
Abstract
Because stromal cells can regulate the growth and metastasis of tumor cells, a compound that modulates the interaction between the stromal cells and the tumor cells can control the tumor progression. In the course of our screening for such a compound, we have isolated a new compound, intervenolin, from the culture broth of Nocardia sp. ML96-86F2. Intervenolin inhibits the growth of human gastric and colorectal cancer cell lines in the coculture with the respective organ-derived stromal cells more strongly than that of the cancer cells cultured alone. Intervenolin shows antitumor effect against a xenograft model of human colorectal cancer cells in vivo. Furthermore, intervenolin exerts selective anti-Helicobacter pylori effect.
Similar content being viewed by others

Introduction
Tumor tissues are composed of not only tumor cells but also surrounding stroma.1 The stroma consists of various normal cells such as endothelial cells and fibroblast-like cells (we call them merely stromal cells) and extracellular matrix.2 Among them, the stromal cells can regulate the growth of tumor cells positively or negatively through the interaction between the stromal cells and the tumor cells.2, 3, 4, 5, 6, 7 The interaction is mediated by the secreted factors or direct adhesion between them.
We have been studying the possibility that the modulation of the tumor–stromal cell interactions can be a new antitumor strategy.8, 9 In general, classical antitumor drugs directly act on tumor cells. Because genetic background of the tumor cells is heterogeneous, problems of sensitivity and resistance to such drugs always arise. On the contrary, stromal cells are normal cells with rather stable genetic background. Thus, targeting the stromal cells for an antitumor strategy could not have such problems. We therefore have been searching compounds that modulate the tumor–stromal cell interactions using human prostate cancer as a starting model. As a result, we have discovered some natural compounds modulating the tumor–stromal cell interactions of prostate cancer.10, 11, 12, 13 All of them show strong growth inhibitory activity against human prostate cancer cells when cocultured with human prostate stromal cells,10, 11, 12, 13 while general cytotoxic compounds such as adriamycin, vinblastine, and cisplatin do not.8, 9 Among them, leucinostatins and NBRI16716s show potent antitumor activity against a mouse xenograft model of human prostate cancer cells.12, 13 These results have made us apply our strategy to other cancer.
In this study, we have applied our strategy to human gastric and colorectal cancer and discovered a new antitumor compound, intervenolin, by screening among natural resources. We report here the isolation, structure determination and biological activities of intervenolin.
Results
Taxonomy of the producing strain ML96-86F2
The strain ML96-86F2 formed well-branched and slightly fragmented substrate mycelia. This strain formed straight to flexuous aerial mycelia with hook or spiral (Figure 1). Mature spore chains were moderately short but usually contained 3 to 10 or more spores per chain. The spore was oval with smooth surface and 0.5 × 0.7–0.9 × 1.2 μm in size. The substrate mycelia were pale yellow to pale yellowish brown. The aerial mycelia were white to pinkish white. These characteristics were observed on oatmeal agar (ISP medium No. 3), inorganic salts-starch agar (ISP medium No. 4), glycerol-asparagine agar (ISP medium No. 5), tyrosine agar (ISP medium No. 7) and sucrose-nitrate agar. Faint reddish to brownish soluble pigment was produced in ISP medium No. 5 and No. 7. The type of diaminopimelic acid isomers in whole-cell hydrolysates of the strain ML96-86F2 was determined to be the meso-form. Mycolic acids were detected. The partial 16S ribosomal RNA gene sequence (1431 bp) was determined. The GenBank/EMBL/DDBJ accession number for the 16S rRNA gene sequence of ML96-86F2 is AB774456. The strain showed high similarity with those of the genus Nocardia such as Nocardia anaemiae (IFM 0323T, 1415/1431 bp, T: Type strain, 98.8%), N. pseudovaccinii (DSM 43406T, 1410/1433 bp, 98.3%) and N. vinacea (MK703-102F1T, 1406/1431 bp, 98.3%). These phenotypic and genotypic data suggested that the strain ML96-86F2 belongs to genus Nocardia. Therefore, the strain was tentatively designated as Nocardia sp. ML96-86F2.

