
Abstract
All viruses rely on host cell proteins and their associated mechanisms to complete the viral life cycle. Identifying the host molecules that participate in each step of virus replication could provide valuable new targets for antiviral therapy, but this goal may take several decades to achieve with conventional forward genetic screening methods and mammalian cell cultures. Here we describe a novel genome-wide RNA interference (RNAi) screen in Drosophila1 that can be used to identify host genes important for influenza virus replication. After modifying influenza virus to allow infection of Drosophila cells and detection of influenza virus gene expression, we tested an RNAi library against 13,071 genes (90% of the Drosophila genome), identifying over 100 for which suppression in Drosophila cells significantly inhibited or stimulated reporter gene (Renilla luciferase) expression from an influenza-virus-derived vector. The relevance of these findings to influenza virus infection of mammalian cells is illustrated for a subset of the Drosophila genes identified; that is, for three implicated Drosophila genes, the corresponding human homologues ATP6V0D1, COX6A1 and NXF1 are shown to have key functions in the replication of H5N1 and H1N1 influenza A viruses, but not vesicular stomatitis virus or vaccinia virus, in human HEK 293 cells. Thus, we have demonstrated the feasibility of using genome-wide RNAi screens in Drosophila to identify previously unrecognized host proteins that are required for influenza virus replication. This could accelerate the development of new classes of antiviral drugs for chemoprophylaxis and treatment, which are urgently needed given the obstacles to rapid development of an effective vaccine against pandemic influenza and the probable emergence of strains resistant to available drugs.
Similar content being viewed by others


Main
Influenza, a highly contagious disease of birds and mammals, is caused by negative-strand RNA viruses of the family Orthomyxoviridae. Influenza outbreaks kill millions of people worldwide during pandemic years and hundreds of thousands during other years. Since their first lethal infection of humans in 1997, H5N1 influenza A viruses have spread throughout Asia and to Europe and Africa, posing a major risk for a new influenza pandemic2. To provide rational bases for improved treatment and control of influenza virus infection, we sought to advance understanding of viral infection mechanisms by elucidating previously unknown virus–host cell interactions. Many steps in the viral life cycle, including intracellular trafficking, gene expression, replication and virion assembly, depend on interactions with specific host cell gene products. Although most such host molecules remain elusive, emerging results indicate that their identification and characterization can provide new insights into the mechanisms by which viruses complete their life cycle, and hence illuminate potentially valuable targets for prophylactic and therapeutic intervention3,4,5.
Systematic, genome-wide RNAi analysis offers an exciting tool to identify host genes that function in viral replication. Such analysis is facilitated by well-developed model systems such as Drosophila, the genome of which contains only ∼14,000 genes, nearly all of which can be specifically targeted for high efficiency messenger RNA depletion by double-stranded RNA (dsRNA) libraries1. Because of its powerful genetics and conservation with vertebrates, Drosophila has been used to make numerous critical contributions to mammalian cell biology6,7,8,9. Thus, in principle, Drosophila RNAi studies could accelerate identification of host interactions essential for influenza virus replication.
