
Abstract
Volume deficits of the hippocampus in schizophrenia have been consistently reported. However, the hippocampus is anatomically heterogeneous; it remains unclear whether certain portions of the hippocampus are affected more than others in schizophrenia. In this study, we aimed to determine whether volume deficits in schizophrenia are confined to specific subfields of the hippocampus and to measure the subfield volume trajectories over the course of the illness. Magnetic resonance imaging scans were obtained from Data set 1: 155 patients with schizophrenia (mean duration of illness of 7 years) and 79 healthy controls, and Data set 2: an independent cohort of 46 schizophrenia patients (mean duration of illness of 18 years) and 46 healthy controls. In addition, follow-up scans were collected for a subset of Data set 1. A novel, automated method based on an atlas constructed from ultra-high resolution, post-mortem hippocampal tissue was used to label seven hippocampal subfields. Significant cross-sectional volume deficits in the CA1, but not of the other subfields, were found in the schizophrenia patients of Data set 1. However, diffuse cross-sectional volume deficits across all subfields were found in the more chronic and ill schizophrenia patients of Data set 2. Consistent with this pattern, the longitudinal analysis of Data set 1 revealed progressive illness-related volume loss (~2–6% per year) that extended beyond CA1 to all of the other subfields. This decline in volume correlated with symptomatic worsening. Overall, these findings provide converging evidence for early atrophy of CA1 in schizophrenia, with extension to other hippocampal subfields and accompanying clinical sequelae over time.
Similar content being viewed by others

Introduction
Abnormalities of the hippocampus are among the most consistently reported findings in studies of schizophrenia, and have been hypothesized to underlie the neuropsychological deficits and symptoms observed in the disorder.1, 2, 3 Meta-analyses of numerous structural magnetic resonance imaging (MRI) studies show reductions of the hippocampus in patients in both the early and chronic stages of illness.4, 5, 6 A recent large-scale multisite consortium study found that among the subcortical regions examined in schizophrenia, the largest magnitude of volume deficits was in the hippocampus.7 However, it is less clear whether the volume deficits of the hippocampus worsen during the course of illness, with some studies finding no atrophy over time8, 9, 10, 11 and other studies suggesting progressive volume loss that begins at early stages.12, 13, 14
It is also unclear whether certain portions of the hippocampus are affected more than others.15 The hippocampus is comprises the dentate gyrus (DG), Cornu Ammonis (CA) regions CA4, CA3, CA2 and CA1 of the hippocampus proper, and the subiculum (Sub).16, 17 The study of these cellularly demarcated, inter-connected hippocampal subfields, which have distinct functions,18, 19, 20, 21, 22 could offer insights into the underlying pathogenic mechanisms of hippocampal abnormalities in schizophrenia.3 With new advances in MRI data acquisition and analysis methods, many studies have shown that it is now possible to examine the subfields of the hippocampus separately.23, 24, 25, 26, 27, 28
Previous structural MRI studies of the hippocampal subfields in schizophrenia have produced mixed results. In cross-sectional studies of schizophrenia, semiautomated shape analyses—which involve manually tracing the perimeters of each individual subject’s hippocampus and high-dimensional mapping with a hippocampal anatomical template—have found deformity in regions corresponding to the CA1 in first-episode29 and chronic patients.30 Also, using an automated approach of labeling the subfields, one study reported volume reductions in CA1 and CA2/3 in chronic patients.31 However, two subsequent, larger-scale cross-sectional studies of subjects with chronic schizophrenia reported the greatest degree of volume reductions in the CA2/3, CA4/DG and Sub instead.32, 33 The discrepancies among these findings could be due to differences in the stages of illness of the schizophrenia patients examined, the image acquisition sequences or the methods used to delineate the hippocampal subfields. Surface-based shape analyses cannot adequately model the subfields that are embedded deep in the hippocampal formation, such as the DG and CA4.34 Also, the initial iteration of the automated method of labeling the subfields (used in the above-mentioned studies)32, 33 relied on an atlas constructed from in vivo hippocampal scans of limited MRI contrast.23, 35, 36 A newly developed approach, used in this study, relies on the much greater spatial resolution obtainable in ultra-high field scans of ex vivo hippocampal tissue.37 The higher level of segmentation accuracy associated with this approach should help clarify remaining questions about the distribution and time course of hippocampal volume loss in schizophrenia.
Hence in this study, we sought to determine the extent and trajectory of volume deficits of hippocampal subfields in schizophrenia, using this novel, automated method to label the subfields. We measured hippocampal subfield volume in two independent cohorts of schizophrenia patients and controls cross-sectionally; one cohort consisted of patients who were primarily in the early stages of illness, whereas the other cohort included a greater number of patients with chronic schizophrenia. Finally, a longitudinal analysis was performed in one cohort, to assess any changes in hippocampal subfield volumes that occur over the course of the illness.
Materials and methods
Participants
Written informed consent was obtained from all subjects in accordance with the guidelines of the National Healthcare Group (Singapore), National Neuroscience Institute (NNI, Singapore), Partners Healthcare and Harvard University (Boston, MA, USA) institutional review boards.
Clinically stable outpatients with schizophrenia were recruited at two sites: the Institute of Mental Health (IMH), Singapore, from 2006 to 2013 (Data set 1), and the Massachusetts General Hospital (MGH) in Boston, MA, USA, from 2008 to 2013 (Data set 2). Diagnosis of schizophrenia for the patients was confirmed by the Structured Clinical Interview for DSM-IV disorders (SCID)-Patient version.38 Healthy controls were recruited from the community at the same time through advertisements by the study team of KS at IMH, and DJH and JLR at MGH, and the Cognitive Neuroscience Laboratory based at both Harvard University and MGH. Healthy controls were screened using the SCID-Non-Patient (SCID-NP) interview; none had any history of Axis I disorders. Also, none of the participants had a history of neurological or neurodevelopmental disorders, or a diagnosis of substance or alcohol abuse 3 months preceding the study, claustrophobia or any other contraindications for having an MRI.
Demographic, clinical and imaging information for Data set 1 (155 patients and 79 controls) and Data set 2 (46 patients and 46 controls) is described in Supplementary Tables 1 and 2. In addition, we conducted a secondary analysis of a subset of 53 patients from Data set 1 in early stages of schizophrenia, who at the time of baseline data collection, had an onset of illness at an age <35 years, zero or just one hospitalization and <5 years of psychosis, and 61 demographically matched controls (Supplementary Table 3). Also, demographic information about the 34 patients and 41 controls (from Data set 1) that were followed up naturalistically for 2–7 years are described in Supplementary Table 4.
