
Abstract
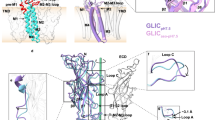
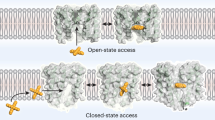
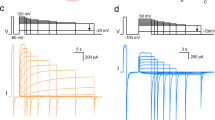
The X-ray structure of a pentameric ligand-gated ion channel from Erwinia chrysanthemi (ELIC) has recently provided structural insight into this family of ion channels at high resolution1. The structure shows a homo-pentameric protein with a barrel-stave architecture that defines an ion-conduction pore located on the fivefold axis of symmetry. In this structure, the wide aqueous vestibule that is encircled by the extracellular ligand-binding domains of the five subunits narrows to a discontinuous pore that spans the lipid bilayer. The pore is constricted by bulky hydrophobic residues towards the extracellular side, which probably serve as barriers that prevent the diffusion of ions. This interrupted pore architecture in ELIC thus depicts a non-conducting conformation of a pentameric ligand-gated ion channel, the thermodynamically stable state in the absence of bound ligand. As ligand binding promotes pore opening in these ion channels and the specific ligand for ELIC has not yet been identified, we have turned our attention towards a homologous protein from the cyanobacterium Gloebacter violaceus (GLIC). GLIC was shown to form proton-gated channels that are activated by a pH decrease on the extracellular side and that do not desensitize after activation2. Both prokaryotic proteins, ELIC and GLIC form ion channels that are selective for cations over anions with poor discrimination among monovalent cations1,2, characteristics that resemble the conduction properties of the cation-selective branch of the family that includes acetylcholine and serotonin receptors3,4. Here we present the X-ray structure of GLIC at 3.1 Å resolution. The structure reveals a conformation of the channel that is distinct from ELIC and that probably resembles the open state. In combination, both structures suggest a novel gating mechanism for pentameric ligand-gated ion channels where channel opening proceeds by a change in the tilt of the pore-forming helices.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
References
Hilf, R. J. & Dutzler, R. X-ray structure of a prokaryotic pentameric ligand-gated ion channel. Nature 452, 375–379 (2008)
Bocquet, N. et al. A prokaryotic proton-gated ion channel from the nicotinic acetylcholine receptor family. Nature 445, 116–119 (2007)
Karlin, A. Emerging structure of the nicotinic acetylcholine receptors. Nature Rev. Neurosci. 3, 102–114 (2002)
Adams, D. J., Dwyer, T. M. & Hille, B. The permeability of endplate channels to monovalent and divalent metal cations. J. Gen. Physiol. 75, 493–510 (1980)
Tasneem, A., Iyer, L. M., Jakobsson, E. & Aravind, L. Identification of the prokaryotic ligand-gated ion channels and their implications for the mechanisms and origins of animal Cys-loop ion channels. Genome Biol. 6, R4 (2004)
Miyazawa, A., Fujiyoshi, Y. & Unwin, N. Structure and gating mechanism of the acetylcholine receptor pore. Nature 423, 949–955 (2003)
Unwin, N. Refined structure of the nicotinic acetylcholine receptor at 4Å resolution. J. Mol. Biol. 346, 967–989 (2005)
Imoto, K. et al. Rings of negatively charged amino acids determine the acetylcholine receptor channel conductance. Nature 335, 645–648 (1988)
Konno, T. et al. Rings of anionic amino acids as structural determinants of ion selectivity in the acetylcholine receptor channel. Proc. R. Soc. Lond. B 244, 69–79 (1991)
Zhou, Y., Morais-Cabral, J. H., Kaufman, A. & MacKinnon, R. Chemistry of ion coordination and hydration revealed by a K+ channel-Fab complex at 2.0 Å resolution. Nature 414, 43–48 (2001)
Sine, S. M. & Engel, A. G. Recent advances in Cys-loop receptor structure and function. Nature 440, 448–455 (2006)