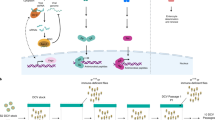
Because Drosophila D-Mel2 cells do not express the human influenza virus receptor α2,6-linked sialic acid (Supplementary Fig. 1), we predicted that wild-type human influenza virus would not be able to infect them. Indeed, we did not detect viral protein expression by immunofluorescence assays in Drosophila D-Mel2 cells inoculated with influenza virus A/WSN/33 (WSN; H1N1) (data not shown). To bypass this block to wild-type influenza virus entry, we generated a genetically modified virus, Flu-VSV-G-GFP (FVG-G), in Madin-Darby canine kidney (MDCK) cells by replacing the receptor-binding haemagglutinin (HA) and neuraminidase (NA) genes with genes encoding vesicular stomatitis virus glycoprotein G (VSV-G) and enhanced green fluorescence protein (eGFP), respectively10,11 (Fig. 1a). Because the envelopes of the resulting virions bear VSV-G, which mediates entry into mammalian, Drosophila and other cells12, FVG-G virions should readily infect Drosophila cells. Twenty-four hours after infection, GFP fluorescence was detected in FVG-G-infected Drosophila D-Mel2 cells (Supplementary Fig. 2). We also confirmed by real-time polymerase chain reaction (PCR) that influenza virus RNA replication occurred in Drosophila cells (Supplementary Fig. 3). However, Drosophila cells infected with FVG-G did not release detectable virions into the medium, as assayed by infectivity tests on MDCK cells and by electron microscopy (data not shown). This was due, at least in part, to failure of Drosophila cells to express some viral proteins required for virion assembly and infectivity (Supplementary Fig. 4). Thus, Drosophila cells can support influenza virus replication from post-entry to at least the protein expression phase of the viral life cycle. This span encompasses multiple other steps in the life cycle, including cytoplasmic release of genomic RNA-containing viral ribonucleoprotein complexes (vRNPs), vRNP import into the nucleus, mRNA synthesis from the negative-strand viral RNA genome, mRNA export to the cytoplasm and translation.

a, b, Schematic diagrams showing recombinant influenza viruses. Shown are FVG-G, in which genes encoding the HA and NA proteins were replaced with the VSV-G and eGFP genes, respectively (a), and FVG-R, in which the genes encoding the HA and NA were replaced with the VSV-G and Renilla luciferase genes, respectively (b). c, Schematic diagram of the systematic analysis of host genes affecting influenza virus replication and gene expression in Drosophila cells. Experimental details are given in Methods.
For high-throughput, functional genomics analysis of influenza virus replication in Drosophila cells, we engineered Flu-VSV-G-R.Luc (FVG-R), in which VSV-G and Renilla luciferase genes replaced the viral HA and NA open reading frames (Fig. 1b). FVG-R virions were then used with an RNAi library (Ambion) against 13,071 Drosophila genes (∼90% of all genes) to identify host genes affecting influenza-virus-directed Renilla luciferase expression (Fig. 1c). Two independent tests of the entire library were performed (Supplementary Table 1). For 176 genes for which dsRNAs inhibited FVG-R-directed luciferase expression in both replicates, repeated secondary tests using alternate dsRNAs to control for possible off-target effects confirmed the effects of 110 genes (Supplementary Tables 2 and 3). This confirmation rate is comparable to that in a Drosophila screen with a natural Drosophila-infecting virus5. Cell viability testing identified six genes with potentially significant cytotoxic effects; these were excluded from further consideration (Supplementary Information and Supplementary Table 3). Secondary tests of candidate genes for which dsRNAs increased FVG-R-directed luciferase expression produced a much lower confirmation rate, suggesting a higher rate of off-target or other false-positive effects in this class (Supplementary Information and Supplementary Table 4).
Among the over 100 candidate genes found to be important for influenza virus replication in Drosophila cells, we selected the human homologues of several encoding components in host pathways/machineries that are known to be involved in the life cycle of influenza virus, for example, ATP6V0D1 (endocytosis pathway), COX6A1 (mitochondrial function) and NXF1 (mRNA nuclear export machinery), for further analysis in mammalian cells to assess the relevance of our Drosophila results13,14,15,16,17. ATP6V0D1 encodes subunit D of vacuolar (H+)-ATPase (V-ATPase), a proton pump that functions in the endocytosis pathway (that is, the acidification and fusion of intracellular compartments18). COX6A1 encodes a subunit of cytochrome c oxidase (COX), an enzyme of the mitochondrial electron transport chain that catalyses electron transfer from cytochrome c to oxygen19. NXF1 encodes a nuclear export factor critical for exporting most cellular mRNAs containing exon–exon junctions20,21.
