rise throughout this century (according to the RCP8.5 scenario of the Intergovernmental Panel on Climate Change)11. The findings from our data are broadly consistent with projections from Earth system climate models12,13,14,15. Our predicted amplification of the annual CO2 cycle displays distinct global patterns that may expose major fisheries in the Southern, Pacific and North Atlantic oceans to hypercapnia many decades earlier than is expected from average atmospheric CO2 concentrations. We suggest that these ocean ‘CO2 hotspots’ evolve as a combination of the strong seasonal dynamics of CO2 concentration and the long-term effective storage of anthropogenic CO2 in the oceans that lowers the buffer capacity in these regions, causing a nonlinear amplification of CO2 concentration over the annual cycle. The onset of ocean hypercapnia (when the partial pressure of CO2 in sea-water exceeds 1,000 micro-atmospheres) is forecast for atmospheric CO2 concentrations that exceed 650 parts per million, with hypercapnia expected in up to half the surface ocean by 2100, assuming a high-emissions scenario (RCP8.5)11. Such extensive ocean hypercapnia has detrimental implications for fisheries during the twenty-first century.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
References
Cripps, I. L., Munday, P. L. & McCormick, M. I. Ocean acidification affects prey detection by a predatory reef fish. PLoS ONE 6, e22736 (2011)
Devine, B. M. & Munday, P. L. Habitat preferences of coral-associated fishes are altered by short-term exposure to elevated CO2 . Mar. Biol. 160, 1955–1962 (2013)
Devine, B. M., Munday, P. L. & Jones, G. P. Homing ability of adult cardinalfish is affected by elevated carbon dioxide. Oecologia 168, 269–276 (2012)
Dixson, D. L., Munday, P. L. & Jones, G. P. Ocean acidification disrupts the innate ability of fish to detect predator olfactory cues. Ecol. Lett. 13, 68–75 (2010)
Ferrari, M. C. O. et al. Putting prey and predator into the CO2 equation – qualitative and quantitative effects of ocean acidification on predator–prey interactions. Ecol. Lett. 14, 1143–1148 (2011)
Jutfelt, F., De Souza, K. B., Vuylsteke, A. & Sturve, J. Behavioural disturbances in a temperate fish exposed to sustained high-CO2 levels. PLoS ONE 8, e65825 (2013)
Munday, P. L. et al. Ocean acidification impairs olfactory discrimination and homing ability of a marine fish. Proc. Natl Acad. Sci. USA 106, 1848–1852 (2009)
Nilsson, G. E. et al. Near-future carbon dioxide levels alter fish behaviour by interfering with neurotransmitter function. Nature Clim. Change 2, 201–204 (2012)
Spady, B. L., Watson, S. A., Chase, T. J. & Munday, P. L. Projected near-future CO2 levels increase activity and alter defensive behaviours in the tropical squid Idiosepius pygmaeus. Biol. Open 3, 1063–1070 (2014)
Watson, S. A. et al. Marine mollusc predator-escape behaviour altered by near-future carbon dioxide levels. Proc. R. Soc. Lond. B 281, 20132377 (2014)
Meinshausen, M. et al. The RCP greenhouse gas concentrations and their extensions from 1765 to 2300. Clim. Change 109, 213–241 (2011)
Aumont, O. & Bopp, L. Globalizing results from ocean in situ iron fertilization studies. Glob. Biogeochem. Cycles 20, GB2017 (2006)
Dunne, J. P. et al. GFDL’s ESM2 global coupled climate–carbon earth system models. Part II: carbon system formulation and baseline simulation characteristics. J. Clim. 26, 2247–2267 (2013)
Jones, C. D. et al. The HadGEM2-ES implementation of CMIP5 centennial simulations. Geosci. Model Dev. 4, 543–570 (2011)
Yang, D. & Saenko, O. A. Ocean heat transport and its projected change in CanESM2. J. Clim. 25, 8148–8163 (2012)
Sasse, T. P., McNeil, B. I. & Abramowitz, G. A novel method for diagnosing seasonal to inter-annual surface ocean carbon dynamics from bottle data using neural networks. Biogeosciences 10, 4319–4340 (2013)
Egleston, E. S., Sabine, C. L. & Morel, F. M. M. Revelle revisited: buffer factors that quantify the response of ocean chemistry to changes in DIC and alkalinity. Glob. Biogeochem. Cycles 24, GB1002 (2010)
Orr, J. C. et al. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 437, 681–686 (2005)
Sabine, C. L. et al. The oceanic sink for anthropogenic CO2 . Science 305, 367–371 (2004)
Shaw, E. C., McNeil, B. I., Tilbrook, B., Matear, R. & Bates, M. L. Anthropogenic changes to seawater buffer capacity combined with natural reef metabolism induce extreme future coral reef CO2 conditions. Glob. Change Biol. 19, 1632–1641 (2013)
Byrne, M., Lamare, M., Winter, D., Dworjanyn, S. A. & Uthicke, S. The stunting effect of a high CO2 ocean on calcification and development in sea urchin larvae, a synthesis from the tropics to the poles. Phil. Trans. R. Soc. Lond. B 368, 20120439 (2013)
Fabry, V. J., Seibel, B. A., Feely, R. & Orr, J. C. Impacts of ocean acidification on marine fauna and ecosystem processes. ICES J. Mar. Sci. 65, 414–432 (2008)
Maas, A. E., Wishner, K. F. & Seibel, B. A. The metabolic response of pteropods to acidification reflects natural CO2-exposure in oxygen minimum zones. Biogeosciences 9, 747–757 (2012)
Food and Agriculture Organization of the United Nations. The State of World Fisheries and Aquaculture 2014 9–12 (FAO, 2014)
Volff, J.-N. Genome evolution and biodiversity in teleost fish. Heredity 94, 280–294 (2013)
Ishii, M. et al. Air–sea CO2 flux in the Pacific Ocean for the period 1990–2009. Biogeosciences 11, 709–734 (2014)