Brejc, K. et al. Crystal structure of an ACh-binding protein reveals the ligand-binding domain of nicotinic receptors. Nature 411, 269–276 (2001)
Celie, P. H. et al. Nicotine and carbamylcholine binding to nicotinic acetylcholine receptors as studied in AChBP crystal structures. Neuron 41, 907–914 (2004)
Lee, W. Y. & Sine, S. M. Principal pathway coupling agonist binding to channel gating in nicotinic receptors. Nature 438, 243–247 (2005)
Sala, F., Mulet, J., Sala, S., Gerber, S. & Criado, M. Charged amino acids of the N-terminal domain are involved in coupling binding and gating in alpha7 nicotinic receptors. J. Biol. Chem. 280, 6642–6647 (2005)
Schofield, C. M., Jenkins, A. & Harrison, N. L. A highly conserved aspartic acid residue in the signature disulfide loop of the alpha 1 subunit is a determinant of gating in the glycine receptor. J. Biol. Chem. 278, 34079–34083 (2003)
Jha, A., Cadugan, D. J., Purohit, P. & Auerbach, A. Acetylcholine receptor gating at extracellular transmembrane domain interface: The cys-loop and M2–M3 linker. J. Gen. Physiol. 130, 547–558 (2007)
Dani, J. A. Open channel structure and ion binding sites of the nicotinic acetylcholine receptor channel. J. Neurosci. 9, 884–892 (1989)
Dani, J. A. & Eisenman, G. Monovalent and divalent cation permeation in acetylcholine receptor channels. Ion transport related to structure. J. Gen. Physiol. 89, 959–983 (1987)
Galzi, J. L. et al. Mutations in the channel domain of a neuronal nicotinic receptor convert ion selectivity from cationic to anionic. Nature 359, 500–505 (1992)
Corringer, P. J. et al. Mutational analysis of the charge selectivity filter of the alpha7 nicotinic acetylcholine receptor. Neuron 22, 831–843 (1999)
Gunthorpe, M. J. & Lummis, S. C. Conversion of the ion selectivity of the 5-HT(3a) receptor from cationic to anionic reveals a conserved feature of the ligand-gated ion channel superfamily. J. Biol. Chem. 276, 10977–10983 (2001)
Pascual, J. M. & Karlin, A. State-dependent accessibility and electrostatic potential in the channel of the acetylcholine receptor. Inferences from rates of reaction of thiosulfonates with substituted cysteines in the M2 segment of the alpha subunit. J. Gen. Physiol. 111, 717–739 (1998)
Wilson, G. G., Pascual, J. M., Brooijmans, N., Murray, D. & Karlin, A. The intrinsic electrostatic potential and the intermediate ring of charge in the acetylcholine receptor channel. J. Gen. Physiol. 115, 93–106 (2000)
Unwin, N. Acetylcholine receptor channel imaged in the open state. Nature 373, 37–43 (1995)
Paas, Y. et al. Pore conformations and gating mechanism of a Cys-loop receptor. Proc. Natl Acad. Sci. USA 102, 15877–15882 (2005)
Cymes, G. D., Ni, Y. & Grosman, C. Probing ion-channel pores one proton at a time. Nature 438, 975–980 (2005)
Cymes, G. D. & Grosman, C. Pore-opening mechanism of the nicotinic acetylcholine receptor evinced by proton transfer. Nature Struct. Mol. Biol. 15, 389–396 (2008)
Kabsch, W. Automatic processing of rotation diffraction data from crystals of initially unknown symmetry and cell constants. J. Appl. Crystallogr. 26, 795–800 (1993)
Leslie, A. G. Recent changes to the MOSFLM package for processing film and image plate data. Joint CCP4 and ESF-EACBM Newsletter on Protein Crystallography (No. 26, Daresbury Laboratory, 1992)
Evans, P. Scaling and assessment of data quality. Acta Crystallogr. D 62, 72–82 (2006)
CCP4. Collaborative Computational Project Nr. 4. The CCP4 Suite: Programs for X-ray crystallography. Acta Crystallogr. D 50, 760–763 (1994)
McCoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658–674 (2007)
Cowtan, K. An automated procedure for phase improvement by density modification. Joint CCP4 and ESF-EACBM Newsletter on Protein Crystallography 34–38 (No. 31, Daresbury Laboratory, 1994)