As a first test for the possible contribution of these gene products to influenza virus replication in mammalian cells, we treated human HEK 293 cells twice at 24-h intervals with short interfering RNAs (siRNAs; siGENOME, Dharmacon) against the human homologue of each selected Drosophilia gene. Twenty-four hours after the second siRNA treatment, the cells were infected with FVG-R virus and, two days later, Renilla luciferase activity was measured to assess viral replication and gene expression. siRNA against ATP6V0D1 or COX6A1 markedly decreased Renilla luciferase activity (Fig. 2a), but not cell viability (Supplementary Fig. 5a), suggesting that these genes have important roles in influenza virus replication in mammalian cells, as in Drosophila cells. Inhibition was not caused by off-target effects because, for each gene, each of four distinct siRNAs inhibited FVG-R-directed expression of Renilla luciferase (Supplementary Table 6). Because COX6A1 encodes a subunit of mitochondrial electron transport chain complex IV, COX, we used specific inhibitors to test whether in HEK 293 cells influenza virus also required other complexes in this chain (Fig. 2c). Inhibitors of complexes III, IV and V selectively inhibited FVG-R-directed Renilla luciferase expression by 50- to 100-fold, whereas complex I and II inhibitors had little or no effect. Thus, in mammalian cells, influenza virus depends on multiple late stages but not early stages in the mitochondrial electron transport chain.

a–c, Renilla luciferase activity was measured in FVG-R-infected HEK 293 cells treated with siRNAs against ATP6V0D1 and COX6A1 (a), NXF1 (b) or the indicated mitochondrial electron transport chain inhibitors (c). Inhibitors of complexes III, IV and V inhibited FVG-R-directed Renilla luciferase expression significantly, whereas complex I and II inhibitors had little or no effect. In contrast, the inhibitors had no significant effects on cell viability and Gaussia luciferase expression of a murine leukaemia virus derivative (MLV-GL) that, like FVG-R, depended on VSV G-mediated entry. All experiments were conducted three times in duplicate, with the results reported as mean ± s.d. 3-NPA, 3-nitroproprionic acid.
Treatment for four days with siRNA against NXF1 decreased mammalian cell viability (data not shown), as predicted by the critical role of NXF1 in general host cell metabolism. Accordingly, the total incubation time with siRNA against NXF1 was shortened to 36 h by transfecting cells with the siRNA twice at a 12-h interval, infecting with FVG-R virus 12 h later, and assaying for Renilla luciferase at 12-h post-infection. Under these conditions, cell viability was not detectably affected (Supplementary Fig. 5b) whereas Renilla luciferase activity was reduced by nearly fivefold (Fig. 2b). Whereas recent results indicated that influenza virus protein NS1 binds to NXF1 to inhibit host mRNA export17, these results imply that influenza virus RNAs and/or proteins are transported by an NXF1-dependent pathway (see also Supplementary Information).
To test the effects of these genes on authentic influenza viruses, we infected siRNA-treated HEK 293 cells with WSN virus or H5N1 influenza A/Indonesia/7/05 (Indonesia 7; isolated from a patient) or with VSV or vaccinia virus as controls. Progeny viruses were collected from the medium at 24 h (Indonesia 7, VSV or vaccinia virus) or 48 h (WSN) post-infection and were titrated. Depleting ATP6V0D1 and COX6A1 did not affect VSV or vaccinia virus replication, but decreased the WSN and Indonesia 7 virus yields by ∼10-fold or more (Fig. 3a). Thus, ATP6V0D1 and COX6A1 are required for replication of influenza viruses but not VSV and vaccinia virus. Depleting the NXF1 nuclear export factor, again using an accelerated 36-h time line, reduced Indonesia 7 virus titres by 20-fold (Fig. 3b), indicating that NXF1 has a critical role in H5N1 influenza virus replication. Unfortunately, because WSN grows in HEK 293 cells more slowly than does Indonesia 7 virus, the WSN virus yield at 12-h post-infection was insufficient to test for an effect of NXF1 depletion (data not shown). In comparison to Indonesia 7 virus, NXF1 depletion had no effect on VSV or vaccinia virus yields (Fig. 3b). Moreover, siRNA against NXF1 inhibited FVG-G, but not adenovirus, which, like influenza virus, depends on nuclear steps for genome transcription and replication (Supplementary Fig. 6). Thus, multiple genes identified in Drosophila cells correspond to important, selective host factors for influenza virus replication in mammalian cells.