Symptom severity of all the patients was assessed using the Positive and Negative Symptom Scale (PANSS). 39 Also, estimates of antipsychotic dosages were calculated using daily chlorpromazine (CPZ) equivalent dosages.
MRI acquisition and image processing
All MRI scans were performed within 2 weeks after the clinical and neuropsychological assessments. All participants of Data set 1, including those scanned a second time 2–7 years later, were scanned on the same 3-Tesla whole-body scanner MRI (Philips Achieva, Best, The Netherlands) with an 8-channel SENSE (Sensitivity Encoding) head coil at the NNI, Singapore. Participants of Data set 2 were scanned on one of two identical 3-Tesla Tim Trio Siemens MRI scanners located at either the Athinoula A Martinos Center for Biomedical Imaging or at the Harvard University Center for Brain Science. There were no major scanner hardware or software upgrades during the MRI data collection period at either site. Scan parameters are detailed in Supplementary Methods 1.
Preprocessing of the structural images collected at both sites was performed using the open-source FreeSurfer pipeline (version 5.3, http://surfer.nmr.mgh.harvard.edu).40, 41, 42, 43 The longitudinal data were additionally processed using a specialized longitudinal processing stream where unbiased subject-specific templates were created, and then within-subject images for each time-point re-processed with common information from the template.44 This has been demonstrated to reduce the variability in within-subject morphological measures, and hence result in greater statistical power and the ability to detect small changes.44
To label the subfields, a new automated algorithm from FreeSurfer was used (Figure 1). This algorithm was based on a computational atlas built from ex vivo MRI data of post-mortem medial temporal tissue from 15 subjects, acquired at an average of 0.13 mm isotropic resolution on a 7-T scanner, and an in vivo atlas that provided information about adjacent extrahippocampal structures.37 Compared with the previous algorithm developed by FreeSurfer,45 the volumes generated by this new algorithm are more comparable with histologically based measurements of the subfields.37 It also provides a more comprehensive, fine-grained segmentation of the structures of the hippocampus, including areas such as the granule cell layer (GCL) of the DG, and the molecular layer (ML) of the CA fields and Sub. See Supplementary Methods 2 for comparisons of volume measures derived from both the older and new segmentation methods.

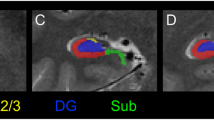
Representative subfield labels of the hippocampus generated by an automated ex vivo hippocampal segmentation approach. Subfield labels of the left anterior hippocampus of a representative healthy control subject from (a) Data set 1 and (b) Data set 2, in the sagittal, axial and coronal planes, are shown.
We measured volumes of the (i) overall hippocampus, generated by the widely used automated FreeSurfer subcortical segmentation script (based on the in vivo atlas)42 and (ii) seven structures considered to be subfields of the hippocampus:16, 17 the GCL, CA4, CA2/3 (CA2 and CA3 were combined in the atlas because of the lack of distinguishing MRI contrast), CA1, ML, the hippocampal tail (the posterior end of the hippocampus, which includes portions of the CA fields and DG undistinguishable with the MRI contrast) and the Sub.
Statistical analysis
All analyses were performed using open-source R software (version 3.1.3).46 Cross-sectional demographic differences between the patients and controls were tested using χ2 tests for categorical variables (gender, handedness and ethnicity) and F tests or independent t-tests for continuous variables (age, intracranial volume (ICV), CPZ and time between scans). Longitudinal change in clinical variables was assessed using paired t-tests (duration of illness, PANSS scores and CPZ). Differences in age, ICV, CPZ, duration of illnesses between the data sets were assessed using the Welch two-sample t-test of unequal variances.
Cross-sectional analyses (Data sets 1 and 2)
We first determined whether there were any group-based differences in the overall mean hippocampus volume of Data set 1 and Data set 2, as well as the early course patients of Data set 1 and matched controls. A multiple linear regression, with volume as the dependent variable, diagnosis as the main predictor, and ICV, age and gender as covariates, was conducted. We then investigated whether schizophrenia differentially affects the volume measures of the inter-related hippocampal subfields. The Shapiro–Wilk test and Bartlett’s test of homoscedasticity was first performed to ensure multivariate normality of the subfield volumes and equal variances of in the healthy control and patient groups of each data set, respectively. The subfield values were then log transformed. A multivariate analysis of covariance, with the 14 subfields as dependent (and correlated) variables, diagnosis as the main predictor, and ICV, age, gender and duration of illness as covariates, was conducted. The alpha was set at P<0.05. This was followed by a post-hoc univariate analysis of covariance to determine which subfield (dependent variable) contributed to the significant overall effect of illness.
Longitudinal analysis (Data set 1)
We sought to determine whether there was an interaction effect between diagnosis and time (between baseline and follow-up scans) for each hippocampal subfield. A separate multi-level model was constructed here, which accounted for the unevenly spaced time-points among subjects and the intra-individual variability in initial subfield volumes and their trajectories (Supplementary Methods 3). After model fitting, fixed effects included diagnosis, time, interaction between diagnosis and time, ICV, CPZ, age, and gender. Random effects included individual intercept and slope of time. The change in volume was modeled linearly, as volume trajectories in studies of gray matter in schizophrenia47 and the hippocampus in childhood-onset schizophrenia (which followed subjects until their late twenties)48 have been shown to be linear. In addition, the annualized rate of change in subfield volume measures was calculated: (Volumefollow-up – Volumebaseline)/(Volumebaseline × time).
Secondary analyses
As some studies have reported effects of treatment with antipsychotics,11 antidepressants49 and mood stabilizers50 on hippocampal structure, we repeated our primary analyses with the dosage or use of medication classes of antipsychotics, antidepressants and mood stabilizers as covariates. To minimize the potential confounding effect of ethnicity, we also repeated our analyses with ethnicity included as a covariate.
Post-hoc correlations between illness severity and hippocampal subfield volumes in subjects with schizophrenia
Cross-sectional analyses: We examined the relationship between clinical measures (PANSS subscales) and absolute volume measures of the subfields that were significantly different in the patients. A linear regression model was used, with the clinical measures as primary variables of interests, and age and sex among the covariates. We also tested the hypothesis that duration of illness is correlated with subfield volumes. In addition, we tested for correlations between antipsychotic dosages and subfield volumes.