Jones, T. A., Zou, J. Y., Cowan, S. W. & Kjeldgaard, M. Improved methods for building protein models in electron density maps and the location of errors in these models. Acta Crystallogr. A 47, 110–119 (1991)
Brunger, A. T. et al. Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr. D 54, 905–921 (1998)
Adams, P. D. et al. PHENIX: Building new software for automated crystallographic structure determination. Acta Crystallogr. D 58, 1948–1954 (2002)
Smart, O. S., Neduvelil, J. G., Wang, X., Wallace, B. A. & Sansom, M. S. HOLE: A program for the analysis of the pore dimensions of ion channel structural models. J. Mol. Graph. 14, 354–360, 376 (1996)
Sanner, M. F., Olson, A. J. & Spehner, J. C. Reduced surface: An efficient way to compute molecular surfaces. Biopolymers 38, 305–320 (1996)
Brooks, B. R. et al. CHARMM: A program for macromolecular energy, minimization, and dynamics calculations. J. Comput. Chem. 4, 187–217 (1983)
Im, W., Beglov, D. & Roux, B. Continuum solvation model: Electrostatic forces from numerical solutions to the Poisson-Boltzmann equation. Comput. Phys. Commun. 111, 59–75 (1998)
Lorenz, C., Pusch, M. & Jentsch, T. J. Heteromultimeric CLC chloride channels with novel properties. Proc. Natl Acad. Sci. USA 93, 13362–13366 (1996)
Acknowledgements
We thank B. Blattmann and C. Stutz-Duocommun for assistance with crystal screening, C. Schulze-Briese and the staff of the X06SA beamline for support during data collection, the protein analysis group at the functional genomics centre of the University of Zurich for help with mass spectrometry, R. MacKinnon for comments on the manuscript and members of the Dutzler laboratory for help in all stages of the project. Data collection was performed at the Swiss Light Source of the Paul Scherrer Institute. The research leading to these results was supported by a grant from the National Center for Competence in Research (NCCR) in Structural Biology and by an EC FP7 grant for the EDICT consortium (HEALTH-201924). R.J.C.H. is affiliated with the Molecular Life Sciences Ph.D. programme of the University/ETH Zurich.
Author Contributions R.D. and R.J.C.H. designed the project. R.J.C.H. carried out all experiments. R.D. assisted in data collection and structure determination. R.D. and R.J.C.H. jointly wrote the manuscript.
Author information
Authors and Affiliations
Corresponding author
Supplementary information
Supplementary Information
This file contains Supplementary Figures S1-S10 with Legends and Supplementary Tables S1 -S2. (PDF 2340 kb)
Supplementary Movie 1
Supplementary Movie 1 contains a morph of conserved parts of the Ca trace between ELIC and GLIC. The view is from the extracellular side. (MPG 1711 kb)
Supplementary Movie 2
Supplementary Movie 2 contains a morph of conserved parts of the Ca trace between ELIC and GLIC. The view is from within the membrane. (MPG 1583 kb)
Rights and permissions
About this article
Cite this article
Hilf, R., Dutzler, R. Structure of a potentially open state of a proton-activated pentameric ligand-gated ion channel. Nature 457, 115–118 (2009). https://doi.org/10.1038/nature07461
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature07461
This article is cited by
-
Origin of acetylcholine antagonism in ELIC, a bacterial pentameric ligand-gated ion channel
Communications Biology (2022)
-
General anesthesia bullies the gut: a toxic relationship with dysbiosis and cognitive dysfunction
Psychopharmacology (2022)
-
Control of structural flexibility of layered-pillared metal-organic frameworks anchored at surfaces
Nature Communications (2019)
-
Membrane directed expression in Escherichia coli of BBA57 and other virulence factors from the Lyme disease agent Borrelia burgdorferi
Scientific Reports (2019)
-
The roles of aromatic residues in the glycine receptor transmembrane domain
BMC Neuroscience (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