a, b, The titres of influenza viruses (WSN and Indonesia 7), VSV and vaccinia virus in HEK 293 cells treated with siRNA against ATP6V0D1 and COX6A1 (a) or against NXF1 (b) are shown. Experimental details are given in Methods and Supplementary Methods. All experiments were conducted three times, with the results reported as mean ± s.d.
The method that we have established, using systematic analysis of a Drosophila RNAi library with confirmation in mammalian cells, can be used to identify host gene products that affect influenza virus replication. This utility was demonstrated in experiments with authentic influenza viruses, including an H5N1 virus isolated from a human patient. One of the candidate genes tested in mammalian cells, COX6A1, which encodes a subunit of COX, acts as a critical enzyme in cytochrome-c-dependent electron transport in mitochondria. Influenza virus PB2 polymerase has a mitochondrial targeting signal14, and PB1-F2, the second protein encoded by the influenza PB1 gene, was reported to localize to inner and outer mitochondrial membranes and to delay influenza virus clearance by host antiviral responses15. Thus, COX6A1 may be involved in PB2- and/or PB1-F2-mediated functions in mitochondria. Additionally, influenza vRNP export from the nucleus requires caspase 3 activation22, which can be induced by cytochrome c release from mitochondria. Moreover, human BCL2 inhibits both cytochrome c release and influenza vRNP export23, and COX function and changes in mitochondrial membrane potential have been linked to caspase activation24. These pathways may underlie our further findings (Fig. 2c) that influenza virus replication and expression in mammalian cells are strongly and selectively modulated by compounds that inhibit COX, cytochrome c-linked electron transport chain complex III, and normal ion transport across mitochondrial membranes (oligomycin and valinomycin). Potential roles of the other two host genes confirmed here in mammalian cells, ATP6V0D1 and NXF1, in influenza virus replication are discussed further in the Supplementary Information.
The above results with multiple, diverse genes, including ATP6V0D1, NXF1 and COX6A1 as well as mitochondrial electron transport complexes III and V, demonstrate the feasibility and value of using Drosophila RNAi screening to identify previously unknown host factors with important and potentially unsuspected roles in influenza virus replication. Simultaneously, the genome-wide results from our Drosophila RNAi screen provide more than 100 additional candidate genes (many with unknown functions) to be tested in mammalian cells. We suggest that the same strategy could be applied to identify previously unknown host factors involved in the replication of other viruses, whenever at least a portion of their replication cycle is supported by Drosophila cells.
Methods Summary
Cells and viruses
MDCK cells, HEK 293 cells, baby hamster kidney (BHK) cells, D-Mel2 cells and DL1 cells were maintained as described in Methods. WSN, FVG-G, FVG-R and Indonesia 7 viruses were generated as described previously10 and propagated in MDCK cells. VSV and vaccinia virus were grown in BHK and CV-1 cells, respectively. Gaussia-luciferase-expressing murine leukaemia virus (MLV-GL) and the GFP-expressing adenovirus were provided by J. Bruce and R. Kalejta25, respectively.
Drosophila RNAi library analysis
Double-stranded RNA of the Drosophila RNAi library (targeting 13,071 Drosophila genes) and DL1 cells were added to each well of 384-well microplates. After two days of incubation, cells were infected with the FVG-R virus. At one day post-infection, Renilla luciferase activity was measured as described below. Two independent analyses of the entire library were performed.
siRNA treatment of mammalian cells
HEK 293 cells were transfected with siRNA by TransIT-TKO (Mirus). Cells were incubated for 12 h in experiments with NXF1 or for 24 h with other genes, were transfected again under the same conditions, and inoculated with virus after 12 h for NXF1 or after 24 h for other genes. After a further 12 h (for NXF1) or 48 h (other genes), the cells were harvested for the indicated analyses.