Longitudinal analysis: We examined whether there was an intra-individual relationship between the rate of change in clinical symptoms—calculated by (Scoresfollow-up–Scoresbaseline)/(Scoresbaseline × time)—and the annualized rate of change in the subfield volume measures using similar regression modeling, controlling for age, sex, CPZ and baseline duration of illness.
To address the multiple testing for the various hypotheses, the Holm–Bonferroni method controlling for family-wise errors at alpha level (0.05) was applied.51
Results
Subject and cohort characteristics
Within each data set, the schizophrenia patients and controls were well-matched in terms of age, gender, ethnicity, and handedness. Between the cross-sectional Data sets 1 and 2, no cohort differences were found in the MRI estimates of head sizes, or in handedness or sex, for either the controls (t106.7=−0.14, P=0.89; χ2=0.34, P=0.56; χ2=1.7, P=0.20, respectively) or the patients (t81.7=0.23, P=0.82; χ2=1.24, P=0.54; χ2=1.39, P=0.25, respectively). Also, there was no difference in mean age of illness onset between the two patient cohorts (t79.33=1.11, P=0.28). However, as expected, the mean age of the subjects was higher in Data set 2 (patients: 42.9±10.2; controls: 41.9±9.1) compared with Data set 1 (patients 32.5±8.8 years; controls 31.2±9.9 years) (controls: t100.6=6.2, P=1.4 × 10−8; patients: t67.1=6.05, P=7 × 10−8). Also, compared with the patients of Data set 1, the patients of Data set 2 had a significantly longer mean duration of illness (Data set 1: 6.6±7.0 years; Data set 2:18.2±11.0 years) (t57.8=46.78, P=7 × 10−9), received higher mean daily doses of antipsychotic medication (Data set 1: 212.3±191.3 mg; Data set 2: 525.4±444.9 mg) (t51.3=4.7, P=2 × 10−5) and were more symptomatic (see Supplementary Tables 1; positive: t61.7=7.45, P=3.58 × 10−10; negative: t58.2=13.67, P=2.2 × 10−16; general psychopathology: t47.1=8.44, P=5.53 × 10−11).
Cross-sectional findings of Data set 1
Comparison of overall hippocampal volumes indicated that, compared with the controls, the schizophrenia patients had a smaller left hippocampus (β=−150.7, s.e.=52.1, t230 =−2.89, P=0.0042) and right hippocampus (β=−169.9, s.e.=47.7, t230=−3.56, P=0.00046). Following measurement of the volumes of the subfields, a significant effect of group on the combined volumes of the subfields GCL, CA4, CA2/3, CA1, ML, tail and Sub (Pillai’s trace=0.11, F14, 215=1.83, P=0.036) was observed (Figure 2a). Post-hoc testing, after correction for multiple comparisons, revealed a significant volume deficit in the left CA1 (but not in the other subfields) in the schizophrenia patients, relative to the controls (P=0.0010; Table 1a).

Cross-sectional hippocampal subfield volume deficits in schizophrenia. Group-based comparisons of the hippocampal subfield volumes in (a) Data set 1 and (b) Data set 2. In each data set, the subfield volumes of individual controls (blue) and schizophrenia subjects (red) are shown after co-adjusting for cohort-averaged head size. Multivariate analysis of covariance of the combined volumes of the 14 hippocampal subfields, followed by post-hoc univariate analysis of covariance and a Holm–Bonferroni correction for multiple comparisons across the 14 subfields showed that the volume deficit is limited to the CA1 in the schizophrenia patients who are at an early-to-mid stage of illness (Data set 1), whereas the volume deficits involve multiple subfields in chronic patients (Data set 2). *Indicates significance in corrected P-values controlling for family-wise error rate of alpha level of <0.05. GCL, granule cell layer; ML, molecular layer; Sub, subiculum.
The first 5 years of illness
To determine whether a change in CA1 is present during the earliest stages of schizophrenia, we conducted a secondary analysis in 53 patients in the first 5 years of illness and 61 healthy controls of Data set 1. This analysis showed reduced volumes of the left hippocampus (β=−154.8, s.e.=63.29, t110=−2.45, P=0.016) and right hippocampus (β=−179.6, s.e.=63.43, t110=−2.83, P=0.0055) in the patients (mean age 27.6±4.9 years), compared with the controls (mean age 27.2±4.7 years). There was a marginal effect of group on the combined subfield volumes (Pillai’s trace=0.12, F14,151=1.58, P=0.09). Post-hoc testing revealed that the subtle hippocampal volume deficit found in the schizophrenia patients of this subgroup was due to a smaller left and right CA1, as well as right GCL (Table 1b).
Cross-sectional findings of Data set 2
Comparisons of the overall hippocampal volumes showed a significant effect of group (that is, smaller in the patients) for both the left (β=−260.1, s.e.=86.2, t88=−3.02, P=0.003) and right (β=−219.4, s.e.=86.6, t88=−2.53, P=0.013) hippocampus. Also, there was a significant effect of group on the combined subfield volumes (Pillai’s trace=0.27, F14, 73=1.91, P=0.037) (Figure 2b). Post-hoc testing revealed that all of the subfields contributed to this overall main effect (Table 1c).
Longitudinal findings of Data set 1
In 41 controls (mean age 31.6±9.3 years) and 34 patients (mean age 30.9±9.1 years, of which 13 were in the first 5 years of illness), we tested whether there was an interaction effect between group and time for each subfield. In this subsample, only a subtle group difference in baseline volumes was seen in CA1 (β=−32.5, s.e.=14.54, P=0.043; uncorrected). Over time, the volumes of all subfields, except the left tail, decreased at a significantly greater rate in the schizophrenia cohort than in the healthy control cohort (Figure 3a). Specifically, subfields showing significant volume loss in the schizophrenia group (as indicated by the beta coefficients, as well as derived P-values that survived multiple comparisons) included the left CA1, right CA1, right ML, right CA2/3, left GCL and right GCL (Table 2a).

Longitudinal change in hippocampal subfield volumes in schizophrenia over time. (a) Spaghetti plots are shown indicating the trajectories of volumes of the hippocampal subfields, which showed a steeper rate of loss in patients compared with controls (in a subset of Data set 1). Bold lines indicate the group mean linear regression line. GCL, granule cell layer; ML, molecular layer; Sub, subiculum. (b) Also, in this cohort, the rate of atrophy of left CA1 was correlated with the rate of increasing negative symptom severity in the patients. The scatter plot showing the standardized rate of change of negative symptoms versus the rate of change of left CA1 volume across all schizophrenia patients (r=−0.54, P=0.0023) is displayed.
