
Abstract
Isolated hypodontia, or congenital absence of one to six permanent teeth (OMIM 300606), is a common condition that affects about 20% of individuals worldwide. We identified two extended Pakistani pedigrees segregating X-linked hypodontia with variable expressivity. Affected males show no other associated anomalies, and obligate carrier females have normal dentition. We analyzed the families with polymorphic markers in the ectodysplasin A (EDA) gene region and obtained significant linkage to the phenotype in each pedigree (Zmax 3.29 and 2.65, respectively, at Ө = 0.00). Sequence analysis of the coding regions of EDA revealed a novel missense mutation c.1091T>C resulting in a methionine to threonine substitution (p.M364T) in the tumor necrosis factor (TNF) homology domain. Met364 is a highly conserved residue located on the outer surface of the EDA protein. From our findings, we suggest that the mutation disturbs but does not destroy the EDA structure, resulting in the partial and unusually mild ED phenotype restricted to hypodontia.
Similar content being viewed by others
Introduction
Hypodontia is defined as the congenital lack of between one and six teeth, which is a common anomaly throughout the world. It is estimated that about 20% of humans are affected by the isolated absence of one or several permanent teeth (Vastardis 2000). Hypodontia may be inherited in an autosomal recessive, autosomal dominant, X-linked dominant, or X-linked recessive mode. The most commonly missing permanent teeth are the third molars (20%), second premolars (3.4%), and maxillary lateral incisors (2.2%) (Simons et al. 1993). Four genes underlying isolated or nonsyndromic hypodontia have been identified so far: PAX9 (Stockton et al. 2000), MSX1 (Vastardis et al. 1996), AXIN2 (Mostowska et al. 2006), and EDA (Tao et al. 2006). Mutations in the EDA gene are usually associated with syndromic hypodontia involving other ectodermal tissues, but two recent reports indicate that certain EDA mutations may cause isolated hypodontia (Tao et al. 2006; Tarpey et al. 2007).
We studied two unrelated extended families with an X-linked recessive form of hypodontia. Affected individuals from both families presented with otherwise normal ectodermal structures, including nails, body hair, and facial features, and they reported normal sweating. Hypodontia affects mainly incisors and to some extent canines and premolars with a variable expression. All female carriers investigated are nonpenetrant.
Materials and methods
Subjects
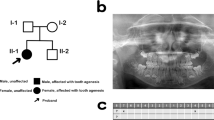
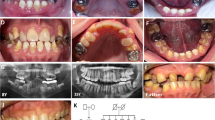
Two unrelated large consanguineous families from geographically distinct areas of Punjab, Pakistan, were ascertained to have hypodontia after clinical examination (Fig. 1a). Analysis of the pedigrees suggested an X-linked recessive mode of inheritance in both families. Altogether, nine affected males were identified in both families, whereas five obligate carrier females appeared asymptomatic. All living normal and affected individuals underwent medical and oral examination at the Nishtar Hospital, Multan, Pakistan. The most striking feature in affected individuals of both families was the congenital absence of at least one mandibular and/or one maxillary incisor in both primary and permanent dentition. In two cases, all eight incisors were missing. There were no visible teeth malformations in affected males. The body and scalp hair, skin, and nails were normal, and the males reported normal sweating and heat tolerance.

a Pedigrees of family A and family B segregating X-linked recessive incisor hypodontia. Affected males are indicated by filled squares, and carrier females are identified by circles with a dot in the center. b Schematic representation of congenitally missing teeth in available affected individuals showing the predominant incisor hypodontia phenotype. c Panoramic X-ray of a 14-year-old affected male (family B, ind. IV:9). Arrows indicate positions of missing teeth. d Deoxyribonucleic acid (DNA) sequence chromatograms from EDA exon 9 showing the different genotypes at position c.1091 (arrows) in three individuals from family B. A healthy male (ind. IV:3; upper panel), a female carrier (ind. IV:8; mid panel), and an affected male (ind. IV:9; lower panel)
Genetic analysis and EDA modelling
DNA was extracted from peripheral blood samples obtained from nine affected and 17 healthy family members. Genetic linkage analysis was performed with the polymorphic markers DXS8040, DXS453, DXS8107, and DXS8052 spanning the EDA gene locus. Two-point log of odds (LOD) score was calculated using the MLINK program of FASTLINK computer package (Cottingham et al. 1993). Sequencing was performed using a BigDye Terminator v3.1 Cycle Sequencing Kit (Invitrogen, San Diego, CA, USA) on an ABI PRISM 3700 sequencer (Applied Biosystems). Chromatograms were analyzed using Sequencher v.4.1.2 (Gene Codes Corporation). Primers and sequencing conditions have been described previously (Visinoni et al. 2003). Informed consent was obtained from all individuals who participated in this study. Modeling of the EDA 3D structure was performed using software ViewerLite v5.0 and PDB file 1RJ7 (Hymowitz et al. 2003).