Scanning electron micrograph of ML96-86F2. Sucrose-nitrate agar, 30 °C, 14 days. Bar=1 μm
Isolation procedure for intervenolin
The culture broth of ML96-86F2 (10 l) was filtered and the mycelium was extracted with MeOH. After removing the mycelium by filtration, the extract was concentrated in vacuo to remove MeOH and diluted with H2O up to 1 l. The aqueous solution was extracted with EtOAc and the organic layer was concentrated in vacuo to give a crude material (1 g). The crude material was applied on a silica gel column (50 g, Wakogel C-200, 75–150 μm; Wako Pure Chemical Industries, Osaka, Japan) prepared with hexane-EtOAc (1:1), and eluted with hexane-EtOAc (1:1), hexane-EtOAc (1:3), and MeOH. The fractions eluted with hexane-EtOAc (1:1) were concentrated in vacuo to give 58.3 mg of crude intervenolin. The crude sample was purified by a reversed-phase HPLC column (Capcell Pak C18 UG120, 5 μm, 20 × 250 mm, 6.0 ml min−1; Shiseido, Tokyo, Japan) with 80% MeOH to afford pure 3.9 mg of intervenolin. The fractions eluted with hexane-EtOAc (1:3) were concentrated in vacuo to give 47.6 mg of crude sample and further purified by a reversed-phase HPLC column with 70% MeOH to afford pure 3.2 mg of CJ-13,13617 and 4.9 mg of CJ-13,217.17
Physico-chemical properties
The physico-chemical properties of intervenolin were summarized in Table 1. Intervenolin was isolated as yellowish oil and soluble in MeOH and DMSO. The molecular formula of intervenolin was determined to be C24H32N2OS2 by HRESI-MS. The general features of its UV and NMR spectra were similar to those of CJ-13,136 and CJ-13,217.17
Structure determination
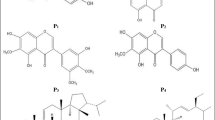
Although 1H- and 13C-NMR spectra of intervenolin (Table 2) were very similar to those of CJ-13,217,17 the signals of two methyl carbons of C-14 (δ 14.8) and C-16 (δ 15.1), a quaternary carbon of C-12 (δ 161.2), and a methylene carbon of C-10 (δ 63.9) instead of an N-methyl carbon were observed only in the 13C-NMR spectrum of intervenolin. The common part of the structure was confirmed by the 1H–1H COSY and 1H–13C HMBC spectra as shown in Figure 2. The remaining part of the structure was determined by 1H–13C HMBC and 1H–15N HMBC spectra. In the 1H–13C HMBC spectrum, methylene protons of 10-H (δ 5.55) correlated to three quaternary carbons of C-2, C-8a and C-12 and two singlet methyl protons of 14-H (δ 2.31) and 16-H (δ 2.70) correlated to a quaternary carbon of C-12 (Figure 2). Furthermore, in the 1H–15N HMBC spectrum, a methine proton of 8-H (δ 7.28) and methylene protons of 1′-H (δ 3.54) correlated to N-1 (δ 132) and methylene protons of 10-H (δ 5.55) correlated to N-11 (δ 282) as shown in Figure 3. Thus, the total structure of intervenolin was elucidated as shown in Figure 4.

1H–1H COSY and 1H–13C HMBC correlations in intervenolin.

1H–15N HMBC correlations in intervenolin.

Structures of intervenolin, CJ-13,136 and CJ-13,217.
Effect of intervenolin on coculture of tumor cells and stromal cells
To determine the growth of tumor cells selectively when cocultured with stromal cells, we stably transfected GFP gene into tumor cells used in this study. As shown in Figure 5, intervenolin inhibited the growth of human gastric cancer MKN-74 cells cultured alone with an IC50 at 3.0 μg ml−1, but it inhibited the growth of MKN-74 cells cocultured with Hs738 human gastric stromal cells more strongly with an IC50 at 0.17 μg ml−1. Intervenolin also inhibited the growth of human gastric cancer MKN-7 cells and human colorectal cancer HCT-15 cells cocultured with Hs738 cells and CCD-18Co human colorectal stromal cells, respectively, at lower concentrations than those of cancer cells cultured alone. While the IC50 for the coculture of MKN-7 and HCT-15 cells were 0.13 μg ml−1 and 0.21 μg ml−1, respectively, the IC50 for the monoculture of MKN-7 and HCT-15 cells were 1.6 μg ml−1 and 1.6 μg ml−1, respectively (Figure 5). On the contrary, intervenolin did not show such selective growth inhibitory activity against human lung cancer NCI-H460 cells. It inhibited the growth of NCI-H460 cells cocultured with NHLF human lung stromal cells to the same extent as the cancer cells cultured alone with IC50 at about 2 μg ml−1. Intervenolin inhibited the growth of various stromal cells with IC50 at around 2–5 μg ml−1, which were almost the same as IC50 for monocultures of cancer cells.