Secondary analyses
After adjusting for different medication classes, the CA1 volume deficit in Data set 1 remained significant (Supplementary Table 5A). Similarly, the extensive volume deficits across the hippocampal subfields of the more chronic patients of Data set 2 remained present, as well as the progressive volume decline across multiple subfields in the patients of the longitudinal cohort (Supplementary Table 5B). Also, the results of our primary analyses remained unchanged after adjusting for ethnicity.
Relationships of the findings to symptoms and illness duration
Across the two cross-sectional data sets, no significant correlations were found between symptom levels and absolute volume measures of all of the subfields, after correction for multiple comparisons. Also, no relationships between medication dosages and subfield volumes were found (Supplementary Table 6). However, negative associations between duration of illness and CA1 volumes were observed in both cohorts (Data set 1, left CA1: r=−0.22, P=0.006, right CA1: r=−0.21, P=0.01; Data set 2, left CA1: r=−0.27, P=0.065, right CA1: r=−0.32, P=0.03) (Supplementary Table 7).
Within the patient cohort of the longitudinal analysis, there were—on average—mild improvements in positive symptoms (P<0.012) and general psychopathology symptoms (P<0.06), but no significant changes in negative symptoms, over time (between baseline and follow-up). However, when we examined intra-individual relationships between symptoms and subfield volumes over time (adjusting for age, sex, antipsychotic dosages and baseline duration of illness), associations between rate of worsening of symptoms across all symptom domains and the rate of change in subfield volume were observed (Table 2b). Figure 3b displays a plot of the correlation between the rate of left CA1 atrophy and rate of increase in negative symptom severity (r=−0.54, P=0.0023).
Discussion
Here we demonstrated that MRI volume estimates of the CA1 of the hippocampus were selectively reduced in patients in the early (including those in the first 5 years of illness) to mid-course of schizophrenia, when compared with healthy controls. In contrast, in a cohort of chronic schizophrenia patients, the volume deficits were widespread across the subfields. Consistent with this pattern, the results of our longitudinal analysis indicated that over time, the focal atrophy associated with early illness extends beyond CA1, involving other subfields such as the CA2/3 and GCL. Finally, correlational analyses revealed that subfield volumes were (1) cross-sectionally smaller with greater illness duration and (2) declined with illness progression (that is, worsening symptoms) over time.
Early changes in CA1 in schizophrenia, with later involvement of other subfields
The pattern of disproportionate reductions in CA1 volume in schizophrenia patients of Data set 1 agrees with convergent evidence from shape morphometric studies of first-episode schizophrenia patients.29 The diffuse subfield volume deficits in the chronic patients of Data set 2 are consistent with prior evidence of widespread volume deficits along the anterior–posterior extent of the hippocampus in patients with long-term schizophrenia.15 Negative correlations between illness duration and subfield volumes were also found in both data sets, similar to prior reports of an association between hippocampal volume reduction and illness duration in chronic schizophrenia.12, 52, 53 Taken together, these cross-sectional findings raise the possibility that a progressive extension of atrophy across the hippocampus occurs during the illness. We tested this hypothesis by conducting a longitudinal analysis in a subset of subjects of Data set 1. Here we found subtle volume reductions at baseline in the left CA1—but not in the other subfields—in the schizophrenia patients when compared with the controls. Over time, significant effects of the illness were observed not only in the CA1 but also in other subfields that are part of the trisynaptic circuit, that is, CA2/3, ML and GCL.16, 54 The extent of atrophy averaged about 2–6% loss per year in the most affected subfields, with the greatest amount of volume change in the CA1 subregion (~6%). We found that the rate of atrophy across the subfields in schizophrenia correlated with the rate of symptom worsening over time. Thus, our findings suggest that the greatest decline in subfield volume over time may occur in patients who have a poor course of illness.
The present finding of an initial, selective loss of CA1 volume during the early stages of illness is consistent with a finding of increased CA1 regional cerebral blood volume in prodromal subjects who subsequently became ill, compared with those that did not.55 In this prior study, CA1 hypermetabolism predated changes in shape measures of CA1 and subicular atrophy in these subjects.55 The present finding is also consistent with prior findings of selective CA1 hypermetabolism or shape deformity in patients in the early-to-mid stages of illness, including patients with mean illness duration of ⩽10 years.29, 30, 56, 57
Our findings, however, are not in line with two recent studies, which found pronounced volume deficits in the CA2/3, CA4/DG and Sub subfields of the hippocampus in schizophrenia patients, with little or no differences in CA1.32, 33 The discrepancy between these prior findings and ours is likely due to differences in hippocampal subfield labeling methods. It has recently been shown that portions of CA1 are misattributed as CA2/3, CA4-DG and Sub by the older automated algorithm used in these prior studies.23, 36, 58 Indeed, our own calculations indicate that volume estimates45 of the CA1 using the prior algorithm are approximately 40% smaller than the current, ex vivo algorithm’s CA1 volume estimates, and that the previous volume estimates of the CA2/3 are four times larger than the ex vivo algorithm’s CA2/3 volume estimates.
