
Abstract
Antibiotics are low-molecular-mass products of secondary metabolism, nonessential for the growth of producing organisms, but very important for human health. They have unusual structures and are most often formed during the late growth phase of the producing microorganisms. Their production arises from intracellular intermediates, which are condensed into more complex structures through defined biochemical pathways. Their synthesis can be influenced by manipulating the type and concentration of nutrients formulating the culture media. Among them, the effect of the carbon source has been the subject of continuous studies for both industry and research groups. Glucose and other carbohydrates have been reported to interfere with antibiotic synthesis and this effect depends on the rapid utilization of the preferred carbon source. Different mechanisms have been described in bacteria and fungi to explain the negative effects of carbon catabolites on antibiotic production. They show important differences depending on the microbe being considered. Their understanding and manipulation have been useful for both perfecting fermentation conditions to produce anti-infectives and for strain improvement. To improve the production of antibiotics, carbon source repression can be decreased or abolished by mutations resulting in antimetabolite resistance. Enzymes reported as regulated by the carbon source have been used as targets for strain improvement. During the last few years, important advances have been reported elucidating the essential aspects of carbon source regulation on antibiotic production at biochemical and molecular levels. The aim of this review is to describe these advances, giving special emphasis to those reported for the genus Streptomyces.
Similar content being viewed by others

Introduction
Antibiotics are low-molecular-mass (<1500 kDa) products of secondary metabolism, usually produced during the late growth phase (idiophase) of a relatively small group of microorganisms. Antibiotics are not essential for the growth of producing cultures but serve diverse survival functions in nature.1 In addition, antibiotics are very important for the health, nutrition and economics of our society.2
Owing to the use of antibiotics and other secondary metabolites, the average life expectancy in the United States increased from 47 years in 1900–1974 (males) to 80 years (females) in the year 2000.3 Probably, the most important use of secondary metabolites has been as anti-infective drugs. In the year 2000, anti-infective secondary metabolites marketed 55 billion dollars,4 but in the year 2007, the market for antibiotics increased to 66 billion dollars.5
If modern medicine is to continue in its present form, novel families of antibiotics must enter the marketplace at regular intervals. Recently, new antibiotics have been introduced to the anti-infectious disease market, for example, augmentin, clarithromycin and ceftriaxone. However, within the next 10 years, aggressive screening programs for the selection of novel natural and modified chemical compounds will be necessary to produce novel antibiotics against resistant bacteria.
In addition to the screening programs for new compounds with antibacterial activity, during the last few decades, the pharmaceutical industry has extended these programs to other disease areas6, 7, 8 such as anticancer drugs, namely bleomycin, dactinomycin, doxorubicin and staurosporin;9 antimycotic agents, namely anidulafungin;10 and antihelmintic compounds, namely ivermectin, milbemycin and spinosyns.11 In addition, the National Cancer Institute has invested considerable effort in evaluating the inhibitory activity of pepstatin on HIV-1 (human immunodeficiency virus-1). Pepstatin A is a small pentapeptide produced by several Streptomyces species with a unique hydroxyamino acid (statine) that sterically blocks the active site of HIV-1 protease.12, 13
The outstanding role of microorganisms in the production of antibiotics is notorious. At present, with <1% of the microbial world having been explored, the advances in techniques for microbial cultivation and extraction of nucleic acids from soil and marine habitats are allowing access to a vast untapped reservoir of genetic and metabolic diversity.14
The main biosynthetic pathways involved in antibiotic production are those forming peptides, polyketides, isoprenes, oligosaccharides, aromatic compounds and β-lactam rings. Knowledge of these pathways varies from those cases in which the amino-acid sequences of the enzymes and nucleotide sequences of the genes are known (for example, for penicillins and cephalosporins), to those in which even the enzymatic steps are still unknown.15
Antibiotics are formed by enzymatic pathways that occur through individual proteins, free or complexed, or through parts of large multifunctional polypeptides carrying out a multitude of enzymatic steps, for example, polyketide synthases and peptide synthetases.16 The genes encoding these enzymes are usually chromosomal. For example, the 30 biosynthetic str/sts and blu genes of streptomycin biosynthesis (and of related antibiotics such as 5′-hydroxystreptomycin and bluensomycin) that were cloned from Streptomyces griseus and Streptomyces glaucescens were all found in a chromosomal region of 30–40 kb.17 Only a few biosynthetic genes have been shown to be plasmid borne, for example, those encoding methylenomicin A synthesis in Streptomyces coelicolor and antibiotic AS-48 in Enterococcus faecalis S-48.18 Whether chromosomal or plasmid borne, antibiotic formation genes are usually clustered, especially in prokaryotes, but not necessarily as single operons. Antibiotic clusters in actinomycetes have different sizes. For instance, although actinorhodin and other simple aromatic polyketides are encoded in chromosomal regions of 20 kb or less, regions of 90–100 kb are required to encode for rapamycin and rifamycin and other complex polyketides. In the latter case, 50–80 kb of DNA are required for the modular poliketyde synthetases (PKSs) themselves.19 Clusters of fungal biosynthetic genes have also been found for biosynthetic processes of sterigmatocystin formation by Aspergillus nidulans.20
Antibiotic formation usually occurs during the late growth phase of the producing microorganism. The temporal nature of their formation is certainly genetic, but expression can be influenced greatly by environmental manipulations. Therefore, synthesis of antibiotics is often brought on by exhaustion of a nutrient, addition of an inducer and/or by a decrease in growth rate.21 These events generate signals that cause a cascade of regulatory events resulting in chemical (secondary metabolism) and morphological differentiation (morphogenesis) of the microbial antibiotic producers. The signal is often a low-molecular-weight butyrolactone inducer that acts by binding to and inactivating a regulatory protein (repressor protein/receptor protein) that normally prevents secondary metabolism and morphogenesis during rapid growth and nutrient sufficiency.22 Formation of antibiotics is also regulated by nutrients (such as nitrogen, phosphorous and carbon source), metals, growth rate, feedback control and enzyme inactivation.23 Among these nutrients, the effect of carbon source on antibiotic production has been the subject of continuous study for both industry and research groups, not only from fermentation but also from biochemical and molecular biological stand points. In this work, we review the regulation of antibiotic biosynthesis by carbon source, and the mechanisms involved, giving special emphasis to antibiotics produced by the genus Streptomyces. We describe how some proteins and protein complex systems are crucial for carbon source effect on antibiotic biosynthesis. These proteins and systems can be grouped as follows: (1) the phosphoenolpyruvate: phosphotransferase system (PTS), one of the most important mechanisms utilized for both Gram-negative and Gram-positive bacteria with low guanine and cytosine (GC) content; (2) CarI (for N-3-(oxohexanoyl)-L-homoserine lactone (OHHL) synthesis), a key factor used for Erwinia; (3) Bld and Reg1, transcriptional regulators in Streptomyces; (4) other Bld proteins (such as BldA, BldG and BldH) for Streptomyces; (5) the Glk enzyme for the genus Streptomyces; (6) the SCO2127 protein, which propels production of metabolic signals responsible for carbon catabolite repression (CCR) in Streptomyces; (7) 3,5-cyclic adenosine monophosphate (cAMP) and its binding protein (cAMP receptor protein (CRP)), very important for Gram-negative bacteria and for the synthesis of some antibiotics in Streptomyces; (8) the CreA zinc-finger repressor in fungi; and (9) the catabolite control protein A (CcpA) binding to cre elements in Bacillus. In addition, direct carbon source inhibition of key enzymes involved in secondary metabolic pathways will also be documented.
Regulation by carbon source
In bacteria and other microorganisms, glucose, usually an excellent carbon source for growth, interferes with the formation of many antibiotics ‘too much of a good thing can be bad.’24 In media containing mixtures of rapidly and slowly used carbon sources, the former is used first to produce cells but little or no antibiotics are synthesized. After the rapidly assimilated compound is depleted, the ‘second-best’ carbon source is used for idiolite formation.
Regulation by the carbon source depends on the rapid utilization of the preferred carbon source. One distinctive characteristic of secondary metabolism is its association with low growth levels;24 thus, low concentrations of the repressing carbon source in the culture medium yield low growth rates and eliminate the interference of antibiotic biosynthesis.
Different mechanisms have been described in bacteria to explain the negative carbon catabolite effects on secondary metabolite production. These mechanisms show important differences depending on the type of bacteria being considered. For a detailed review about these mechanisms, refer to the studies by Deutscher25 and Görke and Stülke.26 A few characteristics of these mechanisms will be mentioned in this review. However, major emphasis will be given to the advances in the mechanism of carbon catabolite repression (mechanisms that inhibit synthesis at the level of DNA transcription) in the genus Streptomyces.
Carbon source regulation of antibiotic production in Gram-negative bacteria
Gram-negative bacteria produce a particular type of secondary metabolites. In some cases, their production is sensitive to carbon source repression. One example is carbapenem biosynthesis in Erwinia carotovora.27 In this microorganism, glycerol suppresses (a mechanism that arrests metabolites formation and may be the consequence of repression or inhibition) carbapenem biosynthesis by repressing transcription of the carI gene responsible for the synthesis of OHHL, a quorum-sensing molecule that induces antibiotic formation. Limitation of OHHL affects the expression of carbapenem biosynthetic genes (carA-H) by preventing formation of the complex between CarR (DNA-binding transcriptional activator of carA-H) and OHHL. The formation of this complex (CarR–OHHL) is a necessary step for transcriptional activation of the carbapenem biosynthetic genes in E. carotovora.28, 29 On the other hand, glycerol does not affect transcription of either carR or hor (transcriptional activator of antibiotic and pigment production) genes.27 Carbapenem-like antibiotics, are also produced by other Gram-negative bacteria such as Erwinia herbicola, Serratia sp. strain American Type Culture Collection 39006 and Photorhabdus luminescens strain TT01.30 Interestingly, in P. luminescens, the carR gene is not present, suggesting a different mechanism for the regulation of antibiotic formation. In addition, in contrast to E. carotovora, P. luminescens carbapenem-like synthesis is not induced by the quorum-sensing molecule N-acyl-homoserine lactone.30
An additional example of carbon regulation has been reported in the ethanol-utilizing strain, Serratia marcesens. In this microorganism (strain S389), Cang et al.31 observed that a low glucose concentration (27.7 mM) added to the growth medium, almost completely repressed formation of the red tripirrole antibiotic, prodigiosin. Other carbon sources such as galactose, fructose and sucrose also repressed antibiotic production. As mentioned above, Serratia also produces the antibiotic carbapenem. It is known that biosynthesis of both compounds (that is, carbapenem and prodigiosin) are under quorum-sensing control.32, 33 These secondary metabolites are controlled by the smaR quorum-sensing locus, which represses both compounds when levels of N-acyl-homoserine lactone are low. Quorum sensing controls the production of prodigiosin, at least in part, by modulating transcription of three genes encoding regulatory proteins. Surprisingly, one of these proteins is a novel putative adenylate cyclase,34 which likely causes a decrease in cellular cAMP concentration affecting pigment production.
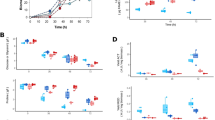
Among the mechanisms of CCR in Gram-negative bacteria, the phosphoenolpyruvate: PTS can be mentioned as one of the most important. PTS utilizes a protein phosphoryl transfer chain to transport and phosphorylate its sugar substrates. In Escherichia coli, PTS consists of four high-energy phosphoprotein intermediates and five protein domains. One of these proteins, EIIAglc is phosphorylated by a phosphoprotein (HPr) on His90. Subsequently, EIIAglc∼P transfers its phosphate from His75 to a high-affinity homodimeric protein EIIB/C found in the cell membrane. The IIC domain crosses the membrane eight times harboring the sugar-binding site. The hydrophilic domain IIB transfers the phosphate group from EIIAglc∼P to glucose, to form glucose 6-phosphate (Figure 1).25

Model for carbon catabolite repression (CCR) mechanism in Escherichia coli. In white background is shown a cell under repressing conditions and in light yellow under non-repressing conditions. The phosphotransferase system (PTS) comprising Enzyme I, HPr and IIAGlc phosphorylating enzymes, is the most important CCR mechanism in E. coli. In the presence of glucose, PTS internalizes and phosphorylates the sugar, rendering dephosphorylated IIAGlc, which in turn exerts inducer exclusion of other non- or less-repressing carbon sources. On the contrary, when glucose is absent, IIAGlc remains phosphorylated and activates the adenylate cyclase (AC). AC synthesizes cAMP, which binds to the cAMP receptor protein (CRP), activating the transcription of CCR-sensitive genes.