Results
Hypodontia and EDA missense mutation
An X-linked recessive inheritance for a phenotype restricted to the absence of three to 12 teeth was evident in both pedigrees. Obligate carrier females had normal dentition and normally shaped teeth. Most strikingly, all affected males showed a reduced number or a total absence of incisors (Fig. 1b, c). Dentition of premolars and molars was less affected. Linkage to the EDA gene region was confirmed using microsatellite markers. A significant LOD score was obtained for marker locus DXS8052 in the two families (Zmax 3.29 and 2.65, respectively, at Ө = 0.00; Table 1). An identical allele size for DXS8052 segregated hypodontia in both families, which suggested a shared ancestral mutation for a single EDA mutation. Sequencing of the eight coding exons and exon–intron boundaries of EDA on genomic DNA from all affected individuals revealed a novel c.1091T>C transition in exon 9 (Fig. 1d). This mutation results in p.M364T located in the highly conserved tumor necrosis factor (TNF) domain of EDA (Fig. 2a). The five women with normal dentition and affected male offspring were all carriers of this mutation. In addition, three healthy female family members without affected offspring were identified as carriers. The C allele in exon 9 was excluded on 200 unrelated Pakistani control chromosomes.

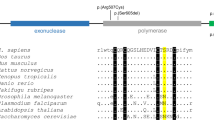
a Multiple species sequence alignment of known ectodysplasin (EDA)-A1 sequences showing a high evolutionary conservation around position M364. Sequences were obtained from ENSEMBL and analyzed using BioEdit v7.5.0.3. b, c Location of the Y320, Q364, and M364 residues within the tumor necrosis factor (TNF)-domain of EDA-A1 in the tertiary structure of the monomer (b, side view) and the quaternary structure of the homotrimer (c, view from top). A Y320C mutation was previously found implicated in the full ED syndrome, whereas a Q358E mutation was associated with isolated hypodontia. d Zoom-in of the TNF domain, indicating the M364 residue with neighboring amino acids. Pictures were generated using ViewerLite v5.0 and PDB file 1RJ7 (Hymowitz et al. 2003)
Position of Met364 in EDA 3D model
Modeling of the EDA 3D structure using ViewerLite v5.0 and PDB file 1RJ7 suggested the Met364 residue is localized on the outer surface of the crystal structure (Fig. 2b, c). The position excludes Met364 from being part of the interacting surface between EDA monomers. The residue occupies a space in immediate proximity to the side chains Ile260, Leu266, and Leu271 and relatively close to the Tyr320 residue (Fig. 2d). Thus, the hydrophobic interactions between Met364 and the nonpolar and hydrophobic side chains of Ile260, Leu266, and Leu271 suggest they are important to maintain the integrity and structure of the EDA monomer.
Discussion
Our results confirm that certain EDA mutations may be associated with isolated hypodontia. Mutations in EDA are known to cause ectodermal dysplasia (OMIM 305100), a clinical condition characterized by hypotrichosis (sparseness of scalp and body hair), hypohidrosis (reduced ability to sweat), and hypodontia (congenital absence of teeth). More than 64 mutations in the EDA gene have been identified in association with ED (Visinoni et al. 2003), but EDA is rarely reported as being implicated in isolated tooth agenesis (Tao et al. 2006; Tarpey et al. 2007). The EDA is a type II transmembrane protein containing a small N-terminal intracellular part and a larger C-terminal extracellular part. The extracellular part consists of a collagen-like domain and a TNF-like domain. The TNF domain has been shown to form homotrimers that are important for interaction with EDA receptors (EDAR; Hymowitz et al. 2003).
The Met364 residue is a highly conserved amino acid found in a series of known EDA proteins (Fig. 2a). M364 localizes to the outer surface of the crystal structure at the bottom of β–strand G (Hymowitz et al. 2003; Fig. 2b). The residue does not directly interfere with the surface between monomers, and it is therefore not localized within the receptor-binding site (Fig. 2c). However, the side chain occupies a space in close proximity to the side chains Ile260, Leu266, and Leu271 (Fig. 2d) and points toward amino acids Glu308 and Val309, both of which have several interactions in the EDA structure (Hymowitz et al. 2003). It is also positioned in close proximity to Tyr320 that if mutated to Y320C, results in an overall disruption of the EDA structure (Hymowitz et al. 2003; Schneider et al. 2001; Fig. 2b, d). The M364T substitution replaces a long nonpolar methionine with a bulkier and shorter polar threonine. This substitution is expected to affect the hydrophobic interactions with the neighboring nonpolar and hydrophobic side chains of Ile260, Leu266, and Leu271. This assumption is consistent with the homology structure modeling, which indicates a loose structure at one end of the molecule (Arnold et al. 2006). Mutations in the EDA protein may cause impaired function by either altering the overall structure, decreased transmembrane trafficking and proprotein cleavage, or by direct inhibition of the interaction with the cognate EDAR. According to the previous report of a Q358E mutation associated with hypodontia (Tarpey et al. 2007), we hypothesize that the M364T substitution partially interferes with the overall structure of EDA. This may lead to reduced stability for the EDA homotrimer and/or reduced affinity for their target receptors. A reduced but not disrupted homotrimerization could also explain the partial phenotype restricted to dentition observed in our families. Carrier females with one normal allele may then have the ability to compensate for the mutant EDA allele. Further functional characterization and crystal structure of mutant EDA protein is now required to clarify the effect of this and other mutations associated with hypodontia.