Possible mechanisms underlying the selective-to-diffuse changes in the hippocampus in schizophrenia
Small et al.3 and Schoebel et al.55 have proposed a framework to explain an early involvement of CA1 in the pathophysiological process underlying psychosis. They suggest a sequence of events involving an increase in synaptic glutamate levels, subsequent increase in metabolic demand and blood flow, and downregulation of GABAergic interneurons in the hippocampus.55 Owing to the greater density of glutamate receptors (N-Methyl-D-aspartate receptor and α-Amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor) in CA1 compared with the other hippocampal subfields,59 CA1 may be particularly vulnerable to dysregulation of glutamatergic neurotransmission and excitotoxic injury.3 Moreover, as Konradi et al.60 have shown, CA1 has the largest numbers of primary hippocampal interneuron subtypes among the subfields, the loss of function of these interneurons, which control information flow arriving from the entorhinal cortex and excitatory hippocampal pyramidal cell activity,61 may produce excessive, feedforward excitation of the trisynaptic hippocampal circuit,54, 62 driving an extension of these abnormalities in excitation–inhibition balance to other portions of the hippocampus and beyond.61, 63, 64, 65, 66 Empirical support for this model has been produced using ketamine-treated mice, which showed hypermetabolism followed by atrophy selectively in CA1 and the Sub; these effects were subsequently blocked by reducing synaptic glutamate levels.55
The progression of atrophy from CA1 to other hippocampal subfields could result from an extension of the pathophysiological process underlying psychosis, such as the one hypothesized above. Consistent with this possibility are the findings of extensive reductions in interneuron subtypes throughout the hippocampus (in the CA1, CA2/3 and CA4 fields) in post-mortem studies of schizophrenia.60 This possibility is also supported by the association between progressive atrophy and symptomatic worsening observed in this study. Alternatively, this progression could result from an interaction between (1) a fundamental cellular abnormality and (2) environmental factors,61 including stress, substance use and/or treatment with antipsychotic medications, all of which have been associated with decreases in brain tissue volume.67, 68, 69 However, when we controlled for antipsychotic medication dosages (as well as the use of mood stabilizers and antidepressants) in the analyses, both our cross-sectional and longitudinal findings in both data sets remained significant, suggesting that progression of atrophy across the hippocampus over time in schizophrenia is not a consequence of medication treatment. Also, it is notable that our longitudinal data were collected in a country (Singapore) with extremely strict prohibitions against (and hence very little) illicit substance use. Thus, although substance use, cannabis in particular, has been linked to reductions in hippocampal volume in both healthy and schizophrenia groups,70, 71, 72 it is unlikely to have had a role in our longitudinal findings.
However, we speculate that elevated stress levels,68 as well as unhealthy lifestyle practices common in patients with chronic schizophrenia, such as poor nutrition, cigarette smoking73, 74 and lack of exercise,75, 76 exacerbate dysfunction and structural changes of the hippocampus in the illness. Future studies can quantitatively measure these environmental factors longitudinally to determine which influence progression of hippocampal atrophy in schizophrenia.
Intriguingly, a recent multisite structural MRI study of major depressive disorder show a pattern of results that is similar to those of this study, with hippocampal volume deficits in chronic patients, but no such deficit in first-episode depression subjects.77 Taken together with evidence for hippocampal hypermetabolism in depression,78, 79 these findings suggest that related, overlapping mechanisms may be responsible for hippocampal abnormalities in schizophrenia and depression, as recently proposed.3 The possibility that identical mechanisms, affecting overlapping but partially distinct circuits, underlie psychosis and depression, could account both for the overlap in symptoms (for example, anhedonia) as well as the phenomenological differences across the two disorders.
Potential limitations and summary
There are several potential limitations of our study. First, although the hippocampal segmentations of all subjects studied here were visually inspected individually, it is possible that schizophrenia disrupts the folding of the multi-layered hippocampus, influencing the accuracy of the hippocampal segmentation process. However, the current segmentation algorithm has been used, with a high level of accuracy, to delineate the subfields of the hippocampus in patients with Alzheimer’s disease—who exhibit greater cellular degradation and atrophy of the hippocampus than patients with schizophrenia,37 suggesting that this method is likely fairly robust to disease-induced alterations.
Second, it is possible that there may be differences in hippocampal structure across ethnic groups that may limit the validity of examining hippocampal subfields across cohorts with different ethnic compositions. However, prior studies, including one that directly compared the brain structures of 140 cognitively matched young and old Chinese Singaporeans and non-Asian Americans, have found no evidence of effects of ethnicity on hippocampal volume measures.80, 81 Hippocampal volume deficits are also consistently reported in studies of schizophrenia across patients of varying ethnicities;4, 7, 82, 83, 84, 85, 86 thus it is unlikely that the differences in ethnicity across the two sites of our study impacts the generalizability of these results.
In conclusion, this study reports selective volume deficits of the CA1 in the early-to-mid phases of schizophrenia, with evidence for an extension of this atrophy to the remaining hippocampal subfields over the course of the illness. Correlations between these findings and measures of illness progression suggest that these anatomical changes may have direct clinical consequences, which could be treated, in future trials, by interventions aimed at restoring or preserving the hippocampus.
References
Harrison PJ . The hippocampus in schizophrenia: a review of the neuropathological evidence and its pathophysiological implications. Psychopharmacology (Berl) 2004; 174: 151–162.
Tamminga CA, Stan AD, Wagner AD . The hippocampal formation in schizophrenia. Am J Psychiatry 2010; 167: 1178–1193.
Small SA, Schobel SA, Buxton RB, Witter MP, Barnes CA . A pathophysiological framework of hippocampal dysfunction in ageing and disease. Nat Rev Neurosci 2011; 12: 585–601.
Adriano F, Caltagirone C, Spalletta G . Hippocampal volume reduction in first-episode and chronic schizophrenia: a review and meta-analysis. Neuroscientist 2012; 18: 180–200.
Steen RG, Mull C, McClure R, Hamer RM, Lieberman JA . Brain volume in first-episode schizophrenia: systematic review and meta-analysis of magnetic resonance imaging studies. Br J Psychiatry 2006; 188: 510–518.
Vita A, De Peri L, Silenzi C, Dieci M . Brain morphology in first-episode schizophrenia: a meta-analysis of quantitative magnetic resonance imaging studies. Schizophr Res 2006; 82: 75–88.
van Erp TG, Hibar DP, Rasmussen JM, Glahn DC, Pearlson GD, Andreassen OA et al. Subcortical brain volume abnormalities in 2028 individuals with schizophrenia and 2540 healthy controls via the ENIGMA consortium. Mol Psychiatry 2015; e-pub ahead of print 2 June 2015; doi:10.1038/mp.2015.63.
Wood SJ, Velakoulis D, Smith DJ, Bond D, Stuart GW, McGorry PD et al. A longitudinal study of hippocampal volume in first episode psychosis and chronic schizophrenia. Schizophr Res 2001; 52: 37–46.
Whitworth AB, Kemmler G, Honeder M, Kremser C, Felber S, Hausmann A et al. Longitudinal volumetric MRI study in first- and multiple-episode male schizophrenia patients. Psychiatry Res 2005; 140: 225–237.
Lieberman J, Chakos M, Wu H, Alvir J, Hoffman E, Robinson D et al. Longitudinal study of brain morphology in first episode schizophrenia. Biol Psychiatry 2001; 49: 487–499.
Mamah D, Harms MP, Barch D, Styner M, Lieberman JA, Wang L . Hippocampal shape and volume changes with antipsychotics in early stage psychotic illness. Front Psychiatry 2012; 3: 96.