In addition to transferring the phosphate group to EIIB/C, EIIAglc∼P activates adenylate cyclase. The activated form of this enzyme synthesizes cAMP, necessary for the synthesis of inducible enzymes and its intracellular levels mediate CCR. To activate transcription, cAMP binds to the DNA promoter region through the CRP.
In the presence of glucose, sugar is transported into the cell and concomitantly phosphorylated. This event causes dephosphorylation of EIIAglc∼P, mediates inducer exclusion and deactivates adenylate cyclase. Inactivation of adenylate cyclase causes a reduction in cytoplasmic cAMP concentration and promotes dissociation of the cAMP–CRP complex from the DNA and deactivation of transcriptional initiation. The gene for EIIAglc is known as crr, because mutants of E. coli lacking this gene are resistant to CCR.25
While studying N-acyl amino-acid synthase clones derived from environmentally obtained DNA, Clardy and Brady35 discovered an N-acyl amino-acid synthase (NasP) containing the cAMP-binding domain. Studies on the regulation of the antibiotic N-acylphenylalanine showed that antibiotic biosynthesis is cAMP dependent. In addition, using affinity chromatography, these authors confirmed a direct interaction between NasP and cAMP.
Pseudomonas species produce several types of antifungal compounds, of which the most studied are pyoluteorin, pyrrolnitrin, phenazines and 2, 4-diacetylphloroglucinol (Phl). In addition to its antifungal activity, Phl has shown an interesting bactericidal activity against methicillin-resistant Staphylococcus aureus.36
Phenazines are heterocyclic compounds the effects of which are attributed to their redox activity. For example, the 2-hydroxy-phenazine-1 carboxylic acid produced by Pseudomonas aureofaciens is believed to destroy competing fungi through production of reactive oxygen species.37 The expression of phenazine biosynthetic genes is regulated by multiple mechanisms, which are strongly influenced by environmental conditions. One of the primary factors governing phenazine production is population density, and in Pseudomonas aeruginosa, this dependency is affected by at least three quorum-sensing systems.38
In several species of Pseudomonas, phenazine production is affected by the carbon source. For example, in Pseudomonas chloraphis, the greatest production of phenazine 1-carboximide was obtained using L-pyroglutamic acid and glucose. However, fructose, sucrose and ribose had negative effects on phenazine 1-carboximide production.38 Similar results were observed in a mutant of Pseudomonas sp. M18G, in which the greatest production of phenazine 1-carboxylic acid was achieved with glucose and ethanol as carbon sources.39
In fluorescent Pseudomonas, Kreb cycle intermediates, such as succinate, induce catabolite repression for the utilization of nonpreferred carbon sources, such as glucose. cAMP and its respective cAMP-binding protein do not seem to be responsible for catabolite repression mechanisms in this genus.40
A master regulator of carbon metabolism in Pseudomonas is an RNA-binding protein named Crc, which, under repressive conditions, inhibits the translation of mRNAs by binding to a sequence just downstream of the AUG start codon (Figure 2).41 Conversely, under nonrepressive conditions, a small RNA named CrcZ binds to the Crc protein to relieve mRNAs from catabolite repression. A two-component system CbrA/CbrB, which is sensitive to the carbon source, controls the expression of CrcZ.40

Schema of catabolite repression exerted by Crc/CrcZ. When a preferred carbon source such as succinate is present in the culture medium, there is a low level of CrcZ sRNA; thus Crc attaches to catabolite repression-sensitive mRNAs, inhibiting translation. Conversely, in the presence of a non-preferred carbon source such as mannitol, the expression of CrcZ sRNA is increased due to the induction of the CbrA/CbrB two-component system. The larger amount of CrcZ is then capable of binding to Crc protein, releasing mRNAs and allowing translation. Modified from Sonnleiter et al.40
In the same manner, other RNA-binding proteins such as RsmA (repressor of secondary metabolism) from Pseudomonas fluorescens avoid translation by attachment to mRNAs related to the synthesis of extracellular antifungal secondary metabolites. Translation of mRNAs can continue when the cell expresses high quantities of small RNAs that compete for binding to RsmA, which is similar to the carbon storage regulator, CsrA from E. coli and the regulation of sRNA expression is promoted by a two-component system named GacS/GacA.42
Carbon source regulation of antibiotic production in Gram-positive bacteria
Gram-positive bacteria are characterized by having multiple layers of peptidoglycans as well as proteins and/or teichoic acids as part of their cell-wall structure. In addition, the GC content of the DNA significantly varies among these bacteria. Thus, the phylum Firmicutes corresponds to Gram-positive bacteria with low GC content, whereas the phylum Actinobacteria contains high GC content in their DNA structure. Firmicutes include the classes Bacilli, Chlostridia and Erysipelotrichi.43 The phylum Actinobacteria is constituted by the class Actinobacteria and includes the order Actinomycetales.44 The phylum Actinobacteria represents one of the largest and most diverse groups of bacteria, encompassing many important and well-characterized organisms, including Streptomyces, Bifidobacterium, Corynebacterium and Mycobacterium.
Gram-positive bacteria have been reported as being able to produce antibiotics either through ribosomal or nonribosomal mechanisms. In this regard, the orders Bacillales, Lactobacillales and Actinomycetales deserve special mention.
Bacillales
The order Bacillales belongs to the class Bacilli and produces a full range of secondary metabolites with antimicrobial and toxin activities. These compounds are often, but not always, polypeptides. Known antibiotic producers from the Bacillaceae family the synthesis of which is subject to CCR include Bacillus cereus that synthesizes cerexin and zwittermicin; Bacillus circulans and Brevibacillus laterosporus that produce circulin; Paenibacillus polymyxa that produces both polymyxin and colistin; and Brevibacillus brevis that makes gramicidin and tyrothricin. Bacillus licheniformis producing bacitracin, Bacillus pumilus pumulin and Bacillus subtilis makes polymyxin, difficidin, subtilin and mycobacillin. In fact, around 4–5% of the B. subtilis genome is devoted to antibiotic production.45 In this regard, it has been reported that in plant-associated B. amyloliquefaciens FZB42, as much as 8.5% of the genome appears to be devoted to antibiotic production.46
Glucose is the preferred substrate in B. subtilis, and its presence represses the utilization of other alternative substrates. On the basis of physiological and transcriptional data from coutilization experiments with eight different carbon substrates, Kleijn et al.47 demonstrated that malate is a second preferred carbon source for B. subtilis, which is rapidly coutilized with glucose and strongly represses the uptake of alternative substrates.
The mechanism of CCR in B. subtilis and other Firmicutes such as Staphylococcus, Streptococcus, Enterococcus and Lactobacillus, differs from that of enterobacteria. These bacteria essentially possess the same protein constituents of the PTS as are found in E. coli. Therefore, the PTS components form a protein phosphorylation cascade, which uses phosphoenol pyruvate as a phosphoryl donor. However, these Gram-positive bacteria possess a bifunctional enzyme, the HPr kinase/phosphatase (HprK/P), which can be activated by several metabolites (fructose 1,6-diphosphate, gluconate-6-P and 2-phosphoglycerate) when grown in the presence of glucose. Activated HprK/P catalyzes phosphorylation of HPr at Ser-46 and dephosphorylation of P-Ser-HPr. Phosphorylated HPr binds to a pleiotropic regulator, CcpA, to allosterically promote catabolite repression and to prevent inducer accumulation by uncoupling sugar transport from H+ symport.25 CcpA functions as a pleiotropic regulator by binding to the so-called catabolite-responsive elements (cre), which are located either upstream in the promoter regions or in open-reading frames.48 It is interesting to note that polyclonal antibodies against CcpA from Bacillus megaterium share antigenic determinants with CcpA in many other Gram-positive bacteria, including bacilli, staphylococci, streptococci, lactic acid bacteria and some actinomycetes.49 In addition, there is also another regulatory protein, CodY, able to repress genes which allow cell movement to places with higher nutrient concentrations, to breakdown these nutrients and transport the resulting products into the cell. The binding of CodY to certain DNA consensus sequences (CodY box), is facilitated by its interaction with GTP or to branched chain amino acids, specially isoleucine and valine. However, the consensus sequences for the binding of CodY to DNA have not been completely established.50
Finally, it has been demonstrated in B. subtilis that subtilin production is controlled by both the culture density (through a quorum-sensing mechanism) and the microbial growth phase.45, 51
Lactobacillales
Lactic acid bacteria display numerous antimicrobial activities; among them, bacteriocins are especially important because of their use as food preservatives.52 Bacteriocins are antimicrobial peptides or proteins synthesized ribosomally (which distinguishes them from enzymatically synthesized peptide antibiotics) by bacteria. They have a specific killing mechanism that usually acts upon closely related species. It is generally accepted that bacteriocins kill cells by permeabilizing the cytoplasmic membrane and generating pores that allow leaking of cellular components.53
Production of bacteriocins involves a quorum-sensing mode of regulation, mediated by a secreted peptide pheromone (induction peptide), a histidine protein kinase and a response regulator. The secreted pheromone, which in most cases is constitutively produced at a low level during exponential growth phase, serves as a means to measure the cell density of a producing culture. At a certain cell density, the induction peptide reaches a critical threshold concentration that triggers a series of phosphorylation reactions on the histidine protein kinase and response regulator, resulting in a phosphorylated response regulator. The latter binds to regulated promoters of the bacteriocin regulon to activate all genes involved in bacteriocin biosynthesis.54 Although this quorum-sensing induction mechanism has been demonstrated for many different systems, there are additional factors to be considered. For instance, Cheigh et al.55 studied nisin Z production by Lactococcus lactis subsp. lactis A164 grown in various carbon sources, and observed that cells grown in sucrose or glucose generated greater cell mass than did those grown with lactose. However, even at high cell density, the culture yielded lower nisin Z activity along with weaker nisZ transcripts. These data indicate that lactose increases transcription of the nisZ structural gene, whereas glucose and fructose reduce nisin production.
In batch culture production of pediocin (another peptide antibiotic) by Pediococcus acidilactici NRRL B-5627, expression is inhibited by increasing glucose concentrations in the growth medium.56 In Streptococcus salivarius, the production of two lantibiotics (with 2.368 and 2.74 kDa), known as salivaricin A2 and salivaricin B (SboB), has been studied. SboB is encoded by an eight-gene cluster and its production can be reduced in agar cultures supplemented with either glucose or lactose at 1% (wt/vol) rather than sucrose, indicating that the SboB locus is susceptible to CCR.57 Furthermore, the bacteriocin bovicin HC5 produced by Streptococcus bovis was also catabolite repressed by glucose at dilution rates higher than 0.6 per h.58
Growth studies with Lactobacillus brevis have shown that this microorganism simultaneously consumes numerous carbon sources and appears to lack the normal hierarchical control of carbohydrate utilization. Analysis of several independent L. brevis isolates indicates that coutilization of xylose and glucose is a common trait for this species. Moreover, carbohydrates that can be used as a single carbon source are simultaneously utilized with glucose. Proteomic and mRNA analyses showed that genes in the xyl operon were expressed in cells grown on xylose or on glucose/xylose mixtures but not in those grown on glucose alone. However, expression levels of XylA and XylB proteins in cells grown on a glucose/xylose mixture were reduced 2.7-fold from that observed in cells grown solely on xylose. These results suggest that regulation of xylose utilization in L. brevis is not as stringently controlled as in other lactic acid bacteria, in which CCR operates to prioritize carbohydrate utilization more rigorously.59
In lactic acid bacteria, Chaillou et al.60 demonstrated that transport and phosphorylation of glucose is undertaken by the mannose PTS (HPr, EI and an EIIMan complex). Mutations in the EIIMan complex inactivate it and elicit loss of the preferential consumption of glucose over other carbon sources, such as lactose in Lactobacillus casei or xylose in Tetragenococcus halophila. The use of mutants affecting mannose PTS expression in S. salivarius has a pleiotropic effect on various metabolic enzymes, as well as on urease activity and on an inducible fructose PTS activity.60 Interestingly, deletion of EIIMan or mpt (mannose-specific PTS) also produces resistance or immunity to different class II bacteriocins.54, 61, 62, 63 In addition, in Streptococcus gordonii, CcpA is implicated in penicillin tolerance both in vitro and in vivo, linking antibiotic survival to bacterial sugar metabolism.64 Consequently, it will be of great interest in future work to analyze the expression of bacteriocin operons with regard to the carbon source used, because it seems possible that the carbon source regulates bacteriocin production by mediating the formation of enzymes involved in modification, immunity or signal transduction.