Renilla luciferase and cell viability assays
Renilla luciferase activity and cell viability were measured with established Renilla luciferase and CellTiter-Glo assay systems (Promega) according to the manufacturer’s instructions; signals were read with a GLOMAX 96 microplate luminometer.
Online Methods
Cells and viruses
MDCK cells, HEK 293 cells and BHK cells were maintained in minimum essential medium containing 5% fetal calf serum and antibiotics at 37 °C in 5% CO2. D-Mel2 cells were maintained in Drosophila-SFM (GIBCO/Invitrogen) at 28 °C. DL1 cells were maintained in Schneider’s Drosophila (SD) medium containing 10% FBS at 28 °C.
WSN, FVG-G, FVG-R and Indonesia 7 viruses were generated by a plasmid-based reverse genetics system10 and were grown and titrated in MDCK cells. MLV-GL was produced by replacing the CD4 open reading frame in pCMMP-CD4-eGFP26 with that of Gaussia luciferase (J. Bruce, unpublished observations). All experiments with Indonesia 7 virus were conducted in a biosafety level 3 containment laboratory approved for such use by the Centers for Disease Control and Prevention and the US Department of Agriculture.
Drosophila RNAi library analysis
The Drosophila RNAi library (Ambion) contained 13,071 individual dsRNAs, each designed to specifically target a single Drosophila gene. Five microlitres of dsRNA (40 ng μl-1) were added to each well of 384-well plates, after which 2 × 104 DL1 cells in 10 μl of SD medium were added to each well and incubated with the dsRNA at 28 °C for 60 min. Twenty microlitres of SD medium containing 20% FBS was then added to each well after incubation. Cells were treated with dsRNA for two days at 28 °C and were then inoculated with an amount of FVG-R virus corresponding to a multiplicity of infection of 10 for MDCK cells, and were then transferred to 33 °C. At one day post-infection, Renilla luciferase activity was measured as described below. Two independent analyses of the entire library were performed.
siRNA treatment of mammalian cells
The non-targeted siRNA used was siCONTROL 1 (Dharmacon). Duplex siRNAs against candidate genes were obtained from siGENOME (Dharmacon, see Supplementary Table 7 for sequences); the effects of siRNA were evaluated by reverse transcription followed by PCR (RT–PCR; Supplementary Fig. 7). The sequences of siRNA against the NP gene of influenza virus (GGAUCUUAUUUCCUUCGGAGUU)27 and the E3L gene of vaccinia virus (AAUAUCGUCGGAGCUGUACAC)28 were reported previously. The sequence of siRNA against the gene encoding the L protein of VSV (CGAGUUAUCCAGCAAUCAUUU) was designed using BLOCK-iT RNAi designer (Invitrogen). A HEK 293 cell suspension was seeded into the wells of a 24-well plate (2.0 × 104 cells per well), incubated for 1 h and transfected with siRNA (to a final concentration 10 nM; Dharmacon) using TransIT-TKO (Mirus), according to the manufacturer’s instructions.
Inhibitor treatment of mammalian cells
Mitochondrial electron transport chain inhibitors were used at the following final concentrations: rotenone (0.5 μM), 3-nitroproprionic acid (1 mM), antimycin A (25 μM), NaN3 (50 mM), oligomycin (25 μM) and valinomycin (0.9 μM). HEK 293 cells were treated with the inhibitors for 5 h before being infected with FVG-R or MLV-GL, incubated 18 h, and assayed for Renilla or Guassia luciferase, respectively. Cell viability was assayed in parallel by a firefly-luciferase-mediated ATP assay (CellTiter-Glo, Promega).