Velakoulis D, Wood SJ, Wong MT, McGorry PD, Yung A, Phillips L et al. Hippocampal and amygdala volumes according to psychosis stage and diagnosis: a magnetic resonance imaging study of chronic schizophrenia, first-episode psychosis, and ultra-high-risk individuals. Arch Gen Psychiatry 2006; 63: 139–149.
Ebdrup BH, Skimminge A, Rasmussen H, Aggernaes B, Oranje B, Lublin H et al. Progressive striatal and hippocampal volume loss in initially antipsychotic-naive, first-episode schizophrenia patients treated with quetiapine: relationship to dose and symptoms. Int J Neuropsychopharmacol 2011; 14: 69–82.
Rizos E, Papathanasiou MA, Michalopoulou PG, Laskos E, Mazioti A, Kastania A et al. A longitudinal study of alterations of hippocampal volumes and serum BDNF levels in association to atypical antipsychotics in a sample of first-episode patients with schizophrenia. PLoS One 2014; 9: e87997.
Weiss AP, Dewitt I, Goff D, Ditman T, Heckers S . Anterior and posterior hippocampal volumes in schizophrenia. Schizophr Res 2005; 73: 103–112.
Burwell RD, Agster KL . Anatomy of the Hippocampus and the Declarative Memory System. In: Eichenbaum HE (ed). Learning and Memory: A Comprehensive Reference, vol. 3. Elsevier: Oxford, 2008, pp 47–66.
Simons JS, Spiers HJ . Prefrontal and medial temporal lobe interactions in long-term memory. Nat Rev Neurosci 2003; 4: 637–648.
Bakker A, Kirwan CB, Miller M, Stark CE . Pattern separation in the human hippocampal CA3 and dentate gyrus. Science 2008; 319: 1640–1642.
Leutgeb JK, Leutgeb S, Moser MB, Moser EI . Pattern separation in the dentate gyrus and CA3 of the hippocampus. Science 2007; 315: 961–966.
McHugh TJ, Jones MW, Quinn JJ, Balthasar N, Coppari R, Elmquist JK et al. Dentate gyrus NMDA receptors mediate rapid pattern separation in the hippocampal network. Science 2007; 317: 94–99.
Rolls ET . A quantitative theory of the functions of the hippocampal CA3 network in memory. Front Cell Neurosci 2013; 7: 98.
Mankin EA, Sparks FT, Slayyeh B, Sutherland RJ, Leutgeb S, Leutgeb JK . Neuronal code for extended time in the hippocampus. Proc Natl Acad Sci USA 2012; 109: 19462–19467.
Bonnici HM, Chadwick MJ, Kumaran D, Hassabis D, Weiskopf N, Maguire EA . Multi-voxel pattern analysis in human hippocampal subfields. Front Hum Neurosci 2012; 6: 290.
Yushkevich PA, Wang H, Pluta J, Das SR, Craige C, Avants BB et al. Nearly automatic segmentation of hippocampal subfields in in vivo focal T2-weighted MRI. Neuroimage 2010; 53: 1208–1224.
Yassa MA, Stark CE . A quantitative evaluation of cross-participant registration techniques for MRI studies of the medial temporal lobe. Neuroimage 2009; 44: 319–327.
Lacy JW, Yassa MA, Stark SM, Muftuler LT, Stark CE . Distinct pattern separation related transfer functions in human CA3/dentate and CA1 revealed using high-resolution fMRI and variable mnemonic similarity. Learn Mem 2011; 18: 15–18.
Ekstrom AD, Bazih AJ, Suthana NA, Al-Hakim R, Ogura K, Zeineh M et al. Advances in high-resolution imaging and computational unfolding of the human hippocampus. Neuroimage 2009; 47: 42–49.
Winterburn JL, Pruessner JC, Chavez S, Schira MM, Lobaugh NJ, Voineskos AN et al. A novel in vivo atlas of human hippocampal subfields using high-resolution 3 T magnetic resonance imaging. Neuroimage 2013; 74: 254–265.
Narr KL, Thompson PM, Szeszko P, Robinson D, Jang S, Woods RP et al. Regional specificity of hippocampal volume reductions in first-episode schizophrenia. Neuroimage 2004; 21: 1563–1575.
Zierhut KC, Grassmann R, Kaufmann J, Steiner J, Bogerts B, Schiltz K . Hippocampal CA1 deformity is related to symptom severity and antipsychotic dosage in schizophrenia. Brain 2013; 136: 804–814.
Kuhn S, Musso F, Mobascher A, Warbrick T, Winterer G, Gallinat J . Hippocampal subfields predict positive symptoms in schizophrenia: first evidence from brain morphometry. Transl Psychiatry 2012; 2: e127.
Mathew I, Gardin TM, Tandon N, Eack S, Francis AN, Seidman LJ et al. Medial temporal lobe structures and hippocampal subfields in psychotic disorders: findings from the Bipolar-Schizophrenia Network on Intermediate Phenotypes (B-SNIP) Study. JAMA Psychiatry 2014; 71: 769–777.
Haukvik UK, Westlye LT, Morch-Johnsen L, Jorgensen KN, Lange EH, Dale AM et al. In vivo hippocampal subfield volumes in schizophrenia and bipolar disorder. Biol Psychiatry 2014; 77: 581–588.
Pluta J, Yushkevich P, Das S, Wolk D . In vivo analysis of hippocampal subfield atrophy in mild cognitive impairment via semi-automatic segmentation of T2-weighted MRI. J Alzheimer' Dis 2012; 31: 85–99.
Iglesias JE AJ, Nguyen K, Player CM, Player A, Wright M, Roy N et al. A computational atlas of the hippocampal formation using ex vivo, ultra-high resolution MRI: application to adaptive segmentation of in vivo MRI. Neuroimage 2015; 115: 117–137.
Wisse LE, Biessels GJ, Geerlings MI . A critical appraisal of the hippocampal subfield segmentation package in FreeSurfer. Front Aging Neurosci 2014; 6: 261.
Iglesias JE, Augustinack JC, Nguyen K, Player CM, Player A, Wright M et al. A computational atlas of the hippocampal formation using ex vivo, ultra-high resolution MRI: application to adaptive segmentation of in vivo MRI. Neuroimage 2015; 115: 117–137.
First MBSR, Gibbon M, Williams JBW . Structured Clinical Interview for DSM-IV-TR Axis I Disorders, Research Version, Patient Edition With Psychotic Screen Biometrics Research, New York State Psychiatric Institute: New York, 2002.