In addition, a role for CcpA in the transcriptional regulation of the xyl regulon in Lactobacillus pentosus has been demonstrated. However, no information on the relationship between activity of the EIIMan complex and CcpA-dependent CCR mediated by glucose is available for S. salivarius. In addition, in S. gordonii, CcpA is implicated in penicillin tolerance both in vitro and in vivo, linking antibiotic survival to bacterial sugar metabolism.64
On the other hand, CCR in Streptococcus mutans can be independent of CcpA and the serine-phosphorylated HPr functions in concert with particular PTS permeases to prioritize carbohydrate utilization by modulating sugar transport and transcription of catabolic operons.65
Actinomycetales
The Actinomycetales are primarily aerobic and chemo-organotrophic microorganisms with high GC content. They comprise filamentous, often branching and morphologically diverse bacteria.66 Most of them are Gram-positive eubacteria, although some species with complex cell-wall structures do not show clear Gram staining. A great number of these species are saprotrophic, but a few are pathogenic in plants or animals, including humans. Actinomycetales include families such streptomycetaceae, nocardiaceae, corynebacteriaceae, pseudonocardiaceae, etc.,67 and have received special attention either because of their industrial applications or because of their capability for causing human diseases. Owing to their importance as antibiotic producers, streptomycetaceae, nocardiaceae and corynebacteriae deserve special attention.
Streptomyces
The Streptomyces genus has been found in different environments, such as soil, fresh water and seawater. Frequently, these environmental conditions turn out to be complex and adverse. Considering the fact that streptomycetes are nonmotile microbes, they cannot escape and must face changing environmental conditions. To develop, they must compete with other microorganisms for nutrients present in the environment. For this purpose, they have the ability to produce a wide range of secondary metabolites, such as antibacterial and antifungal substances.68
Nutrients in nature are usually present in a wide range of complexities; therefore, microorganisms must have the necessary tools to succeed in their utilization. For this purpose, 819 potentially secreted proteins have been predicted to operate in S. coelicolor. Among them, cellulases/endoglucanases, chitinases/chitosanases, amylases, proteases/peptidases and pectate lyases are of special importance and many of these enzymes also have commercial interest.69 In addition, because of their capability to degrade multiple natural polymers, streptomycetes have an important role in soil ecology. Very often, the complex and changing environmental conditions can produce microbial stress. To confront this, streptomycetes are armed with many proteins (614 for S. coelicolor), which help them survive under adverse conditions. These protective proteins include transporters, drug efflux proteins and hydrolases.69
The complex life cycle of streptomycetes, their high physiological versatility and ability to produce a great variety of secondary metabolites is reflected in their genome size. With sizes of 8.54, 8.7 and 9.03 Mb, the genomes of S. griseus, S. coelicolor and Streptomyces avermitilis, respectively, are among the largest genomes found in the microbial world.70, 71 Moreover, streptomycetes have higher GC content (>70%) than nearly all other organisms. Unlike most other eubacterial chromosomes, the chromosome of this genus is linear with terminal-inverted repeats and covalently bound terminal proteins at the 5′ end. Housekeeping genes are mainly found in the central region of the linear chromosome, whereas nonessential functions are often located near its ends.19 It is in this region where genes coding for secondary metabolites or transposons are located.72
The ability to produce secondary metabolites, including antibiotics and bioactive compounds, is one of the most interesting properties of Streptomyces. This genus produces ∼70% of the clinically useful antibiotics and anticancer agents.73
For antibiotic production, common intracellular intermediates such as amino acids, sugars, fatty acids, terpenes, etc. are condensed into more complex structures by defined biochemical pathways.74 Nearly 5% of the streptomycete genome (between 23 and 30 gene clusters) is devoted to the synthesis of secondary metabolites.75 The highest percentage reported is 6.43% for the S. avermitilis genome, as this microorganism has the highest proportion of predicted secondary metabolite gene clusters of all bacterial genomes sequenced to date.74
Streptomyces antibiotics are manufactured by microbial fermentation. The onset of biosynthesis is responsive to environmental cues including phosphate ions76 and oxygen concentration, the nature and levels of the carbon and nitrogen source, as well as typical variables, such as temperature, light and pH. Several precise regulatory mechanisms are involved in the onset, maintenance and conclusion of antibiotic production.23 Among these mechanisms, carbon source regulation is one of the main factors required for controlling secondary metabolism.
It is well known that several sugars are commonly used as carbon sources for growth and secondary metabolite production, but some of them are preferred by this genus. At the molecular level, the preferential use of one carbon source over the other and the synthesis of secondary metabolites responds either to prevention of transcriptional activation77 or to repression. Repression by the carbon source is commonly known as CCR.78, 79, 80 This is a phenomenon usually caused by glucose, but in different organisms, other rapidly metabolized carbon sources can cause repression and, indeed, sometimes repress the catabolism of glucose itself.23
More than 20 examples of antibiotics are reported to be suppressed by the carbon source. Glucose and other carbohydrates, such as glycerol, maltose, mannose, sucrose and xylose, have been found to interfere with the synthesis of antibiotics (Table 1). For instance, glucose depresses the formation of β-lactam antibiotics and macrocyclic polyketides. Cephamycin C is a β-lactam antibiotic produced by Streptomyces clavuligerus the synthesis of which is hindered by glycerol through the repression of the enzymes cephamycin C synthetase and expandase. Moreover, expandase activity is inhibited by phosphorylated intermediates of glycolysis, such as glucose 6-phosphate and fructose 1–6 bis-phosphate.81 In addition to cephamycin C, S. clavuligerus produces clavulanic acid, a β-lactamase inhibitor. Although structurally related, these two β-lactams are derived from different biosynthetic precursors. Surprisingly, although glycerol abolishes cephamycin C production, it concomitantly increases clavulanic acid formation. Indeed, as this actinomycete cannot utilize glucose,82 glycerol is a good carbon source for clavulanic acid fermentation.83
Macrocyclic polyketides produced by type I and II PKSs are highly significant secondary metabolites because of their clinical applications. Examples produced by type I PKSs comprise rifamycin and erythromycin, useful against mycobacterial infections; FK506, rapamycin and avermectin are used as immunosuppressant, antitumor and veterinary agents, respectively.84 Polyketides produced by type II PKSs include actinorhodin, tetracenomycin, anthracyclines and tetracyclines.84 The production of polyketydes is also suppressed by different carbon sources. For instance, glucose inhibits actinorhodin production in Streptomyces lividans by repressing the synthesis of afsR2 mRNA that encodes for a global regulatory protein involved in the stimulation of secondary metabolite biosynthesis. As expected, no repression is observed when glucose is substituted by glycerol in this microorganism.85 As in S. lividans, this gene is necessary in S. coelicolor for actinorhodin production. Using two-dimensional gel electrophoresis, it has recently been demonstrated that AfsR2 binds to SCO6569, suggesting that this protein is an AfsR2-dependent downregulator for actinorhodin biosynthesis in S. coelicolor.86
Additional examples of antibiotics the production of which is regulated by the carbon source include the aminoglycoside antibiotics (such as streptomycin, kanamycin, istamycin, neomycin and gentamicin) by repression of biosynthetic enzymes.17, 24 Glucose suppresses streptomycin and neomycin production by S. griseus and Streptomyces fradiae, respectively. The mechanism involves mannosidostreptomycinase and alkaline phosphatase repression for streptomycin and neomycin biosynthesis, respectively.87, 88 For gentamicin production, the glucose effect seems to occur at a step beyond antibiotic intermediate 2-deoxystreptamine.89
Actinomycin production by Streptomyces antibioticus is also subject to control by the carbon source. In this microorganism, glucose transcriptionally represses hydroxykynureninase, an enzyme of the antibiotic pathway. In a control culture of S. antibioticus with galactose as the carbon source, the specific mRNA is low during the trophophase and high in the idiophase. When this culture is compared with that in a glucose medium, the mRNA is much lower and accordingly, the increase in enzyme activity observed in the control by 48 h is almost completely repressed by glucose or glycerol even after 96 h incubation.90 In addition to hydroxykynureninase, the phenoxazinone synthase is repressed by glucose. This 88 000 Mr enzyme catalyzes the condensation of 4-methyl-3-hydroxyanthraniloyl pentapeptide to produce actinomycin. Compared with control cells grown in galactose, the specific activity of phenoxazinone synthase and its mRNA levels is decreased in a glucose medium.91
There are more examples of antibiotics the synthesis of which is impaired by the carbon source, but with scarce information about the target enzymes. This is the case for the synthesis of retamycin, an anthracycline antitumoral complex produced by Streptomyces olindensis using a polyketide pathway. This compound can be produced in a chemostat using a phosphate-limited defined medium. Anthracycline production from this microorganism can be repressed by 139 mM glucose. Under these conditions, an increase in the excretion of organic acids (such as pyruvate, citrate, succinate and lactate), with reduction in the biomass yield is observed, suggesting that flux through the glycolytic pathway has an important role in the repression of the biosynthesis of this antitumoral agent.92
Doxorubicin also belongs to the anthracycline family of antitumor compounds. Its synthesis can be hindered by glucose and galactose in Streptomyces peucetius var. caesius, a strain derived by mutation from the daunorubicin producer S. peucetius. The glucose effect is observed when sugar is added initially or after 24 h fermentation, but not when added during the stationary growth phase. In addition, in a resting cell system containing protein-synthesis inhibitors, anthracycline formation is not affected by high glucose concentrations, suggesting a repressive rather than an inhibitory effect as the possible regulatory mechanism.93
Nystatin belongs to the family of aminoglycosides. When Streptomyces noursei American Type Culture Collection 11455 is supplied with glucose as the carbon source, although high nystatin titers are achieved because of high biomass concentration, glucose tends to negatively influence nystatin production.94 Conversely, glycerol supports a high specific growth rate and high nystatin yields and therefore represents a better choice of carbon source than glucose.