Effect of intervenolin on coculture of tumor cells and stromal cells. The growth of tumor cells cocultured with stromal cells (•) or that of tumor cells alone (○) in the presence of the indicated concentrations of intervenolin was determined measuring GFP fluorescence intensity. MKN-74 and MKN-7 human gastric cancer cell lines were cocultured with Hs738 gastric stromal cells. HCT-15 human colorectal cancer cells and NCI-H460 human lung cancer cells were cocultured with CCD-18Co colorectal stromal cells and NHLF lung stromal cells, respectively. The growth of stromal cells cultured with the indicated concentrations of intervenolin for 3 days was determined by MTT. Values are means of duplicate determinations. Each s.e. is less than 10%.
CJ-13,136 and CJ-13,217 also showed selective growth inhibitory activity against the growth of MKN-74 cells cocultured with Hs738 cells compared with the monoculture (Figure 6). The IC50 for coculture and monocultures were 0.01 and 0.25 μg ml−1 (CJ-13,136) and 0.006 and ∼0.03 μg ml−1 (CJ-13,217), respectively, and they were very low compared with that of intervenolin. However, their acute toxicities against mice were very strong. Maximum tolerated dose was estimated for CJ-13,136 at 2.5 mg kg−1 by i.v. injection and for CJ-13,217 at 3 mg kg−1 by i.p. injection. On the contrary, maximum tolerated dose for intervenolin was estimated more than at 50 mg kg−1 by i.v. injection, because no fatality was observed up to 50 mg kg−1.

Effects of CJ-13,136 and CJ-13,217 on coculture of MKN-74 cells and Hs738 cells. The growth of MKN-74 cells cocultured with Hs738 cells (•) or that of MKN-74 cells alone (○) in the presence of the indicated concentrations of test compounds was determined measuring GFP fluorescence intensity. Values are means of duplicate determinations. Each s.e. is less than 10%.
Antitumor effect of intervenolin in vivo
Because the acute toxicity of intervenolin was very low, we examined the antitumor effect against a mice xenograft model of human colorectal cancer. While HCT-15 colorectal cancer cells made tumors when inoculated s.c. into nude mice, the growth was apparently increased by the coinoculation of CCD-18Co stromal cells (608.3±128.0 mg versus 715.1±68.0 mg with CCD-18Co, P<0.05) (Figure 7). Intervenolin at 25 mg kg−1 was found to significantly inhibit the tumor growth of HCT-15 cells in vivo to the same extent as that with coinoculation of CCD-18Co stromal cells (Figure 7).

Effect of intervenolin on tumor growth of HCT-15 cells in vivo. HCT-15 human colorectal cancer cells were inoculated s.c. with (below) or without (above) CCD-18Co colorectal stromal cells in female nude mice. Intervenolin was administered i.v. at 12.5 and 25 mg kg−1 on days indicated by arrows. The mice were killed on day 21 after the tumor inoculation, and the tumors were excised. The values are means±s.d. of five mice. *P<0.05 and **P<0.01 versus the values with vehicle.
Anti-Helicobacter pylori effect
CJ-13,136 and CJ-13,217 were reported to show anti-Helicobacter pylori effect.17 We next examined whether intervenolin exerts anti-H. pylori effect. As shown in Table 3, intervenolin showed potent anti-H. pylori activity against H. pylori JCM12093 and H. pylori JCM12095 strains with MICs at 0.016 and 0.008 μg ml−1, respectively. The MICs of intervenolin against H. pylori were almost the same as clarithromycin used as a positive control, but intervenolin failed to show antimicrobial activity against other bacteria such as Staplylococcus aureus, Enterococcus faecalis and Haemophilus influenzae, unlike clarithromycin and ampicillin.
Discussion
Growing evidence strengthens the important role of stromal cells in tumor progression.1, 2, 3, 4, 5, 6, 7 We have been focusing on the stromal cells and finding small molecules that modulate the tumor–stromal cell interactions.10, 11, 12, 13 Because some of the compounds successfully exert antitumor effects in vivo,12, 13 we are confident that targeting the stromal cells can be a new antitumor strategy. We have done initiative studies using prostate cancer cells and now we are applying the same strategy to other cancer. Developing the screening systems for cocultures of lung, gastric and colorectal cancer with the respective organ-derived stromal cells, we have been searching a modulator of the tumor–stromal cell interactions. As a result, we have discovered a novel compound, intervenolin, from natural resources.