Kay SR, Fiszbein A, Opler LA . The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophr Bull 1987; 13: 261–276.
Fischl B, Salat DH, van der Kouwe AJ, Makris N, Segonne F, Quinn BT et al. Sequence-independent segmentation of magnetic resonance images. Neuroimage 2004; 23: S69–S84.
Dale AM, Fischl B, Sereno MI . Cortical surface-based analysis. I. Segmentation and surface reconstruction. Neuroimage 1999; 9: 179–194.
Fischl B, Salat DH, Busa E, Albert M, Dieterich M, Haselgrove C et al. Whole brain segmentation: automated labeling of neuroanatomical structures in the human brain. Neuron 2002; 33: 341–355.
Fischl B, Sereno MI, Dale AM . Cortical surface-based analysis. II: inflation, flattening, and a surface-based coordinate system. Neuroimage 1999; 9: 195–207.
Reuter M, Schmansky NJ, Rosas HD, Fischl B . Within-subject template estimation for unbiased longitudinal image analysis. Neuroimage 2012; 61: 1402–1418.
Van Leemput K, Bakkour A, Benner T, Wiggins G, Wald LL, Augustinack J et al. Automated segmentation of hippocampal subfields from ultra-high resolution in vivo MRI. Hippocampus 2009; 19: 549–557.
The R Core Team. R: A language and environment for statistical computing. R foundation for statistical computing. Vienna, Austria, 2014.
van Haren NE, Hulshoff Pol HE, Schnack HG, Cahn W, Brans R, Carati I et al. Progressive brain volume loss in schizophrenia over the course of the illness: evidence of maturational abnormalities in early adulthood. Biol Psychiatry 2008; 63: 106–113.
Mattai A, Hosanagar A, Weisinger B, Greenstein D, Stidd R, Clasen L et al. Hippocampal volume development in healthy siblings of childhood-onset schizophrenia patients. Am J Psychiatry 2011; 168: 427–435.
Frodl T, Jager M, Smajstrlova I, Born C, Bottlender R, Palladino T et al. Effect of hippocampal and amygdala volumes on clinical outcomes in major depression: a 3-year prospective magnetic resonance imaging study. J Psychiatry Neurosci 2008; 33: 423–430.
Yucel K, Taylor VH, McKinnon MC, Macdonald K, Alda M, Young LT et al. Bilateral hippocampal volume increase in patients with bipolar disorder and short-term lithium treatment. Neuropsychopharmacology 2008; 33: 361–367.
Holm S . A simple sequentially rejective multiple test procedure. Scand J Stat 1979; 6: 65–70.
Velakoulis D, Pantelis C, McGorry PD, Dudgeon P, Brewer W, Cook M et al. Hippocampal volume in first-episode psychoses and chronic schizophrenia: a high-resolution magnetic resonance imaging study. Arch Gen Psychiatry 1999; 56: 133–141.
Chakos MH, Schobel SA, Gu H, Gerig G, Bradford D, Charles C et al. Duration of illness and treatment effects on hippocampal volume in male patients with schizophrenia. Br J Psychiatry 2005; 186: 26–31.
Benes FM . Evidence for altered trisynaptic circuitry in schizophrenic hippocampus. Biol Psychiatry 1999; 46: 589–599.
Schobel SA, Chaudhury NH, Khan UA, Paniagua B, Styner MA, Asllani I et al. Imaging patients with psychosis and a mouse model establishes a spreading pattern of hippocampal dysfunction and implicates glutamate as a driver. Neuron 2013; 78: 81–93.
Schobel SA, Lewandowski NM, Corcoran CM, Moore H, Brown T, Malaspina D et al. Differential targeting of the CA1 subfield of the hippocampal formation by schizophrenia and related psychotic disorders. Arch Gen Psychiatry 2009; 66: 938–946.
Talati P, Rane S, Kose S, Blackford JU, Gore J, Donahue MJ et al. Increased hippocampal CA1 cerebral blood volume in schizophrenia. Neuroimage Clin 2014; 5: 359–364.
Schoene-Bake JC, Keller SS, Niehusmann P, Volmering E, Elger C, Deppe M et al. In vivo mapping of hippocampal subfields in mesial temporal lobe epilepsy: relation to histopathology. Hum Brain Map 2014; 35: 4718–4728.
Coultrap SJ, Nixon KM, Alvestad RM, Valenzuela CF, Browning MD . Differential expression of NMDA receptor subunits and splice variants among the CA1, CA3 and dentate gyrus of the adult rat. Brain Res Mol Brain Res 2005; 135: 104–111.
Konradi C, Yang CK, Zimmerman EI, Lohmann KM, Gresch P, Pantazopoulos H et al. Hippocampal interneurons are abnormal in schizophrenia. Schizophr Res 2011; 131: 165–173.
Heckers S, Konradi C . GABAergic mechanisms of hippocampal hyperactivity in schizophrenia. Schizophr Res 2015; 167: 4–11.
Benes FM, Berretta S . GABAergic interneurons: implications for understanding schizophrenia and bipolar disorder. Neuropsychopharmacology 2001; 25: 1–27.
Heckers S, Konradi C . Hippocampal pathology in schizophrenia. Curr Topics Behav Neurosci 2010; 4: 529–553.
Lisman JE, Coyle JT, Green RW, Javitt DC, Benes FM, Heckers S et al. Circuit-based framework for understanding neurotransmitter and risk gene interactions in schizophrenia. Trends Neurosci 2008; 31: 234–242.
Moghaddam B, Javitt D . From revolution to evolution: the glutamate hypothesis of schizophrenia and its implication for treatment. Neuropsychopharmacology 2012; 37: 4–15.
Lodge DJ, Grace AA . Hippocampal dysregulation of dopamine system function and the pathophysiology of schizophrenia. Trends Pharmacol Sci 2011; 32: 507–513.
Fusar-Poli P, Smieskova R, Kempton MJ, Ho BC, Andreasen NC, Borgwardt S . Progressive brain changes in schizophrenia related to antipsychotic treatment? A meta-analysis of longitudinal MRI studies. Neurosci Biobehav Rev 2013; 37: 1680–1691.
Mondelli V, Cattaneo A, Belvederi Murri M, Di Forti M, Handley R, Hepgul N et al. Stress and inflammation reduce brain-derived neurotrophic factor expression in first-episode psychosis: a pathway to smaller hippocampal volume. J Clin Psychiatry 2011; 72: 1677–1684.