Spiramycin is a 16-membered macrolide antibiotic produced by Streptomyces ambofaciens used to treat toxoplasmosis.95 Antibiotic production is regulated by glucose and glycerol.96 Although glycerol increases growth rate and internal adenosine triphosphate (ATP), it depresses spiramycin production. Specific spiramycin production is increased 10-fold by fed-batch cultures with glycerol and ammonium feeding.97 In a batch culture under ammonium depletion, excess glucose led to pyruvate and α-ketoglutarate accumulation and maintained an antibiotic production phase from these acids after glucose exhaustion.98
One of the conclusions from the above-mentioned information is that not all secondary metabolite genes are equally sensitive to carbon source regulation control. For instance, actinorhodin production is sensitive to glucose concentrations in the range of 100 mM; however, higher concentrations are necessary to repress doxorubicin production.93
Mechanism of CCR in Streptomyces
CCR control by PTS
The mechanism for CCR in Streptomyces is not clearly understood and there is probably more than one mechanism involved. The possible mechanisms have been studied primarily in S. coelicolor, by far one of the better known biological models.19, 99 One of these mechanisms involves the PTS described above.100, 101 In S. coelicolor, there are at least four PTS transporters, all of which use the universal PTS energy-coupling enzymes enzyme I (EI, encoded by ptsI) and HPr (encoded by ptsH).102, 103, 104 Two of the transporters have been characterized in detail, and are responsible for the transport of D-fructose and N-acetylglucosamine.102, 105 Glucose is not transported by the PTS, but through the specific GlcP permease, which is probably the main reason why PTS is not involved in glucose repression.106
Recently, a pleiotropic effect of N-acetylglucosamine (GlcNAc) uptake on development and production of the blue-pigmented polyketide antibiotic actinorhodin has been reported in S. coelicolor.107 This effect depends on the nutritional conditions: under famine (poor) conditions, GlcNAc stimulates development and antibiotic production, whereas under feast (rich) conditions, it blocks development.108 In nature, under starvation conditions, the most likely source of GlcNAc is the autolytic degradation of the bacterial cell wall, whereas under rich conditions, the most likely source is chitin. Hence, GlcNAc signals the nutritional status of the habitat and controls the process of programmed cell death that coincides with development. GlcNAc enters the cell and is phosphorylated through the GlcNAc-specific phosphoenolpyruvate-dependent PTS permease complex that is composed of the general phosphotransferases EI, HPr and IIACrr and the specific IIB and IIC components NagF and NagE2, respectively.107 Phosphorylated GlcNAc (GlcNac-6-P) is further deacetylated by NagA (GlcNAc-6-P deacetylase) producing glucosamine-6-phosphate (GlcN-6-P), an allosteric effector of the global transcription factor DasR (deficient in aerial mycelium and spore formation).108 DasR belongs to the GntR family and controls actinorhodin production by binding to an operator site (dre for DasR-responsive element) overlapping the promoter for actII-ORF4, the pathway-specific activator gene for the actinorhodin biosynthetic cluster. In addition, DasR represses nagF and nagE2 genes, which encode the structural IIB and IIC domains of the GlcNac permease. The effector molecule is GlcN-6-P, thus upon binding to GlcN-6-P, DasR is released from its target genes,109 relieving the inhibition of antibiotic production and morphological differentiation. This effect is absent in a mutant defective in GlcNAc transport.109 Therefore, transcription of nagE2 is induced by GlcNAc but repressed by DasR.
As mentioned above, besides its role in the control of primary and secondary metabolism, DasR also controls the onset of morphological differentiation.109, 110 The manner by which it does that is yet unclear, but it is most likely primarily due to a complete upset of primary metabolism rather than the control of developmental genes, such as the bld or whi genes (GP van Wezel, personal communication). As an example, one of the major targets of DasR control besides the pts and nag regulons is the dasABCD chitobiose utilization operon that lies immediately adjacent to (and is oppositely oriented toward) dasR. Similar to the pts genes, dasA is required for sporulation, and dasA deletion mutants have a bald phenotype.111 Extracellular complementation experiments with different bld mutants showed that the dasR mutant is arrested at a very early stage of the developmental program.109 A complete GlcNAc-dependent signaling model was recently proposed for Streptomyces by Nothaft et al.107 from the perspective of the environmental nutritional status to the onset of antibiotic production through DasR. The start of the signaling cascade is the transport of GlcNAc by NagF, and it culminates in the control of pathway-specific activators for antibiotic production (such as ActII-ORF4). Interestingly, both are subject to multilevel control107 as both nagE2 and actII-ORF4 transcription are activated by AtrA, a TetR-family regulator originally identified as an activator of actinorhodin biosynthesis.77
The evidence provided points to the function of PTS proteins as key elements in CCR exerted by fructose and GlcNac in Streptomyces.110 However, deletion of the ptsH gene encoding HPr has no effect on glucose repression of agarase, galactokinase and glycerol kinase, indicating that PTS responds only to the presence of fructose and GlcNac.105, 112, 113 Therefore, at least one additional mechanism is necessary to explain how glucose and other carbon sources exert CCR.
The role of bld and reg1 in CCR
In addition to the PTS system, the analysis of Streptomyces mutants insensitive to CCR has suggested the involvement of the products of a number of genes such as bld and reg1 in the mechanism of CCR.114, 115 In S. coelicolor, the synthesis of antibiotic and the formation of aerial hyphae initiate at approximately the same time during microbial development, and there is some evidence suggesting that both events are coordinated at the molecular level. In this regard, BldB has been characterized as a protein involved in aerial hyphae formation.114 Interestingly, it has been observed that after rapid growth has ceased, culture supernatants become protein rich, supporting the idea that protein secretion is also a stationary-phase phenomenon.116
It is known that on different carbon sources, BldB null mutants have a bald phenotype, that is, lacking an obvious aerial mycelium, are pleiotropically blocked for antibiotic biosynthesis110 and defective in CCR.117, 118, 119 Thus, bldB, which encodes a small protein containing a putative DNA-binding helix-turn-helix motif, seems to link secondary metabolite production and development to carbon source-dependent gene regulation. In addition to bldB mutants from S. coelicolor, other bld mutants from this microorganism that fail to develop normal aerial mycelia and to produce antibiotics have been isolated by ultraviolet irradiation. The mutants are grouped into four distinct phenotypic classes. In each class, the pleiotropic phenotype is due to a single mutation and separately involves the bldA, bldB, bldG and bldH genes. Among them, bldA, bldG and bldH are repressed during growth on media containing glucose or cellobiose. Repression is partially relieved using alternative carbon sources (arabinose, galactose, glycerol, mannitol and maltose) for bldA and bldG mutants and totally relieved for bldH mutants.114 Therefore, all bld mutants isolated, exhibit carbon source-dependent differentiation.
The bldA gene encodes for a tRNA capable of efficient translation of the uncommon UUA (leucine) codon, which is rarely present in the Streptomyces genome as TTA. Interestingly, it has been shown that the gene for a bldA-dependent protease inhibitor proved to be dependent on the TTA-containing pleiotropic regulatory gene adpA.116 adpA, also known as bldH, mediates the effects of bldA on morphological differentiation and antibiotic production.114 With regard to bldG, it has been proposed that this gene encodes a putative anti-anti-sigma factor that might control transcription of both aerial mycelia formation and antibiotic formation.120
Using transposon mutagenesis, Chouayekh et al.121 isolated S. lividans TK24 mutants the α-amylase expression of which was resistant to glucose catabolite repression. Mutant characterization showed interruption of the sblA gene (equivalent to SCO0479 of S. coelicolor). This gene was predicted to encode a bifunctional enzyme of 274 amino acids with phospholipase C-like and phosphoinositide phosphatase activities. In addition, the sblA mutant strain was shown to enter prematurely into the stationary phase when grown on minimal medium containing glucose as the main carbon source. When cultured in a complex glucose-rich liquid medium, the wild-type strain showed diauxy growth, resuming growth after the transition phase. However, the sblA mutant does not resume growth after the transition phase, when cultured in the same medium, and sporulates much earlier than the wild-type strain when plated on solid-rich medium. These phenotypic differences were abolished when glucose was omitted from the rich medium and correlated with reduced ability for glucose uptake of the sblA mutant strain. Cleavage of phosphoinositides by SblA seems necessary to trigger the glucose-dependent renewed growth that follows the transition phase through an unknown mechanism. The temporal control of sblA expression was shown to depend on two operator sites. One site, located just upstream of the −35 promoter region, likely constitutes a repressor-binding site. The other site, located 170 bp downstream of the GTG sblA translational start codon, may be involved in regulation of the degradation of the sblA transcript.
Involvement of glucose kinase in CCR
D-glucose is one of the easiest utilizable carbon sources for growth in the genus Streptomyces. High concentrations of sugar elicit CCR over the use of alternative carbon sources and the synthesis of several secondary metabolites. Glucose repression in S. coelicolor operates at the transcriptional level to repress enzymes involved in the use of glycerol, arabinose, fructose and galactose122 and this effect seems to be due to either intermediates of carbohydrate catabolism, for example, fructose 1,6-diphosphate and glucose 6-phosphate123, 124 or enzymes of the glucose catabolic pathway, such as glucose kinase.125, 126, 127, 128 In this regard, it has been reported that mutants of S. coelicolor resistant to the nonutilizable glucose analog, 2-deoxyglucose (DOG), appear to be generally deficient in glucose repression.122 These mutants (DogR) can utilize glycerol, arabinose, fructose and galactose in the presence of glucose. Of these mutants, 85% cannot grow on glucose. The loss of glucose repression in DogR mutants has been correlated with the absence of an ATP-dependent glucose kinase (Glk) activity.129 Complementation of these mutants with the Glk gene (glkA) restores not only GlkA activity but also DOG sensitivity and partially restores glucose repression.125, 126 Similarly, when the S. coelicolor glkA gene, is introduced into a S. lividans DogR mutant (unable to utilize glucose and the chitinase production of which is resistant to glucose repression), sensitivity to DOG and the ability to utilize glucose are restored, but glucose repression of chitinase production is only partially recovered.128 Interestingly, when S. coelicolor DogR mutants were complemented with the glk gene of the Gram-negative Zymomonas mobilis, Glk activity and glucose utilization are restored, but not glucose repression.130 These results suggested a regulatory role, different from Glk catalytic activity, necessary for CCR in this microorganism. The regulatory role for Glk in glucose repression of S. coelicolor was further supported by the experiments conducted by Kwakman and Postma.127 These authors demonstrated that the CCR of glycerol kinase and agarase is relieved in cultures of DogR mutants, grown in a series of carbon sources, such as galactose and glycerol, which are not metabolized through Glk. These results strongly suggested that catabolite repression is not regulated by the flux through Glk and that the protein itself has a regulatory role in CCR.127
With regard to the possible mechanism by which Glk exerts a regulatory effect on CCR, it has been observed that this enzyme does not contain obvious DNA-binding sites and thus, it is unfeasible that it could directly regulate transcription. Therefore, it has been proposed that GlkA can transmit its CCR signal by interaction with transcription factors.126 These may include pathway-specific regulators such as MalR and GylR, repressors of the maltose and glycerol operons, respectively, or pleiotropic regulators such as BldB.131, 132 To verify such relationship, MalR, GylR and BldB proteins were heterologously overproduced, purified and assayed for interaction with GlkA. This was approached using several means, including surface plasmon resonance, pull-down assays, a peptide library server, immunoprecipitation and in vivo using two-hybrid technology.133 However, these attempts did not allow identification of a GlkA-interacting protein making interaction between Glk and these transcriptional regulators unlikely. On the other hand, apparent binding between Glk and the glucose transporter GlcP has been reported,134 suggesting that a Glk–GlcP complex is required for efficient glucose metabolism.
Implication of SCO2127 in CCR
Thus, there is mounting evidence that Glk itself is not sufficient to elicit CCR in the genus Streptomyces. For instance, it has been previously shown in S. coelicolor that resistance to DOG yields DogR mutants with decreased glucose kinase activity and reversal of glucose repression.122 A derivative of the temperate phage ϕC31 containing glkA alone is not enough to completely restore the wild-type phenotype when used to lysogenize a S. coelicolor glk null mutant with the DogR phenotype. Complete restoration to the wild-type phenotype is observed with the temperate phage ϕC31 KC896 containing both glkA and sco2127 genes.126 Additional evidence has been supplied by Flores et al.135 showing no differences in the Glk levels between the parental strain of Streptomyces kanamyceticus and a mutant derived from this microorganism, insensitive to CCR for α-amylase and kanamycin formation. Therefore, these results suggest the need for additional mechanisms, in addition to Glk, to explain the CCR mechanism exerted by glucose.