Intervenolin showed stronger growth inhibitory activity against some cancer cell lines when cocultured with stromal cells compared with the cancer cells cultured alone. However, its effect is considered to be selective, because intervenolin did not show such effect against lung cancer cells suggesting the involvement of some specific interactions between tumor cells and stromal cells. As the IC50 of intervenolin for monocultures of cancer cells are almost the same as the IC50 for stromal cells, these effects on monocultures are considered to be non-specific cytotoxicity. Although the structure of intervenolin is similar to that of CJ-13,136 and CJ-13,217, the side chain (-C-N=C-(SCH3)2) is very unique to intervenolin. CJ compounds showed somewhat same effect as intervenolin on the coculture of gastric cancer cells, but their acute toxicities were very strong. In contrast, acute toxicity of intervenolin was very weak, so we could assess the antitumor effect in vivo. As a result, we have found that intervenolin significantly inhibits the tumor growth of colorectal cancer cells in vivo without any severe adverse effects. There is a possibility that the side chain of intervenolin, (-C-N=C-(SCH3)2), potentiates its bioavailability.
Unexpectedly, intervenolin inhibited the tumor growth of colorectal cancer cells regardless of the coinoculation of CCD-18Co human colorectal stromal cells. However, our preliminary experiments have shown that stromal cells derived from a host mouse apparently get in the xenograft tumor tissues. Thus, it is considered that intervenolin could act on the interactions between the tumor cells and the mouse stromal cells. To confirm the interactions in vitro, we are trying to isolate the mouse stromal cells from the xenograft tumor tissues. We are also now studying the mechanism of action of intervenolin, but our preliminary results suggest that intervenolin would affect secreted factors from the stromal cells.
Because CJ compounds are reported to show anti-H. pylori activity, we examined the antimicrobial activity of intervenolin. As a result, intervenolin expectedly showed potent selective anti-H. pylori activity without effect on any other bacteria. Although whether the mechanism of action of intervenolin against H. pylori correlates with antitumor effect is still unknown, intervenolin is a very unique compound possessing both potential activities.
The strain Nocardia sp. ML96-86F2 is found to also produce CJ-13,136 and CJ-13,217 at a trace amount, but the strain is not identical to the CJ compounds-producing Pseudonocardia sp. CL38498.17 However, the productivity of intervenolin is low and not satisfactory for further animal experiments. To obtain sufficient amount of intervenolin for our further experiments and also for the creation of more potent derivatives, we are now trying to prepare the compounds by chemical synthesis.
Methods
Microorganism
The strain ML96-86F2 was isolated from a soil sample collected in Tokyo, Japan. The isolate was maintained in glucose-asparagine agar consisting of 1% glucose, 0.05% L-asparagine, 0.05% K2HPO4 and 2% agar (pH 7.0), and in yeast-starch agar consisting of 0.2% yeast extract, 1% soluble starch and 2% agar (pH 7.0). ML96-86F2 was deposited in the culture collection of NITE Biological Resource Center (Chiba, Japan) as NITE P-1464.
Taxonomic studies
Cultural characteristics were determined by growth of ISP medium No. 2-7.14 The morphological properties were observed by a electron microscopy (Hitachi S-500; Hitachi, Tokyo, Japan) scanning a strain grown on sucrose-nitrate agar consisting of 3% sucrose, 0.2% NaNO3, 0.1% K2HPO4, 0.05% KCl, 0.05% MgSO4·7 H2O, 0.001% FeSO4·7 H2O and 2% agar (pH 7.2), at 30 °C for 14 days. For chemotaxonomic studies, freeze-dried cells were obtained from cultures grown in yeast extract-glucose medium (YD-medium consisting of 1% yeast extract and 1% glucose, pH 7.0) on a rotary shaker at 28 °C for 7 days. Analyses of the isomers of diaminopimelic acid in whole-cell hydrolysate were done by the method of Stanek and Roberts.15 Extraction and analysis of mycolic acids were assessed by the method of Minnikin et al.16 Genomic DNA was purified using Genomic DNA Extraction Kit Mini (RBC Bioscience, Taipei, Taiwan). The 16 S rDNAs were amplified by PCR with a PCR thermal cycler (iCycler, Bio-Rad Laboratories, CA, USA) using universal primers: 9 F (5′-GAGTTTGATCCTGGCTCAG-3′) and 1541 R (5′-AAGGAGGTGATCCAGCC-3′).