References
Kuttenkeuler, D. & Boutros, M. Genome-wide RNAi as a route to gene function in Drosophila . Brief. Funct. Genomic. Proteomic. 3, 168–176 (2004)
Webster, R. G. & Govorkova, E. A. H5N1 influenza — continuing evolution and spread. N. Engl. J. Med. 355, 2174–2177 (2006)
Kushner, D. B. et al. Systematic, genome-wide identification of host genes affecting replication of a positive-strand RNA virus. Proc. Natl Acad. Sci. USA 100, 15764–15769 (2003)
Ludwig, S., Planz, O., Pleschka, S. & Wolff, T. Influenza-virus-induced signaling cascades: targets for antiviral therapy? Trends Mol. Med. 9, 46–52 (2003)
Cherry, S. et al. Genome-wide RNAi screen reveals a specific sensitivity of IRES-containing RNA viruses to host translation inhibition. Genes Dev. 19, 445–452 (2005)
Hahn, H., Wojnowski, L., Miller, G. & Zimmer, A. The patched signaling pathway in tumorigenesis and development: lessons from animal models. J. Mol. Med. 77, 459–468 (1999)
Dearolf, C. R. JAKs and STATs in invertebrate model organisms. Cell. Mol. Life Sci. 55, 1578–1584 (1999)
Lemaitre, B., Nicolas, E., Michaut, L., Reichhart, J. M. & Hoffmann, J. A. The dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 86, 973–983 (1996)
Takeda, K., Kaisho, T. & Akira, S. Toll-like receptors. Annu. Rev. Immunol. 21, 335–376 (2003)
Neumann, G. et al. Generation of influenza A viruses entirely from cloned cDNAs. Proc. Natl Acad. Sci. USA 96, 9345–9350 (1999)
Watanabe, T., Watanabe, S., Noda, T., Fujii, Y. & Kawaoka, Y. Exploitation of nucleic acid packaging signals to generate a novel influenza virus-based vector stably expressing two foreign genes. J. Virol. 77, 10575–10583 (2003)
Wyers, F., Richard-Molard, C., Blondel, D. & Dezelee, S. Vesicular stomatitis virus growth in Drosophila melanogaster cells: G protein deficiency. J. Virol. 33, 411–422 (1980)
Perez, L. & Carrasco, L. Involvement of the vacuolar H(+)-ATPase in animal virus entry. J. Gen. Virol. 75, 2595–2606 (1994)
Carr, S. M., Carnero, E., Garcia-Sastre, A., Brownlee, G. G. & Fodor, E. Characterization of a mitochondrial-targeting signal in the PB2 protein of influenza viruses. Virology 344, 492–508 (2006)
Zamarin, D., Ortigoza, M. B. & Palese, P. Influenza A virus PB1-F2 protein contributes to viral pathogenesis in mice. J. Virol. 80, 7976–7983 (2006)
Momose, F. et al. Cellular splicing factor RAF-2p48/NPI-5/BAT1/UAP56 interacts with the influenza virus nucleoprotein and enhances viral RNA synthesis. J. Virol. 75, 1899–1908 (2001)
Satterly, N. et al. Influenza virus targets the mRNA export machinery and the nuclear pore complex. Proc. Natl Acad. Sci. USA 104, 1853–1858 (2007)
Stevens, T. H. & Forgac, M. Structure, function and regulation of the vacuolar (H+)-ATPase. Annu. Rev. Cell Dev. Biol. 13, 779–808 (1997)
Gnaiger, E., Lassnig, B., Kuznetsov, A., Rieger, G. & Margreiter, R. Mitochondrial oxygen affinity, respiratory flux control and excess capacity of cytochrome c oxidase. J. Exp. Biol. 201, 1129–1139 (1998)
Cullen, B. R. Nuclear mRNA export: insights from virology. Trends Biochem. Sci. 28, 419–424 (2003)
Reed, R. & Cheng, H. TREX, SR proteins and export of mRNA. Curr. Opin. Cell Biol. 17, 269–273 (2005)
Wurzer, W. J. et al. Caspase 3 activation is essential for efficient influenza virus propagation. EMBO J. 22, 2717–2728 (2003)
Hinshaw, V. S., Olsen, C. W., Dybdahl-Sissoko, N. & Evans, D. Apoptosis: a mechanism of cell killing by influenza A and B viruses. J. Virol. 68, 3667–3673 (1994)