Lenton, A. et al. Sea–air CO2 fluxes in the Southern Ocean for the period 1990–2009. Biogeosciences 10, 4037–4054 (2013)
Sarma, V. V. S. S. et al. Sea–air CO2 fluxes in the Indian Ocean between 1990 and 2009. Biogeosciences 10, 7035–7052 (2013)
Schuster, U. et al. Trends in North Atlantic sea-surface fCO2 from 1990 to 2006. Deep Sea Res. II 56, 620–629 (2009)
Shaw, E. C., McNeil, B. I. & Tilbrook, B. Impacts of ocean acidification in naturally variable coral reef flat ecosystems. J. Geophys. Res. Oceans 117, C03038 (2012)
Taylor, K. E., Stouffer, R. J. & Meehl, G. A. An overview of CMIP5 and the experiment design. Bull. Am. Meteorol. Soc. 93, 485–498 (2012)
Key, R. M. et al. A global ocean carbon climatology: results from global data analysis project (GLODAP). Glob. Biogeochem. Cycles 18, GB4031 (2004)
Kohonen, T. Self-organizing Maps 3rd edn (Springer, 2001)
Acknowledgements
We thank R. Matear and A. Lenton from the CSIRO for conversations, performing initial model runs and analysis of CMIP5 model output for use here. We acknowledge the World Climate Research Programme’s Working Group on Coupled Modelling, which is responsible for CMIP, and we thank the climate modelling groups (Hadley Centre, IPSL, Can and GFDL) for producing their models and making the output openly accessible. For CMIP, the US Department of Energy’s Program for Climate Model Diagnosis and Intercomparison provides coordinating support, and led the development of software infrastructure in partnership with the Global Organization for Earth System Science Portals. We acknowledge support from the Australian Research Council discovery grant DP110104955.
Author information
Authors and Affiliations
Contributions
B.I.M. conceived and designed the project, interpreted the results and wrote the paper. T.P.S. helped design the project and carried out all of the numerical and data analysis, and contributed important interpretations, discussions and revisions to the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
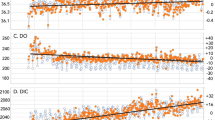
Extended Data Figure 1 Ocean Revelle factor.
Distribution of the mean Revelle or ‘buffer’ factor in the surface ocean in 2000, determined from our study.
Extended Data Figure 2 Quantifying biases in the SOMLO approach.
Bias in  concentrations (expressed as a percentage) in 2100 due to using steady-state assumptions in the SOMLO analysis. Positive (negative) values indicate that SOMLO over-estimates (under-estimates)
concentrations (expressed as a percentage) in 2100 due to using steady-state assumptions in the SOMLO analysis. Positive (negative) values indicate that SOMLO over-estimates (under-estimates)  .
.
Extended Data Figure 3 Future CO2 amplification as predicted by climate models.
a–d, CO2 amplification factors by the year 2100 as predicted by four ESMs: HadGEM2-ES (a), IPSL-CM5A-MR (b), CanESM2 (c) and GFDL-ESM2M (d); see Extended Data Table 1 for model details.
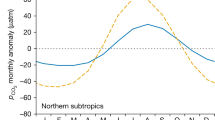
Extended Data Figure 4 Comparison of present-day natural CO2 variability as predicted by model and data.
Taylor diagram comparing the ability of ESMs to capture the dynamics and magnitude of the annual cycle of CO2 with our data-based approach. Here, the open circle on the x axis represents the standard deviation in our data-based predictions, while the grey contour lines represent the residual standard error between the ESMs and our data-based predictions.
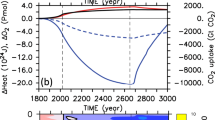
Extended Data Figure 5 Interannual variability of future CO2 amplification.
CO2 amplification factors within the HadGEM2 model for each year from 2097 to 2100.
Extended Data Figure 6 Nonlinearity of future CO2 distributions between SOMLO and models.
Surface  distribution (normalized such that the mean
distribution (normalized such that the mean  level across the whole surface ocean is shifted to zero) as predicted by SOMLO and two climate models (CanESM2 and HadGEM2-ES), highlighting the change in distribution between 2006 and 2100. The nonlinearity is evidenced by ‘bulging’ in the 2100 distributions for positive
level across the whole surface ocean is shifted to zero) as predicted by SOMLO and two climate models (CanESM2 and HadGEM2-ES), highlighting the change in distribution between 2006 and 2100. The nonlinearity is evidenced by ‘bulging’ in the 2100 distributions for positive  , relative to those for negative
, relative to those for negative  .
.
Rights and permissions
About this article

Cite this article
McNeil, B., Sasse, T. Future ocean hypercapnia driven by anthropogenic amplification of the natural CO2 cycle. Nature 529, 383–386 (2016). https://doi.org/10.1038/nature16156
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature16156
This article is cited by
-
Arctic Ocean annual high in \({{\boldsymbol{p}}}_{{{\bf{CO}}}_{{\bf{2}}}}\) could shift from winter to summer
Nature (2022)
-
Direct effects of elevated dissolved CO2 can alter the life history of freshwater zooplankton
Scientific Reports (2022)
-
Alternative Biogeochemical States of River Pools Mediated by Hippo Use and Flow Variability
Ecosystems (2021)
-
Spontaneously producing syngas from MFC-MEC coupling system based on biocompatible bifunctional metal-free electrocatalyst
Science China Materials (2021)
-
Ocean acidification does not impair the behaviour of coral reef fishes
Nature (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