Fermentation
The strain Nocardia sp. ML96-86F2 grown on an agar slant was inoculated into 100 ml of a seed medium containing 2% galactose, 2% dextrin, 1% Bacto Soytone (BD Biosciences, Tokyo, Japan), 0.5% corn steep liquor (Kogo Starch, Chiba, Japan), 0.2% (NH4)2SO4 and 0.2% CaCO3 (adjusted at pH7.2 before sterilization) and cultured at 30 °C for 8 days on a rotary shaker at 220 r.p.m. Two milliliters of the seed culture was inoculated into a 500-ml flask containing 100 ml of a culture medium consisting of 1% glucose, 2% corn starch (Nihon Shokuhin Kako, Tokyo, Japan), 0.5% wheat germ (Nisshin Seifun, Tokyo, Japan), 0.5% NZ Amine type A (Wako Pure Chemical Industries, Osaka, Japan), 0.5% Bacto yeast extract (BD Biosciences), 0.4% CaCO3 and 0.0001% CoCl2 (adjusted at pH 6.7 before sterilization) as described17 and cultured at 25 °C for 11 days on a rotary shaker at 220 r.p.m.
Analytical measurement
UV spectrum was recorded on a Hitachi U-2910 spectrometer. 1H- and 13C-NMR spectra were measured on a JEOL JNM A400 spectrometer (JEOL, Tokyo, Japan) using tetramethylsilane as an internal standard. 1H and 15N NMR spectra were measured on a JEOL JNM ECA600 spectrometer (JEOL) using tetramethylsilane as an internal standard. HRESI-MS spectrum was measured with a Thermo Fisher Scientific LTQ Orbitrap XL mass spectrometer (Thermo Fisher Scientific, Waltham, MA, USA). IR spectrum was measured with a Horiba FT-210 Fourier transform infrared spectrometer (Horiba, Kyoto, Japan).
Cells and reagents
Human gastric cancer MKN-7 and MKN-74 cell lines were obtained from RIKEN cell bank (Tsukuba, Japan), human colorectal cancer HCT-15 cells and human lung cancer NCI-H460 cells were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). All cancer cell lines were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM) (Nissui, Tokyo, Japan) supplemented with 10% fetal bovine serum (FBS; Sigma, Tokyo, Japan), 100 units per ml penicillin G (Life Technologies, Carlsbad, CA, USA) and 100 μg ml−1 streptomycin (Life Technologies) at 37 °C with 5% CO2. Hs738 human gastric stromal cells (CRL-7869) and CCD-18Co human colorectal fibroblasts (CRL-1459) were obtained from ATCC. NHLF human lung stromal cells were obtained from BioWhittaker (Walkersville, MD, USA). All stromal cells were maintained in DMEM supplemented with 10% FBS, 100 units per ml penicillin G, 100 μg ml−1 streptomycin, ITH (5 μg ml−1 insulin, 5 μg ml−1 transferrin and 1.4 μM hydrocortisone) and 5 ng ml−1 basic FGF (Pepro Tech, Rocky Hill, NJ, USA) at 37 °C with 5% CO2 as described.8 CJ-13,136 and CJ-13,21717 were purified in our laboratory.
GFP transfection
Cells (2 × 105) were cultured in six-well plates in 10% FBS-DMEM for 2 days. The medium was replaced by 800 μl of OPTI-MEM (Life Technologies), and then 200 μl of OPTI-MEM containing 1 μg of pEGFP-C1 vector (BD Biosciences), 4 μl of lipofectamine reagent (Life Technologies) and 6 μl of PLUS Reagent (Life Technologies) was added to the cells. After 3 h incubation, 1 ml of 20% FBS-OPTI-MEM was added to each well and the cells were cultured for 24 h. Stably transfected cells were selected with 400 μg ml−1 of G418 (Promega, Tokyo, Japan).