Kadenbach, B., Arnold, S., Lee, I. & Huttemann, M. The possible role of cytochrome c oxidase in stress-induced apoptosis and degenerative diseases. Biochim. Biophys. Acta 1655, 400–408 (2004)
He, T. C. et al. A simplified system for generating recombinant adenoviruses. Proc. Natl Acad. Sci. USA 95, 2509–2514 (1998)
Bruce, J. W., Bradley, K. A., Ahlquist, P. & Young, J. A. Isolation of cell lines that show novel, murine leukemia virus-specific blocks to early steps of retroviral replication. J. Virol. 79, 12969–12978 (2005)
Ge, Q. et al. RNA interference of influenza virus production by directly targeting mRNA for degradation and indirectly inhibiting all viral RNA transcription. Proc. Natl Acad. Sci. USA 100, 2718–2723 (2003)
Dave, R. S. et al. siRNA targeting vaccinia virus double-stranded RNA binding protein [E3L] exerts potent antiviral effects. Virology 348, 489–497 (2006)
Acknowledgements
We thank M. McGregor and K. Wells for technical assistance, M. Hatta, S. Yamada and M. Ito for an H5N1 influenza virus, S. Watanabe and Y. Hatta for FVG-R and FVG-G viruses, R. Kalejta and J. Bruce for adenovirus and murine leukaemia virus vectors, T. Noda for electron microscopy, J. Gilbert for editing the manuscript and Y. Kawaoka for illustrations. This work was supported, in part, by US National Institute of Allergy and Infectious Diseases Public Health Service research grants, by the US National Institute of General Medical Sciences grant GM35072, by a grant-in-aid for Specially Promoted Research and by a contract research fund for the Program of Funding Research Centers for Emerging and Reemerging Infectious Diseases from the Ministry of Education, Culture, Sports, Science and Technology, and by grants-in-aid from the Ministry of Health, Labor, Welfare of Japan. P.A. is a Howard Hughes Medical Institute investigator.
Author Contributions L.H., A.S., P.A. and Y.K. designed research; L.H., A.S., T.W. and C.A.N. performed research; L.H., A.S., T.W., E.S., M.A.N., P.A. and Y.K. analysed data; and L.H., A.S., T.W., P.A. and Y.K. wrote the manuscript.
Author information
Authors and Affiliations
Corresponding authors
Supplementary information
Supplementary Information 1
The file contains Supplementary Methods, additional references, and Supplementary Figures S1-S9 with Legends. (PDF 2686 kb)
Supplementary Information 2
The file contains Supplementary Table S1: Genome-wide primary screen data for influenza virus replication. (PDF 690 kb)
Supplementary Information 3
The file contains Supplementary Tables S2-S7. (PDF 711 kb)
Rights and permissions
About this article
Cite this article
Hao, L., Sakurai, A., Watanabe, T. et al. Drosophila RNAi screen identifies host genes important for influenza virus replication. Nature 454, 890–893 (2008). https://doi.org/10.1038/nature07151
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature07151
This article is cited by
-
M6PR interacts with the HA2 subunit of influenza A virus to facilitate the fusion of viral and endosomal membranes
Science China Life Sciences (2024)
-
Influenza virus mRNAs encode determinants for nuclear export via the cellular TREX-2 complex
Nature Communications (2023)
-
Host factor TNK2 is required for influenza virus infection
Genes & Genomics (2023)
-
RUNX1 inhibits the antiviral immune response against influenza A virus through attenuating type I interferon signaling
Virology Journal (2022)
-
Cellular glycan modification by B3GAT1 broadly restricts influenza virus infection
Nature Communications (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