In addition to DOG-resistant mutants from S. coelicolor122 and S. lividans,126 DogR mutants have also been isolated from S. peucetius var. caesius. This microorganism, with an 8.16-Mb genome size, is a doxorubicin producer.136 In addition to presenting CCR insensitivity and low Glk activity, DogR mutants also show difficulties in transporting glucose.93 As previously reported for a DogR mutant from S. coelicolor,125 a partial reversion of the DogR phenotype is observed when transforming the mutant with glkA. However, a complete reversion of this phenotype is obtained when the mutant is transformed with the sco2127 gene alone.137, 138 Thus, in the recombinant strain, glucose uptake and Glk activity values are reverted back to the level of the original strain or even higher, and the recombinant regains sensitivity to CCR. These results were surprising considering that the sco2127 region does not seem to encode for either a glucose permease106 or a Glk,126 and also lacks DNA-binding motifs. Dot-blot analysis of the S. peucetius var. caesius DogR mutant suggested that sco2127 encodes a protein, which stimulates transcription of glk and probably that of the glucose permease gene as well.137 In conclusion, these data suggest participation of an integral regulatory system that is initiated by an increase in glucose incorporation and its metabolism, resulting in increased synthesis of catabolites, which may be involved in eliciting CCR in this microorganism. In agreement with this possibility, among several products of glucose metabolism, fructose 1,6 bis-phosphate and phosphoenolpyruvate exert CCR on anthracycline formation in S. peucetius var. caesius123 with fructose 1,6 bis-phosphate being the most effective (Figure 3). Therefore, it is feasible that phosphorylated sugars can mediate CCR. In this regard, it is known that 2-deoxyglucose, but not 3-O-methylglucose, interferes with tylosin production by S. fradiae in a process sensitive to glucose and phosphate. It has been reported that 2-deoxyglucose is translocated and phosphorylated by most bacteria, whereas 3-O-methylglucose is imported but rarely phosphorylated, supporting glucose phosphorylation as a necessary step for CCR.24

Hypothetical model explaining the stimulatory effect of SCO2127 in S. coelicolor. Abbreviations: bldB, gene encoding for aerial hyphae formation aporepressor; GlcP1, glucose permease; Glk, glucose kinase; gylR, gene encoding for glycerol aporepressor; malR, gene encoding for maltose aporepressor; PEP, phosphoenolpyruvate. The interrupted arrow ( ) indicates enzyme activation. (
) indicates enzyme activation. ( ) indicates gene expression.
) indicates gene expression.
The sco2127 region (576 pb) is located upstream of the glkA gene and no evident function has been conferred to its possible expression product. As sco2127 lacks DNA-binding motifs, direct regulation of the glk promoter cannot be expected. Recently, the protein was overexpressed and purified and polyclonal anti-SCO2127 antibodies were produced. The antibodies were able to detect SCO2127 from crude extracts of both S. coelicolor and S. peucetius var. caesius, yielding a single protein band of 34 kDa in western blot analyses.139 SCO2127 and its corresponding ortholog from S. peucetius var. caesius show a 61% identity in amino-acid composition (JK Sohng, personal communication). When the protein was monitored in S. coelicolor cultures grown in the presence of 50 mM glucose, SCO2127 was detected at the beginning of fermentation and during the logarithmic growth phase, but decreased at later stages of microbial growth. Its production during logarithmic growth agrees with its predicted involvement in CCR, as SCO2127 expression is closely linked to glucose concentration present in the culture medium.139 Detection of SCO2127 at the beginning of fermentation may be related to its possible role in carbohydrate transport138 as the seed cultures were grown in the presence of 50 mM mannitol. On the other hand, cultures grown in 100 mM glucose showed SCO2127 even at later stages of microbial growth coinciding with the presence of 40% of the initial available glucose.
It seems reasonable to think that stimulation of both activities allow efficient glucose consumption with the concomitant production of glucose catabolites, compounds presumably involved in eliciting CCR.123 However, a direct effect of SCO2127 on the activation of Glk or GlcP cannot be discarded. In this regard, the apparent binding of Glk to the major glucose transport system of S. coelicolor (GlcP) reported by van Wezel et al.134 might be mediated by SCO2127, increasing the efficiency of glucose metabolism.
Participation of cAMP and CRP in CCR
For a long time, the existence cAMP in Streptomyces was in doubt. However, cAMP and its binding protein are present in this genus. In addition, 3,5-cyclic guanosine monophosphate is also present in streptomycetes.140 There are several examples suggesting the participation of cAMP in CCR. In S. kanamyceticus, cAMP relieves glucose repression of N-acetylkanamycin amidohydrolase.141 In addition, dibutiryl cAMP supplementation to cultures of S. fradiae, under conditions of growth limitation by ferric ammonium citrate, stimulates tylosin production by 25–30%.142 In the same line, variants of this microorganism with high tylosin production contain 20–50% more cAMP than the parental strain.24
Spore germination, aerial mycelium formation and actinorhodin production are also dependent on cAMP in S. coelicolor.143 Disruption of cya, the gene encoding adenylate cyclase in S. coelicolor generated a mutant (BZ1) unable to produce cAMP and defective in aerial mycelium formation.143 Both wild-type and mutant cultures showed acidification of the medium from 7 to 4.5, but only the parent could partially neutralize the medium during formation of aerial mycelia. Addition of exogenous cAMP or the use of a pH 7 buffer allows the mutant to make aerial mycelia and increases actinorhodin production. Conversely, synthesis of undecylprodigiosin did not respond to cAMP in the presence or absence of buffer. This effect fits well with the selective repression of actinorhodin production in cultures grown in high glucose.144 In addition, the existence of CRP in S. coelicolor has been demonstrated. Thus, SCO3571 (crp) encodes for a protein of 224 amino acids that clusters to the CRP subfamily.144 As reported for cya mutants from S. coelicolor,143 strains defective in crp also showed a lack of germination and actinorhodin production. However, synthesis of several glucose-repressive enzymes (such as α-amylase, mannanase, xylanase and cellulose), induced by their specific substrates was glucose repressed in the crp mutant, suggesting that CRP has no role in CCR in S. coelicolor.145
In contrast to cya mutants, addition of exogenous cAMP does not restore the crp mutant phenotype, indicating that CRP participation is not due to a lack of cAMP biosynthesis. Although it has been suggested that S. coelicolor may use a Cya–cAMP–CRP system to trigger morphogenesis and actinorhodin formation,145 further research is necessary to unequivocally demonstrate the mechanism of CRP participation.
Contribution of ATP and adenosine in CCR
Another nucleotide involved in the regulation of secondary metabolite biosynthesis is ATP. Li et al.146 reported the effect of extracellular ATP (exATP) on antibiotic formation by S. coelicolor. When this microorganism is grown in the presence of 10 mM exATP, actinorhodin concentration increases by 90% compared with a culture grown in the absence of the nucleotide. Conversely, only a 23% increase in undecylprodigiosin is observed. These authors also claim that stimulation by exATP is extended to antibiotics produced by other streptomycetes, including S. lividans, S. griseus, Streptomyces violaceoruber and S. avermitilis.146 The authors suggest that exATP acts as a signaling molecule at the bacterial surface leading to intracellular signaling events and induction of transcriptional activators of antibiotic production. However, in higher exATP, cultures exhibit high intracellular ATP levels and a decreased production in both actinorhodin and undecylprodigiosin.
The nucleoside adenosine also seems to have a role in the production of actinorhodin. Adenosine kinase phosphorylates adenosine to produce AMP using ATP as the main phosphoryl donor. In S. lividans mutants lacking adenosine kinase, no AMP is produced and adenosine is accumulated.147 Under these conditions, production of actinorhodin is suppressed in the mutant and a loss of sporulation is observed. Conversely, production of undecylprodigiosin (the red antibiotic) is enhanced. A promoter-probe assay verified the repressive effect of adenosine on transcription of the pathway-specific activator, actII-ORF4 of actinorhodin production and induction of the undecylprodigiosin promoter.147 No connection between the ATP and adenosine effects on antibiotic production are visualized as S. coelicolor cya mutants, unable to produce cAMP, are expected to accumulate intracellular ATP, which might phosphorylate adenosine avoiding repression of actinorhodin production. Conversely, accumulation of intracellular ATP could be expected to occur in a S. lividans adk mutant, favoring cAMP and actinorhodin formation, which is not the case.
Saccharopolyspora
Saccharopolyspora erythraea is used for industrial-scale production of the antibiotic erythromycin A. Derivatives of this antibiotic have a crucial role in medicine for the treatment of infectious diseases and as a gastrointestinal motor stimulator.5 Its chromosome comprises 8.2 Mb pairs, predicted to encode 7264 genes.148 As reported for other actinomycetes, S. erythraea has a circular chromosome. Its genome contains at least 25 gene clusters for production of known or predicted secondary metabolites, and at least 72 genes have been predicted to confer resistance to a range of common antibiotic classes.148 Erythromycin is produced from propionyl-CoA or succinyl-CoA, through a process of assembly that involves at least 28 active sites arranged along three large proteins. This is followed by hydroxylation and glycosilation processes involving 18 additional proteins. Finally, microbial ribosomes are protected from high antibiotic-specific toxicity by specific methylation of the rRNA.149
Glucose transiently represses antibiotic formation150 probably by repressing methylmalonyl-CoA-mutase.151 Although this enzyme is repressed by glucose, methylmalonyl-CoA decarboxylase production is not affected by the carbon source, suggesting that the negative effect of glucose on erythromycin production could be due, in part, to lower pools of succinyl-CoA and methylmalonyl-CoA.151 Recently, engineering S. erythraea through duplication of the methylmalonyl-CoA mutase led to a 50% increase in erythromycin production.152 Moreover, glucose or glycerol inhibits S-adenosylmethionine erythromycin O-methyl transferase activity. Reeve and Baumberg153 measured the effect of glucose on the transcription of the eryAI gene, encoding the type I polyketide synthase. With an increase in glucose concentration, they found a decrease in the lag before the onset of erythromycin production, but a decrease in the final levels of eryAI expression.
Substitution of glucose by cane molasses accompanied by a reduction in ammonium sulfate concentration and supplementation of the medium with n-propanol has been utilized for increasing erythromycin production and reduction in costs of antibiotic production.154
In S. erythraea, the erythromycin biosynthetic cluster lacks a regulatory gene.155 However, a 17.7-kDa bldD ortholog has recently been described, which positively regulates all promoters in the erythromycin production cluster, suggesting the existence of a transcriptional activator of the ery gene cluster.155 In S. coelicolor, BldD negatively regulates the expression of key developmental genes.156 bldD mutants pleiotropically affect both formation of aerial hyphae and antibiotic production in this microorganism.157 Similarly, S. erythraea bldD mutants produce a bald phenotype and seven-fold less erythromycin than the S. erythraea wild-type strain (NRRL2338).155
Nocardia
Bacteria from the genus nocardia produce a wide variety of secondary metabolites. Nocardicin A and B (Nocardia sp.), ryfamicin (Nocardia mediterranea), ansamitocin (Nocardia brasiliensis), 3-O-demethyl mutactimycin (Nocardia transvalensis), neo-nocardin (Nocardia kuroishi) and cephamycin C (Nocardia lactamdurans) are some examples of antibiotics produced by this genus.
Although reported as a good antibiotic producer, studies on the effect of carbon source on idiolite formation are limited in this genus. For instance, it has been reported that various sugars and their metabolites inhibit cephamycin C production by N. lactamdurans resting cells.158 At the biochemical level, glucose-6-phosphate and fructose-1,6-diphosphate inhibit deacetoxycephalosporin C (DAOC) synthase, one of the pathway enzymes dealing with antibiotic formation. With regard to cAMP levels, a similar behavior to that reported for several streptomycetes is observed, that is, high during growth and low during antibiotic production, suggesting that this nucleotide is not involved in derepressing secondary metabolism.