Cell growth and coculture experiment
Cells were inoculated into a 96-well plate at 5 × 103 cells per well in 0.1 ml of DMEM supplemented with 1% dialyzed FBS (D-FBS) and ITH. After culture for the indicated days, the cell growth was determined using 3-(4,5-dimethyllythiazol-2-yl)-2,5- diphenyltetrazolium bromide (MTT; Sigma) as described18 or measuring GFP fluorescence intensity (excitation at 485 nm and emission at 538 nm) by the lysis of cells in 10 mM Tris-HCl (pH 7.4), 150 mM NaCl, 0.9 mM CaCl2 and 1% Triton X-100. For coculture experiment, stromal cells were first inoculated in 96-well plates at 5 × 103 cells per well in 0.1 ml of DMEM supplemented with 1% D-FBS and ITH. Test samples were added into the well and the stromal cells were cultured for 2 days. Then, 10 μl of cancer cell suspension (5 × 103) in serum-free DMEM were inoculated onto a monolayer of the stromal cells and the cells were further cultured for 3 days. For monoculture of cancer cells, only assay medium with test samples was first incubated for 2 days, and then cancer cells were inoculated as described above and further cultured for 3 days. The growth of the cancer cells was determined measuring GFP fluorescence intensity.
Acute toxicity in vivo
Female ICR mice, 4-week old, were purchased from Charles River Breeding Laboratories (Yokohama, Japan) and maintained in a specific pathogen-free barrier facility according to our institutional guidelines. Samples were injected i.v. or i.p. into mice. During 2-week observation, half of a dose that caused death or severe toxic effect is designated as a maximum tolerated dose in this study.
Antitumor effect in vivo
Female BALB/c nude mice, 5 weeks old, were purchased from Charles River Breeding Laboratories (Yokohama, Japan) and maintained in a specific pathogen-free barrier facility according to our institutional guidelines. HCT-15 cells (8 × 106) were trypsinized and resuspended with or without CCD-18Co (8 × 106) in 0.3 ml of 10% FBS-DMEM and then combined with 0.5 ml of growth factor-reduced Matrigel (BD Biosciences). One hundred microliters of the cell suspension (1 × 106 cells) were injected s.c. in the left lateral flank of mice. Five mice were used for each experimental set. Test samples were administered as indicated. Tumor volume was estimated using the following formula: tumor volume (mm3)=(length × width2)/2. After the indicated times, tumors were surgically dissected.
Antimicrobial effect
Helicobacter pylori JCM12093, H. pylori JCM12095 and Enterococcus faecalis JCM5803 were purchased from Japan Collection of Microorganisms (Tsukuba, Japan). Haemophilus influenzae T-196 and H. influenzae ARD476 were generously provided by Prof. Kimiko Ubukata (Kitasato University). MICs were determined by using microdilution broth method in 96-well plates. Twofold serial dilutions of the test samples were prepared with dimethyl sulfoxide. The dilution series were initially prepared at 200-fold final concentrations from this stock to provide a final test range of 128–0.001 μg ml−1. Fifty microliters of each dilution and 50 μl of test organisms (2–9 × 104 cfu per ml) were then dispensed into each well. The MIC was defined as the lowest concentration of compound at which no visible growth could be seen. The test organisms, medium and culture conditions were following: Helicobacter pylori JCM12093 and H. pylori JCM12095 were cultured in a Brain Heart Infusion Broth medium (BD Biosciences) with 10% FBS at 37 °C for 144 h under microaerobic conditions of 85% N2, 5% O2 and 10% CO2; Staphylococcus aureus FDA209P and Escherichia coli K-12 were cultured in a nutrient broth medium consisting of 1% polypentone (Nihon Phamaceutical, Tokyo, Japan), 1% fish extract (Kyokuto, Tokyo, Japan) and 0.2% NaCl 18 h at 37 °C; Enterococcus faecalis JCM5803 was cultured in a Heart Infusion Broth medium (BD Biosciences) at 37 °C for 18 h; H. influenzae T-196 and H. influenzae ARD476 were cultured in a Muller Hinton medium (BD Biosciences) with 5% Fildes Enrichment (BD Biosciences) at 37 °C for 18 h.
References
Wernert, N. The multiple roles of tumour stroma. Virchows Arch. 430, 433–443 (1997).
Tuxhorn, J. A. et al. Reactive stroma in human prostate cancer: induction of myofibroblast phenotype and extracellular matrix remodeling. Clin. Cancer Res. 8, 2912–2923 (2002).
Wiseman, B. S. & Werb, Z. Stromal effects on mammary gland development and breast cancer. Science 296, 1046–1049 (2002).
Hwang, R. F. et al. Cancer-associated stromal fibroblasts promote pancreatic tumor progression. Cancer Res. 68, 918–926 (2008).