References
Arnold K, Bordoli L, Kopp J, Schwede T (2006) The SWISS-MODEL workspace: a web-based environment for protein structure homology modeling. Bioinformatics 22:195–201
Broman KW, Murray JC, Scheffield VC, White RL, Weber JL (1998) Comprehensive human genetic maps: individual and sex-specific variation in recombination. Am J Hum Genet 63:861–869
Cottingham R, Indury RM, Schaffer AA (1993) Faster sequential genetic linkage, computations. Am J Hum Genet 53:252–263
Hymowitz SG, Compaan DM, Yan M, Wallweber HJA, Dixit VM, Starovasnik MA, de Vos AM (2003) The crystal structures of EDA-A1 and EDA-A2: Splice variants with distinct receptor specificity. Structure 11:1513–1520
International Human Genome Sequence Consortium (2001) Initial sequence and analysis of the human genome. Nature 409:860–891
Mostowska A, Biedziak B, Jagodzinski P (2006) Axis inhibition protein 2 (AXIN2) polymorphisms may be a risk factor for selective tooth agenesis. J Hum Genet 51:262–266
Schneider P, Street SL, Gaide O, Hertig S, Tardivel A, Tschopp J, Runkel L, Alevizopoulos K, Ferguson BM, Zonana J (2001) Mutations leading to X-linked hypohidrotic ectodermal dysplasia affect three major functional domains in the tumor necrosis factor family member ectodysplasin-A. J Biol Chem 276:18819–18827
Simons AL, Stritzel F, Stamatiou J (1993) Anomalies associated with hypodontia of the permanent lateral incisors and second premolars. J Clin Pediatr Dent 17:109–111
Stockton DW, Das P, Goldenberg M, D’Souza RN, Patel PI (2000) Mutation of PAX9 is associated with oligodontia. Nat Genet 24:18–19
Tao R, Jin B, Guo SZ, Qing W, Feng GY, Brooks DG, Liu L, Xu J, Taiwei Li, Yujuan Yan, He L (2006) A novel missense mutation of the EDA gene in a Mongolian family with congenital Hypodontia. J Hum Genet 51:498–502
Tarpey P, Pemberton TJ, Stockton DW, Das P, Ninis V, Edkins S, Futreal PA, Wooster R, Kamath S, Rabindra N, Stratton MR, Patel PI (2007) A Novel Gln358Glu mutation in ectodysplasin A associated with X-linked dominant incisor hypodontia. Am J Med Genet Part A 143A:390–394
Vastardis H (2000) The genetics of human tooth agenesis: New discoveries for understanding dental anomalies. Am J Orthod Dentofacial Orthop 117:650–656
Vastardis H, Karimbux N, Guthua SW, Seidman JG, Seidman CE (1996) A human MSX1 homeodomain missense mutation causes selective tooth agenesis. Nat Genet 13:417–421
Visinoni AF, de Souza RL, Freire-Maia N, Gollop TR, Chautard-Freire-Maia EA (2003) X-linked hypohidrotic ectodermal dysplasia mutations in Brazilian families. Am J Med Genet 122A:51–55
Acknowledgments
We thank the families and patients that participated in this study. This study was supported by The Swedish Research Council, The Indevelops Fund, Thuréus Fund, Torsten and Ragnar Söderbergs Foundation, Uppsala University, and the Uppsala University Hospital. MR is supported by a Ph.D. fellowship from Higher Education Commission (HEC), Islamabad, Pakistan.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Rasool, M., Schuster, J., Aslam, M. et al. A novel missense mutation in the EDA gene associated with X-linked recessive isolated hypodontia. J Hum Genet 53, 894–898 (2008). https://doi.org/10.1007/s10038-008-0323-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10038-008-0323-x
Keywords
This article is cited by
-
Nonsyndromic oligodontia
Journal of Orofacial Orthopedics / Fortschritte der Kieferorthopädie (2017)
-
DNA methylation is critical for tooth agenesis: implications for sporadic non-syndromic anodontia and hypodontia
Scientific Reports (2016)
-
Novel missense mutation in the EDA gene in a family affected by oligodontia
Journal of Orofacial Orthopedics / Fortschritte der Kieferorthopädie (2016)
-
Genetic background of nonsyndromic oligodontia: a systematic review and meta-analysis
Journal of Orofacial Orthopedics / Fortschritte der Kieferorthopädie (2013)
-
Functional analysis of Ectodysplasin-A mutations causing selective tooth agenesis
European Journal of Human Genetics (2010)