Schneider CE, White T, Hass J, Geisler D, Wallace SR, Roessner V et al. Smoking status as a potential confounder in the study of brain structure in schizophrenia. J Psychiatr Res 2014; 50: 84–91.
Smith MJ, Cobia DJ, Reilly JL, Gilman JM, Roberts AG, Alpert KI et al. Cannabis-related episodic memory deficits and hippocampal morphological differences in healthy individuals and schizophrenia subjects. Hippocampus 2015; 25: 1042–1051.
Solowij N, Walterfang M, Lubman DI, Whittle S, Lorenzetti V, Styner M et al. Alteration to hippocampal shape in cannabis users with and without schizophrenia. Schizophr Res 2013; 143: 179–184.
Lorenzetti V, Solowij N, Whittle S, Fornito A, Lubman DI, Pantelis C et al. Gross morphological brain changes with chronic, heavy cannabis use. Br J Psychiatry 2015; 206: 77–78.
Goff DC, Cather C, Evins AE, Henderson DC, Freudenreich O, Copeland PM et al. Medical morbidity and mortality in schizophrenia: guidelines for psychiatrists. J Clin Psychiatry 2005; 66: 183–194, quiz 147, 273-184.
Jiang J, See YM, Subramaniam M, Lee J . Investigation of cigarette smoking among male schizophrenia patients. PLoS One 2013; 8: e71343.
Brambilla P, Perlini C, Rajagopalan P, Saharan P, Rambaldelli G, Bellani M et al. Schizophrenia severity, social functioning and hippocampal neuroanatomy: three-dimensional mapping study. Br J Psychiatry 2013; 202: 50–55.
Zipursky RB, Reilly TJ, Murray RM . The myth of schizophrenia as a progressive brain disease. Schizophr Bull 2013; 39: 1363–1372.
Schmaal L, Veltman DJ, van Erp TG, Samann PG, Frodl T, Jahanshad N et al. Subcortical brain alterations in major depressive disorder: findings from the ENIGMA Major Depressive Disorder working group. Mol Psychiatry 2015; e-pub ahead of print 30 June 2015; doi:10.1038/mp.2015.69.
Lui S, Parkes LM, Huang X, Zou K, Chan RC, Yang H et al. Depressive disorders: focally altered cerebral perfusion measured with arterial spin-labeling MR imaging. Radiology 2009; 251: 476–484.
Videbech P, Ravnkilde B, Pedersen TH, Hartvig H, Egander A, Clemmensen K et al. The Danish PET/depression project: clinical symptoms and cerebral blood flow. A regions-of-interest analysis. Acta Psychiatr Scand 2002; 106: 35–44.
Brickman AM, Schupf N, Manly JJ, Luchsinger JA, Andrews H, Tang MX et al. Brain morphology in older African Americans, Caribbean Hispanics, and whites from northern Manhattan. Arch Neurol 2008; 65: 1053–1061.
Chee MW, Zheng H, Goh JO, Park D, Sutton BP . Brain structure in young and old East Asians and Westerners: comparisons of structural volume and cortical thickness. J Cogn Neurosci 2011; 23: 1065–1079.
Yang Y, Raine A, Han CB, Schug RA, Toga AW, Narr KL . Reduced hippocampal and parahippocampal volumes in murderers with schizophrenia. Psychiatry Res 2010; 182: 9–13.
Kalmady SV, Venkatasubramanian G, Shivakumar V, Gautham S, Subramaniam A, Jose DA et al. Relationship between Interleukin-6 gene polymorphism and hippocampal volume in antipsychotic-naive schizophrenia: evidence for differential susceptibility? PLoS One 2014; 9: e96021.
Kim SH, Lee JM, Kim HP, Jang DP, Shin YW, Ha TH et al. Asymmetry analysis of deformable hippocampal model using the principal component in schizophrenia. Hum Brain Map 2005; 25: 361–369.
Yamasue H, Iwanami A, Hirayasu Y, Yamada H, Abe O, Kuroki N et al. Localized volume reduction in prefrontal, temporolimbic, and paralimbic regions in schizophrenia: an MRI parcellation study. Psychiatry Res 2004; 131: 195–207.
Verma S, Sitoh YY, Ho YC, Poon LY, Subramaniam M, Chan YH et al. Hippocampal volumes in first-episode psychosis. J Neuropsychiatry Clin Neurosci 2009; 21: 24–29.
Acknowledgements
This study was supported by the National Medical Research Council under the Centre Grant Programme (Institute of Mental Health, Singapore) (NMRC/CG/004/2013) (NFH) and by the National Healthcare Group, Singapore (SIG/05004; SIG/05028), and the Singapore Bioimaging Consortium (RP C-009/2006) research grants awarded to KS, as well grants from the National Institute of Mental Health (DJH: K23MH076054; JLR: K23MH084059) and a Howard Hughes Medical Institute Physician Scientist Early Career Award and grant from Pamlab (JLR). This research was also supported by the Biomedical Research Council, Singapore (awarded to JZ, BMRC 04/1/36/372), the Agency for Science, Technology, and Research (A*STAR) and Duke-NUS Graduate Medical School Signature Research Program funded by Ministry of Health, Singapore. We thank Dr. Jean Augustinack for her valuable comments on the manuscript. We also thank Garth Coombs III, Alexandra S. Tanner, Adam Z. Nitenson, and Liana J. Petruzzi for their indispensable help with the MRI and clinical data acquisition, as well as Stephanie N. DeCross and Franklin C. Hungtington for their administrative support.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on the Molecular Psychiatry website
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Ho, N., Iglesias, J., Sum, M. et al. Progression from selective to general involvement of hippocampal subfields in schizophrenia. Mol Psychiatry 22, 142–152 (2017). https://doi.org/10.1038/mp.2016.4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mp.2016.4
This article is cited by
-
Smaller anterior hippocampal subfields in the early stage of psychosis
Translational Psychiatry (2024)
-
Changes in responses of the amygdala and hippocampus during fear conditioning are associated with persecutory beliefs
Scientific Reports (2024)
-
Longitudinal hippocampal subfield development associated with psychotic experiences in young people
Translational Psychiatry (2024)
-
Longitudinal study on hippocampal subfields and glucose metabolism in early psychosis
Schizophrenia (2024)
-
Effects of web-based adapted physical activity on hippocampal plasticity, cardiorespiratory fitness, symptoms, and cardiometabolic markers in patients with schizophrenia: a randomized, controlled study
European Archives of Psychiatry and Clinical Neuroscience (2024)