Picard, O., Rolland, Y. & Poupon, M. F. Fibroblast-dependent tumorigenicity of cells in nude mice: implication for implantation of metastases. Cancer Res. 46, 3290–3294 (1986).
Camps, J. L. et al. Fibroblast-mediated acceleration of human epithelial tumor growth in vivo. Proc. Natl Acad. Sci. USA 87, 75–79 (1990).
Micke, P. & Ostman, A. Tumour-stroma interaction: cancer-associated fibroblasts as novel targets in anti-cancer therapy? Lung Cancer 45 (Suppl 2), S163–S175 (2004).
Kawada, M. et al. A microplate assay for selective measurement of growth of epithelial tumor cells in direct coculture with stromal cells. Anticancer Res. 24, 1561–1568 (2004).
Kawada, M., Inoue, H., Masuda, T. & Ikeda, D. Insulin-like growth factor I secreted from prostate stromal cells mediates tumor-stromal cell interactions of prostate cancer. Cancer Res. 66, 4419–4425 (2006).
Kawada, M., Inoue, H., Usami, I. & Ikeda, D. Phthoxazolin A inhibits prostate cancer growth by modulating tumor-stromal cell interactions. Cancer Sci. 100, 150–157 (2009).
Kawada, M., Momose, I., Someno, T., Tsujiuchi, G. & Ikeda, D. New atpenins, NBRI23477 A and B, inhibit the growth of human prostate cancer cells. J. Antibiot. 62, 243–246 (2009).
Kawada, M. et al. NBRI16716A, a new antitumor compound against human prostate cancer cells, produced by Perisporiopsis melioloides Mer-f16716. J. Antibiot. 63, 319–323 (2010).
Kawada, M. et al. Leucinostatin A inhibits prostate cancer growth through reduction of insulin-like growth factor-I expression in prostate stromal cells. Int. J. Cancer 126, 810–818 (2010).
Shirling, E. B. & Gottlieb, D. Methods for characterization of Streptomyces species. Int. J. Syst. Bacteriol. 16, 313–340 (1966).
Stanek, J. L. & Roberts, G. D. Simplified approach to identification of aerobic actinomycetes by thin-layer chromatography. Appl. Microbiol. 28, 226–231 (1974).
Minnikin, D. E., Hutchinson, I. G., Caldicott, A. B. & Goodfellow, M. Thin-layer chromatography of methanolysates of mycolic acid-containing bacteria. J. Chromatogr. 188, 221–233 (1980).
Dekker, K. A. et al. New quinolone compounds from Pseudonocardia sp. with selective and potent anti-Helicobacter pylori activity: taxonomy of producing strain, fermentation, isolation, structural elucidation and biological activities. J. Antibiot. 51, 145–152 (1998).
Fukazawa, H., Mizuno, S. & Uehara, Y. A microplate assay for quantitation of anchorage-independent growth of transformed cells. Anal. Biochem. 228, 83–90 (1995).
Acknowledgements
We thank Dr R Sawa and Ms Y Kubota for analysis of HRESI-MS and NMR measurements, and M Umekita, K Adachi, E Sato and K Hitosugi for their technical assistance. We thank Professor K Ubukata (Kitasato University) for providing us H. influenzae strains. We also thank Dr I Momose, Dr T Someno and Professor M Shibasaki for their valuable discussions. This paper is dedicated to the memory of Naoko Kinoshita, who suddenly passed away in 2012. This study was supported by MEXT KAKENHI Grant Number 23501321.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kawada, M., Inoue, H., Ohba, Si. et al. Intervenolin, a new antitumor compound with anti-Helicobacter pylori activity, from Nocardia sp. ML96-86F2. J Antibiot 66, 543–548 (2013). https://doi.org/10.1038/ja.2013.42
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ja.2013.42
Keywords
This article is cited by
-
The therapeutic potential of mitochondrial toxins
The Journal of Antibiotics (2021)
-
Bioactive molecules fromNocardia: diversity, bioactivities and biosynthesis
Journal of Industrial Microbiology and Biotechnology (2019)
-
Biological activity of intervenolin analogs with a phenyl substituent
The Journal of Antibiotics (2018)
-
Novel approaches for identification of anti-tumor drugs and new bioactive compounds
The Journal of Antibiotics (2018)
-
Small molecules modulating tumor–stromal cell interactions: new candidates for anti-tumor drugs
The Journal of Antibiotics (2016)