Corynebacterium
Corynebacterium kutscheri and Corynebacterium xerosis produce antimicrobials effective against bacteria and fungi. It has been established that antibiotic production in these species is greatly influenced by variation in the carbon source. Along this line, ribose and lactose suppress the antimicrobial activity of C. kutscheri and C. xerosis, respectively.159
The corynebacteria have genes for the PTS-regulatory mechanism of CCR. Two energy-coupling proteins, E1 and Hpr, and permeases for glucose and fructose are present. Moreover, it contains two distinct genes encoding an IIABPtx-like and a novel HPr-type protein of unknown function. In addition, a possible PTS gene target was elucidated as a possible regulatory gene encoding an antiterminator protein.160
Carbon source regulation of secondary metabolism biosynthesis in fungi
As observed for actinomycetes, fungi also synthesize antibiotics in response to physiological stresses, such as nutrient limitation (carbon, nitrogen or phosphate sources). The growth medium markedly influences the production of antibiotics.161 In addition, the carbon source may exert complex regulation on gene expression and enzyme activities for polyketide synthesis.162
Table 2 shows some examples of antibiotics produced by fungi that are repressed by different carbon sources, and their target enzymes.
Penicillins and cephalosporins are β-lactam antibiotics. The formation of hydrophobic penicillins has been reported in fungi only, notably Penicillium chrysogenum and A. nidulans, whereas the hydrophilic cephalosporins are produced by both fungi, for example, Acremonium chrysogenum (cephalosporin C) and bacteria.163
In P. chrysogenum, glucose represses the transcription of penicillin biosynthetic genes pcbAB, pcbC and penDE164 which encode for δ-(L-α-aminoadipyl)-L-cysteinyl-D-valine synthetase (ACVS), isopenicillin N-synthase and IPN acyltransferase, respectively. Whereas glucose negatively affects pcbAB, pcbC and penDE gene promoters, alkaline pH exerts a small positive effect on these promoters.164 The P. chrysogenum pcbC gene was cloned downstream of the strong, inducible Hansenula polymorpha alcohol oxidase promoter (PAOX) and was integrated at the PAOX locus in the H. polymorpha genome and cultivated on methanol medium to induce the PAOX and then to mid-exponential growth phase. When glucose (0.5%) was added to the cultures, a treatment that results in complete repression of the PAOX (30 min after glucose addition), the isopenicillin N-synthase protein levels in the cells decreased by >50%.165
In the same manner, genes aatA and ipnA but not acvA (homologs to penDE, pcbC and pcbAB, respectively) are repressed by glucose in A. nidulans.166, 167 A Cre consensus sequence 5′-SYGGRG-3′, is located in the upstream region of the ipnA gene and the binding of CreA (CCR) has been confirmed in vitro.168 However, it has been shown that A. nidulans creA mutants cultured in glucose, still present repression of ipnA transcription.169
CreA is the major repressor protein regulating the CCR of genes involved in carbon metabolism in A. nidulans.170 It binds to the consensus DNA sequence 5′-SYGGRG-3′ (where S could be C or G; Y could be C or T and R could be A or G) in promoters of glucose-repressible genes, turning off their expression. Cre-binding sites are found in the promoters of cre genes from diverse filamentous fungi.171
The promoters of many glucose-repressible target genes have Cre-binding consensus sequences.172 The Cre1 protein (from the cre1 gene) sequence shows high similarity to other glucose-repressor proteins from filamentous fungi, suggesting a similar regulation and a related role in CCR. Janus et al.171 identified a 114-bp minimal promoter with two Cre1-binding sites, the first minimal regulatory promoter sequence for the β-lactam antibiotic-producing fungus. The identification of 114-bp CT-rich sequences in the nontranscribed cre1 upstream region suggests that it is required to trigger glucose-dependent gene expression.171
Mutations in creB and creC have shown little effect on carbon regulation of penicillin biosynthesis. All these results point to a second mechanism of carbon repression, which is creA independent.173
In A. chrysogenum, glucose represses cephalosporin C formation. As this organism contains cre1- and cre1-binding sites upstream of the isopenicillin N-synthase (pcbC) and the DAOC/deacetylcephalosporin C synthase (cefEF) genes, it is considered that cre1 might be involved in repression of antibiotic formation. Indeed, in the wild-type strain, glucose increased the level of cre1 transcripts sixfold. As expected, in a commercial high-producing A. chrysogenum strain, glucose did not affect the cre1 transcript levels. Therefore, it is feasible that cre1 might be involved in glucose repression of cephalosporin C production and that during the program of strain improvement, this control mechanism could have been deregulated.171, 174 CT-rich regions are putative activator sequences in close proximity to transcription start sites.175 In A. chrysogenum, the CT-rich region contains two closely spaced binding sites (namely BS14 and BS15) that cooperatively bind to Cre1. The cre1 promoter from A. chrysogenum displays a positive autoregulation upon glucose induction. This 114-bp minimal promoter with two Cre1 consensus binding sites is an example of a minimal regulatory promoter sequence for this β-lactam antibiotic-producing fungus.171
Direct inhibition (a mechanism that restrains synthesis at the level of enzyme activity, therefore the effect is immediate) of the action of secondary metabolic enzymes may also be involved in carbon source control. Thus, glucose and a number of its phosphorylated metabolites inhibit ACVS, the first enzyme of cephalosporin synthesis, in crude extracts of A. chrysogenum and S. clavuligerus.176 As no inhibition is observed with the purified enzyme, it seems that this phenomenon might be caused by competition for ATP between primary (Embden–Meyerhof pathway) and secondary metabolism. Accordingly, it has been found that inhibition of ACVS by sugars can be prevented by addition of more ATP. Conversely, both the crude and the purified forms of ACVS are inhibited by glyceraldehyde-3-phosphate (G3P). This effect seems to be due to the ability of G3P to chemically complex and remove cysteine, a substrate of ACVS. The in vivo significance of such inhibitory phenomena remains to be determined.
With regard to the mechanism of CCR in A. nidulans, carbon catabolism is a tightly regulated process, whereby the most energetically favorable carbon sources are used preferentially over less readily metabolized carbon sources. It has been demonstrated that a Glk (GlkA4) and a hexokinase (Hxk) participate equally in sugar signaling. Single glkA4 and hxkA mutants were able to substitute for each other in their function in catalysis and in CCR. However, double mutants in glkA4 and hxkA genes showed derepression in genes, such as facA for acetate catabolism, xlnA for xylan degradation and alcA and alcR for ethanol utilization.177 These observations suggest that glucose phosphorylation could be the relevant signal for glucose repression in A. nidulans.177 In addition, the creA, creB, creC and cre1 genes178 were identified as coding for regulatory proteins involved in CCR in A. nidulans. CreA and Cre1 contain two Cys2His2 DNA-binding zinc fingers similar to the Saccharomyces cerevisiae glucose-repressor protein Mig1.170, 179, 180 creB encodes a deubiquitinating enzyme, and creC encodes a WD40 protein that contains five WD40 repeats and a proline-rich region. CreC forms a complex with CreB.181, 182 The proposed model for CCR through CreA modification or stability involves the CreB deubiquitination enzyme complexing with the CreC protein to modify or stabilize CreA under carbon-repressing conditions.183
An additional gene implicated in the mechanism of A. nidulans CCR is creD. Mutations in this gene suppress the creB15 and creC27 mutant phenotypes, showing derepression of facA and alcA genes. The creD gene encodes a protein that contains arresting domains and PY motifs and interacts with the HECT (homologous to the E6-associated protein carboxyl terminus) ubiquitine ligase.184 Although the elements of ubiquitination and deubiquitination networks have been disclosed, their direct targets still remain to be elucidated.170 Recently, it was found that the mechanism of action of CreA in CCR does not require its degradation nor its cellular localization, but seems to entail some modification or interaction with another protein to act as a repressor.170 In a study elucidating the kinetics of glucose uptake in A. nidulans, two energy-requiring glucose transport systems were identified: a high-affinity glucose-repressible system and a low-affinity glucose inducible system (MstE). Besides glucose, mstE expression is induced in the presence of other repressing carbon sources, and is dependent on the function of the transcriptional repressor CreA.185
Pleuromutilin is a diterpenoid antibiotic produced by Pleurotus mutilis-04. Pleuromutilin is currently used as an intermediate for the production of tiamulin and valnemulin for veterinary use. In addition, retapamulin is a promising antibiotic with the potential for human use in the treatment of Gram-positive bacterial infections.186 Compared with glucose as the sole carbon source, soybean oil was beneficial for pleuromutilin production in shaker flask cultures.187 This result suggested a suppressive effect of glucose on antibiotic production.
Fusarium fujikuroi produces the red pigment bikaverin, which displays antibiotic activity against some protozoa and phytopathogenic fungi.188 Antibiotic production occurs during the stationary growth phase, after exhaustion of the nitrogen source. After 12 days of incubation, antibiotic production was stimulated by sucrose, but strongly suppressed in the presence of equivalent carbon concentrations, up to 8% glucose, by glucose, galactose, fructose, maltose, xylose and glycerol.189 Sucrose is converted to glucose and fructose by invertase activity; however, neither fructose nor a mixture of glucose and fructose reproduced the sucrose stimulatory effect observed in F. fujikuroi, suggesting that the inducing signal is sucrose itself, and not a sucrose-derived metabolite.
Strain improvement by avoiding or removing CCR
Although it has been more than 60 years since the first penicillin-overproducing mutant was obtained by X-ray treatment of P. chrysogenum X-1612,190 conventional breeding (random mutagenesis and massive tracing of strains) remains the preferred method when molecular genetic tools have not been developed for the producer microorganism.191
Nutritional repression can be decreased by mutation to antimetabolite resistance. An example of a selective agent is DOG, utilized for enzymes and secondary metabolites, the synthesis of which is controlled by carbon source regulation. This is illustrated by the generation of spontaneous mutants from S. peucetius var. caesius resistant to DOG.136 When one of these mutants is recombined with a mutant isolated as resistant to growth inhibition, as determined by high doxorubicin concentrations, the recombinant offspring show resistance to the mentioned compounds, insensitivity to CCR and improvement in antibiotic formation.192
Although random mutagenesis and screening procedures have been widely used for genetic improvement in antibiotic production, there are certain disadvantages, such as the time necessary to obtain a favorable mutation. Thus, several rounds of improvement are normally required, before a good increase in antibiotic production is observed. In addition, there are risks of unwanted mutations resulting in crippled strains, and random approaches may not always produce the optimal solution in terms of specific mutations that are required for the desirable phenotype. However, there is a great potential in the use of metabolic engineering combined with advances in the biochemical knowledge and understanding of the biosynthetic pathways to successfully improve antibiotic production. On the basis of this, antibiotic production can be improved by metabolic engineering in several ways. A flux increase in the biosynthetic pathway can be improved by directed mutagenesis to elevate precursor availability or reduce by-product formation.193 For instance, by cloning acetyl-CoA carboxylase into an expression vector and introducing it into a wild-type strain of S. coelicolor, the conversion of acetyl-CoA into malonyl-CoA was enhanced and channeled to actinorhodin production.194 In a similar work, deletion of the pfkA2 gene, corresponding to one of the three reported homologs of phosphofructokinase in S. coelicolor, increased actinorhodin and undecylprodigiosin production approximately four times as compared with the wild-type strain.124 In addition, it is known that G3P and L-arginine are precursors in the biosynthesis of clavulanic acid in S. clavuligerus. G3P is converted into 1,3-diphosphoglicerate (1,3-BPG) by G3P dehydrogenases. These enzymes are encoded by genes gap1 and gap2 in this microorganism. Therefore, to improve clavulanic acid production, the glycolitic pathway has been genetically engineered by targeted gene disruption of both gap genes. The procedure allowed accumulation of G3P and an increase in antibiotic production of more than twofold when L-arginine was fed to the cultures.195
We have already mentioned some key enzymes for secondary metabolism that are regulated or repressed by the carbon source (Table 1). A few of these enzymes have been used as targets for strain improvement. For instance, in the case of cephalosporin production, a lead enzyme, DAOC synthase (or expandase), which is affected by CCR, has been chosen as a target to increase production of these antibiotics. An initial approach, carried out by Basch and Chiang196 in an A. chrysogenum strain, increased the copy number of the expandase gene (cefEF) encoding for DAOC synthetase. In the resulting transformants, they observed a 40–70% reduction in deacetoxycephalosporin C levels and an increase in the cephalosporin C titer. A more recent study shows an increase in cephalosporin production based on the disruption of the cefEF gene of A. chrysogenum, followed by cloning and one-step replacement with the cefE gene from S. clavuligerus. Through this procedure, recombinant strains were obtained producing high titers of penicillin N, which is converted to DAOC. Deacylation of DAOC is carried out by two final enzymatic bioconversions catalyzed by D-amino acid oxidase and glutaryl acylase yielding 7-aminocephalosporanic acid, precursor of all clinically important semi-synthetic cephalosporin derivatives.197
In S. clavuligerus, expandase was modified by site-directed mutagenesis to promote the conversion of penicillin G to 7-phenylacetylamino-deacetoxycephalosporonic acid, precursor of 7-aminodeacetoxycephalosporonic acid. The resulting mutants showed increased kcat/Km values and 13-fold increase in relative activity toward penicillin G.198 Addition of different side chains to 7-aminodeacetoxycephalosporonic acid converts this compound into the oral cephalosporins: cephradine, cephalexin and cephadroxil.
S-adenosyl-L-methionine (SAM) is most commonly used in living systems as a methyl donor involved in methylation reactions. The synthesis of SAM in the cell is solely catalyzed by an enzyme called SAM synthetase (MetK, ATP: L-methionine S-adenosyltransferase, EC 2.5.1.6). Overexpression of SAM especially enhances the production of a number of secondary metabolites, including actinorhodin and calcium-dependent antibiotics in S. coelicolor, pristinamycin in Streptomyces pristinaespiralis, granaticin in S. violaceoruber, oleandomycin in S. antibioticus, avermectin in S. avermitilis and doxorubicin in S. peucetius var. caesius.199, 200, 201, 202 Although the overall mechanism underlying this activity is not completely understood, the obvious methylation of intermediates leading to the biosynthesis of antibiotics involving SAM provides a strong explanation for the high yield of these compounds.203
There have been notorious advances in the elucidation of the regulatory mechanisms of antibiotic formation at the biochemical and molecular levels. Thorough investigation of these mechanisms could allow us to increase the efficiency of metabolic pathways involved in antibiotic production, making the genetic improvement of industrial strains a more efficient and oriented task, compared with classical strategies.204
Concluding remarks
The outstanding role of microorganisms in the production of antibiotics is notorious. By far, most of the industrially important antibiotics are produced by the genus Streptomyces. The complex life cycle, high physiological versatility and ability to produce a great variety of secondary metabolites by this genus, is reflected in its large genome size. Antibiotics are usually formed during the late growth phase through enzymatic pathways carrying out various enzymatic steps. They are often brought on by exhaustion of nutrients, a situation that generates signals that cause a cascade of regulatory events resulting in chemical and morphological differentiation. Among the nutrients, the effect of the carbon source on secondary metabolite production has been the subject of this review.
Regulation by the carbon source depends on the rapid utilization of the preferred carbon source. Different mechanisms have been described in bacteria and fungi to explain the negative carbon catabolite effects on antibiotic production. These mechanisms show important differences depending on the microbe being considered.
With the available information, some proteins and protein complex systems have been found to be crucial for the carbon source effect on antibiotic biosynthesis. In addition, direct carbon source inhibition of key enzymes involved in secondary metabolite pathways has also been documented.
For a desired antibiotic to become a commercial reality, the genetic improvement in its production is a necessary step before getting the required titers for industrial production. For every new strain developed genetically, there are further opportunities to increase its titers. The relevance of the understanding of CCR mechanisms will provide not only scientific profit, but also rational strategies for selection of secondary metabolite overproducing strains. Both strain improvement and nutritional modification will preclude the scaling up into the pilot scale by biochemical engineers and later, into factory size fermentors.
References
Demain, A. L. & Fang, A. The natural functions of secondary metabolites. in Advances in Biochemical Engineering/Biotechnology: History of Modern Biotechnology I (ed. Fietcher, A.) Vol. 69, 2–39 (Springer, Berlin, 2000).
Berdy, J. Bioactive microbial metabolites. A personal view. J. Antibiot. 58, 1–26 (2005).
Lederberg, J. Infectious history. Science 288, 287–293 (2000).
Barber, M., Giesecke, U., Reichert, A. & Minas, W. Industrial enzymatic production of cephalosporin-based β-lactams. Adv. Biochem. Eng./Biotechnol. 88, 179–215 (2004).
Demain, A. L. & Sanchez, S. Microbial drug discovery: 80 years of progress. J. Antibiot. 62, 5–16 (2009).
Cardenas, M. E., Sanfridson, A., Cutler, N. S. & Heitman, J. Signal-transduction cascades as targets for therapeutic intervention by natural products. Trends Biotechnol. 16, 427–433 (1998).
Kremer, L., Douglas, J. D., Baulard, A. R., Morehouse, C. & Guy, M. R. Thiolactomycin and related analogues as novel anti-mycobacterial agents targeting KasA and KasB condensing enzymes in Mycobacterium tuberculosis. J. Biol. Chem. 275, 16857–16864 (2000).
Demain, A. L. Prescription for an ailing pharmaceutical industry. Nat. Biotechnol. 20, 331 (2002).
Minotti, G., Menna, P., Salvatorelli, E., Cairo, G. & Gianni, L. Anthracyclines: molecular advances and pharmacologic developments in antitumor activity and cardiotoxicity. Pharmacol. Rev. 56, 185–229 (2004).
Ikeda, F. et al. Role of micafungin in the antifungal armamentarium. Curr. Med. Chem. 14, 1263–1275 (2007).
Kirst, H. A. et al. Evaluation and development of spinosyns to control ectoparasites on cattle and sheep. Curr. Top. Med. Chem. 2, 675–699 (2002).
Cragg, G. M. & Newman, D. J. Medicinals for the millennia. The historycal record. Ann. NY Acad. Sci. 953a, 3–25 (2001).
Yang, S. S., Cragg, G. M., Newman, D. J. & Bader, J. P. Natural products based anti-HIV drug discovery and development facilitated by the NCI developmental therapeutics program. J. Nat. Prod. 64, 265–277 (2001).
Sanchez, S. & Olson, B. The bright and promising future of microbial manufacturing. Curr. Opin. Microbiol. 8, 229–233 (2005).
Paradkar, A., Trefzer, A., Chakraburtty, R. & Stassi, D. Streptomyces genetics: a genomic perspective. Crit. Rev. Biotechnol. 23, 1–27 (2003).
Demain, A. L. Induction of microbial secondary metabolism. Int. Microbiol. 1, 259–264 (1998).
Piepersberg, W. & Distler, J. Aminoglycosides and sugar components in other secondary metabolites. in Products of Secondary Metabolism. (eds Rehm, H.J., Reed, G.) 397–488 (VCH-Verlagsgesells-chaft, Weinheim, 1997).
Martínez-Bueno, M., Gálvez, A., Valdivia, E. & Maqueda, M. A transferable plasmid associated with AS-48 production in Enterococcus faecalis. J. Bacteriol. 172, 2817–2818 (1990).
Hopwood, D. A. Forty years of genetics with Streptomyces: from in vivo through in vitro to in silico. Microbiology 145, 2183–2202 (1999).
Brown, D. W. et al. Twenty-five coregulated transcripts define a sterigmatocystin gene cluster in Aspergillus nidulans. Proc. Natl Acad. Sci. USA 93, 1418–1422 (1996).
Bibb, M. J. Regulation of secondary metabolism in streptomycetes. Curr. Opin. Microbiol. 8, 208–215 (2005).
Ohnishi, Y., Yamazaki, H., Kato, J., Tomono, A. & Horinouchi, S. AdpA, a central transcriptional regulator in the A-factor regulatory cascade that leads to morphological development and secondary metabolism in Streptomyces griseus. Biosci. Biotechnol. Biochem. 69, 431–439 (2005).
Sanchez, S. & Demain, A. L. Regulation of fermentation processes. Enzyme Microb. Technol. 31, 895–906 (2002).
Demain, A. L. Carbon source regulation of idiolite biosynthesis. in Regulation of Secondary Metabolism in Actinomycetes. (ed. Shapiro, S.) 127–134 (CRC Press, Boca Raton, FL, 1989).
Deutscher, J. The mechanisms of carbon catabolite repression in bacteria. Curr. Opin. Microbiol. 11, 87–93 (2008).
Görke, B. & Stülke, J. Carbon catabolite repression in bacteria: many ways to make the most out of nutrients. Nat. Rev. Microbiol. 6, 613–624 (2008).
Coulthurst, S. J., Barnard, A. M. L. & Salmond, G. P. C. Regulation and biosynthesis of carbapenem antibiotics in bacteria. Nat. Rev. Microbiol. 3, 295–306 (2005).
McGowan, S. et al. Carbapenem antibiotic production in Erwinia carotovora is regulated by CarR, a homologue of the LuxR transcriptional activator. Microbiology 141, 541–550 (1995).
Veselova, M. et al. Production of N-acyl homoserine lactone signal molecules by Gram-negative soil-borne and plant-associated bacteria. Folia Microbiol. 48, 794–798 (2003).
Derzelle, S., Duchaud, E., Kunst, F., Danchin, A. & Bertin, P. Identification, characterization, and regulation of a cluster of genes involved in carbapenem biosynthesis in Photorhabdus luminescens. Appl. Environ. Microbiol. 68, 3780–3789 (2002).
Cang, S. et al. High production of prodigiosin in Serratia marcesens grown on ethanol. Biotech. Lett. 22, 1761–1765 (2000).
Thomson, N. R., Crow, M. A., McGowan, S. J., Cox, A. & Salmond, G. P. Biosynthesis of carbapenem antibiotic and prodigiosin pigment in Serratia is under quorum sensing control. Mol. Microbiol. 36, 539–556 (2000).
Williamson, N. R., Fineran, P. C., Leeper, F. J. & Salmond, G. P. C. The biosynthesis and regulation of bacterial prodiginines. Nat. Rev. Microbiol. 4, 887–899 (2006).
Fineran, P. C., Slater, H., Everson, L., Hughes, K. & Salmond, G. P. C. Biosynthesis of tripyrrole and β-lactam secondary metabolites in Serratia: integration of quorum sensing with multiple new regulatory components in the control of prodigiosin and carbapenem antibiotic production. Mol. Microbiol. 56, 1495–1517 (2005).
Clardy, J. & Brady, S. F. Cyclic AMP directly activates NasP, an N-acyl amino acid antibiotic biosynthetic enzyme cloned from an uncultured β-Proteobacterium. J. Bacteriol. 189, 6487–6489 (2007).
Isnansetyo, A., Cui, L., Hiramatsu, K. & Kamei, Y. Antibacterial activity of 2, 4-diacetylphloroglucinol produced by Pseudomonas sp. AMSN isolated from a marine alga, against vancomycin-resistant Staphylococcus aureus. Int. J. Ant. Agents 22, 545–547 (2003).
Chin-A-Woeng, T. F. C., Bloemberg, G. V. & Lugtenberg, B. J. J. Phenazines and their role in biocontrol by Pseudomonas bacteria. New Phytol. 157, 503–523 (2003).
van Rij, E. T., Wessenlink, M., Chin-A-Woeng, T. F., Bloemberg, G. V. & Lugtenberg, B. J. Influence of environmental conditions on the production of phenazine-1-carboxamide by Pseudomonas chlororaphis PCL 1391. Mol. Plant Microbe Interact. 17, 557–566 (2004).
Li, Y., Jiang, H., Xu, Y. & Zhang, X. Optimization of nutrient components for enhanced phenazine1-carboxylic acid production by gacA-inactivated Pseudomonas sp. M18G using response surface method. Appl. Microbiol. Biotechnol. 77, 1207–1217 (2008).
Sonnleitner, E., Abdou, L. & Haas, D. Small RNA as global regulator of carbon catabolite repression in Pseudomonas aeruginosa. Proc. Natl Acad. Sci. USA 106, 21866–21871 (2009).
Moreno, R., Marzi, S., Romby, P. & Rojo, F. The Crc global regulator binds to an unpaired A-rich motif at the Pseudomonas putida alkS mRNA coding sequence and inhibits translation initiation. Nucleic. Acids Res. 37, 7678–7690 (2009).
Kay, E., Dubuis, C. & Haas, D. Three small RNAs jointly ensure secondary metabolism and biocontrol in Pseudomonas fluorescens CHA0. Proc. Natl Acad. Sci. USA 102, 17136–17141 (2005).
Ludwig, W., Schleifer, K. H. & Whitman, W. B. Revised road map to the phylum Firmicutes. in The Firmicutes. Bergey's Manual of Systematic Bacteriology. 2nd edn, Vol 3. (eds Vos, P., Garrity, G., Jones, D., Krieg, N.R., Ludwig, W., Rainey, F.A., et al.) (Springer-Verlag, New York, 2009).
Garrity, G. M. et al. Part 10–The Bacteria: Phylum ‘Actinobacteria’: Class Actinobacteria. Taxonomic Outline of the Bacteria and Archaea, Release 7.7. 399–539 (Michigan State University Board of Trustees, East Lansing, MI, 2007).
Stein, T. Bacillus subtillis antibiotics: structure, syntheses and specific functions. Mol. Microbiol. 56, 845–857 (2005).
Chen, X. H. et al. Comparative analysis of the complete genome sequence of the plant growth–promoting bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 25, 1007–1014 (2007).
Kleijn, R. J. et al. Metabolic fluxes during strong carbon catabolite repression by malate in Bacillus subtilis. J. Biol. Chem. 285, 1587–1596 (2009).
Lulko, A. T., Buist, G., Kok, J. & Kuipers, O. P. Transcriptome analysis of temporal regulation of carbon metabolism by CcpA in Bacillus subtilis reveals additional target genes. J. Mol. Microbiol. Biotechnol. 12, 82–95 (2007).
Küster, E., Luesink, E. J., de Vos, W. M. & Hillen, W. Immunological crossreactivity to the catabolite control protein CcpA from Bacillus megaterium is found in many Gram-positive bacteria. FEMS Microbiol. Lett. 139, 109–115 (2006).
Sonenshein, A. L. Control of key metabolic intersections in Bacillus subtilis. Nat. Rev. Microbiol. 5, 917–927 (2007).
Fawcett, P., Eichenberger, P., Losick, R. & Youngman, P. The transcriptional profile of early to middle sporulation in Bacillus subtilis. Proc. Natl Acad. Sci. USA 97, 8063–8068 (2000).
Ganzle, M. G. From gene to function: metabolic traits of starter cultures for improved quality of cereal foods. Int. J. Food Microbiol. 134, 29–36 (2009).
Mantovani, H. C. & Russell, J. B. Bovicin HC5, a lantibiotic produced by Streptococcus bovis HC5, catalyzes the efflux of intracellular potassium but not ATP. Antimicrob. Agents Chemother. 52, 2247–2249 (2008).
Diep, D. B., Mathiesen, G., Eijsink, V. G. H. & Nes, I. F. Use of lactobacilli and their pheromone-based regulatory mechanism in gene expression and drug delivery. Curr. Pharm. Biotechnol. 10, 62–73 (2009).
Cheigh, C. I., Park, H., Choi, H. J. & Pyun, Y. R. Enhanced nisin production by increasing genes involved in nisin Z biosynthesis in Lactococcus lactis subsp. lactis A164. Biotechnol. Lett. 27, 155–160 (2005).
Pérez-Guerra, N., Bernárdez, P. J. & Pastrana Castro, L. P. Fed-batch pediocin production on whey using different feeding media. Enzyme Microb. Technol. 41, 397–406 (2007).
Hyink, O. et al. Salivaricin A2 and the novel lantibiotic salivaricin B are encoded at adjacent loci on a 190-kilobase transmissible megaplasmid in the oral probiotic strain Streptococcus salivarius K12. Appl. Environ. Microbiol. 73, 1107–1113 (2007).
Mantovani, H. C. & Russell, J. B. Factors affecting the antibacterial activity of the ruminal bacterium, Streptococcus bovis HC5. Curr. Microbiol. 46, 18–23 (2003).
Kim, J. H., Shoemaker, S. P. & Mills, D. A. Relaxed control of sugar utilization in Lactobacillus brevis. Microbiology 155, 1351–1359 (2009).
Chaillou, S., Postma, P. W. & Pouwels, P. H. Contribution of the phosphoenolpyruvate: mannose phosphotransferase system to carbon catabolite repression in Lactobacillus pentosus. Microbiology 147, 671–679 (2001).
Kowalczyk, M. & Bardowski, J. Regulation of sugar catabolism in Lactococcus lactis. Crit. Rev. Microbiol. 33, 1–13 (2007).
Saulnier, D. M. A., Molenaar, D., de Vos, W. M., Gibson, G. R. & Kolida, S. Identification of prebiotic fructooligosaccharide metabolism in Lactobacillus plantarum WCFS1 through microarrays. Appl. Environ. Microbiol. 73, 1753–1765 (2007).
Andersson, U., Molenaar, D., Radstrom, P. & de Vos, W. M. Unity in organisation and regulation of catabolic operons in Lactobacillus plantarum, Lactococcus lactis and Listeria monocytogenes. Syst. Appl. Microbiol. 28, 187–195 (2005).
Bizzini, A., Entenza, J. M. & Moreillon, P. Loss of penicillin tolerance by inactivating the carbon catabolite repression determinant CcpA in Streptococcus gordonii. J. Antimicrob. Chemother. 59, 607–615 (2007).
Zeng, L. & Burne, R. A. Seryl-phosphorylated HPr regulates CcpA-independent carbon catabolite repression in conjunction with PTS permeases in Streptococcus mutans. Mol Microbiol. 75, 1145–1158 (2010).
Madigan, M. M., Martinko, J. & Parker, J. Brock Biology of Microorganisms. 10th edn. (Prentice Hall, Upper Saddle River, NJ, 2003).
Garrity, G. M., Bell, J. A. & Lilburn, T. G. Taxonomic outline of the prokaryotes. Bergey's Manual of Systematic Bacteriology. (Springer-Verlag, New York, 2004).
Xiong, L., Li, J. & Kong, F. Streptomyces sp. 173, an insecticidal micro-organism from marine. Lett. Appl. Microbiol. 38, 32–37 (2004).
Bentley, S. D. et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 417, 141–147 (2002).
Ohnishi, Y. et al. Genome sequence of the streptomycin-producing microorganism Streptomyces griseus IFO 13350. J. Bacteriol. 190, 4050–4060 (2008).
Weber, T., Welzel, K., Pelzer, S., Vente, A. & Wohlleben, W. Exploiting the genetic potential of polyketide producing streptomycetes. J. Biotechnol. 106, 221–232 (2003).
Mochizuki, S. et al. The large linear plasmid pSLA2-L of Streptomyces rochei has an unusually condensed gene organization for secondary metabolism. Mol. Microbiol. 48, 1501–1510 (2003).
Demain, A. L. Pharmaceutically active secondary metabolites of microorganisms. Appl. Microbiol. Biotechnol. 52, 455–463 (1999).
Omura, S. et al. Genome sequence of an industrial microorganism Streptomyces avermitilis: deducing the ability of producing secondary metabolites. Proc. Natl Acad. Sci. USA 98, 12215–12220 (2001).
Ikeda, H. et al. Complete genome sequence and comparative analysis of the industrial microorganism Streptomyces avermitilis. Nat. Biotechnol. 21, 526–531 (2003).
Martin, J. F. Phosphate control of the biosynthesis of antibiotics and other secondary metabolites is mediated by the PhoR-PhoP system: an unfinished story. J. Bacteriol. 186, 5197–5201 (2004).
Uguru, G. C. et al. Transcriptional activation of the pathway-specific regulator of the actinorhodin biosynthetic genes in Streptomyces coelicolor. Mol. Microbiol. 58, 131–150 (2005).
Hodgson, D. A. Primary metabolism and its control in streptomycetes: a most unusual group of bacteria. Adv. Microb. Physiol. 42, 47–238 (2000).
Brückner, R. & Titgemeyer, F. Carbon catabolite repression in bacteria: choice of the carbon source and autoregulatory limitation of sugar utilization. FEMS Microbiol. Lett. 209, 141–148 (2002).
Titgemeyer, F. & Hillen, W. Global control of sugar metabolism: a Gram-positive solution. Anton. Leeuw. Int J. G. 82, 59–71 (2002).
Lebrihi, A., Lefebvre, G. & Germain, P. Carbon catabolite regulation of cephamycin C and expandase biosynthesis in Streptomyces clavuligerus. Appl. Microbiol. Biotechnol. 28, 44–51 (1988).
Garcia-Dominguez, M., Martin, J. F. & Liras, P. Characterization of sugar uptake in wild-type Streptomyces clavuligerus, which is impaired in glucose uptake, and in a glucose-utilizing mutant. J. Bacteriol. 171, 6808–6814 (1989).
Saudagar, P. S. & Singhal, R. S. Optimization of nutritional requirements and feeding strategies for clavulanic acid production by Streptomyces clavuligerus. Biores. Technol. 98, 2010–2017 (2007).
Lal, R. et al. Regulation and manipulation of the gene clusters encoding type-I PKSs. Trends Biotechnol. 18, 264–274 (2000).
Kim, E. S., Hong, H., Choi, C. Y. & Cohen, S. N. Modulation of actinorodin biosynthesis in Streptomyces lividans by glucose repression of afsR2 gene transcription. J. Bacteriol. 183, 2198–2203 (2001).
Im, J. H., Lee, M. J., Lee, S. Y. & Kim, E. S. A putative secreted solute binding protein, SCO6569 is a possible AfsR2-dependent down-regulator of actinorhodin biosynthesis in Streptomyces coelicolor. Process Biochem. 44, 373–377 (2009).
Demain, A. L. & Inamine, E. Biochemistry and regulation of streptomycin and mannosidostreptomycinase (α-D-mannosidase) formation. Bacteriol. Rev. 34, 1–19 (1970).
Bandyopadhyay, S. K. & Majumdar, S. K. Regulation of the formation of alkaline phosphatase during neomycin biosynthesis. Antimicrob. Agents Chemother. 5, 431–434 (1974).
Escalante, L., González, R., Obregón, A. M. & Sánchez, S. Carbon catabolite regulation of gentamicin formation. J. Antibiot. 45, 757–761 (1992).
Brown, D. D., Hitchcock, M. J. M. & Katz, E. Evidence for a constitutive and inducible form of kynurenine formamidase in an actinomycin-producing strain of Streptomyces parvulus. Arch. Biochem. Biophys. 202, 18–22 (1980).
Jones, G. H. Regulation of phenoxazinone synthase expression in Streptomyces antibioticus. J. Bacteriol. 163, 1215–1221 (1985).
Inoue, O., Netto, W. S., Padilla, G. & Facciotti, M. C. R. Carbon catabolite repression of retamycin production by Streptomyces olindensis Icb20. Brazilian J. Microbiol. 38, 58–61 (2007).
Escalante, L., Ramos, I., Imriskova, I., Langley, E. & Sanchez, S. Glucose repression of anthracycline formation in Streptomyces peucetius var. caesius. Appl. Microbiol. Biotechnol. 52, 572–578 (1999).
Jonsbu, E., McIntyre, M. & Nielsen, J. The influence of carbon sources and morphology on nystatin production by Streptomyces noursei. J. Biotechnol. 95, 133–144 (2002).
Karray, F. et al. Organization of the biosynthetic gene cluster for the macrolide antibiotic spiramycin in Streptomyces ambofaciens. Microbiology 153, 4111–4122 (2007).
Lounès, A., Lebrihi, A., Benslimane, C., Lefebvre, G. & Germain, P. Regulation of spiramycin synthesis in Streptomyces ambofaciens: effects of glucose and inorganic phosphate. Appl. Microbiol. Biotechnol. 45, 204–211 (1996a).
Lounès, A., Lebrihi, A., Benslimane, C., Lefebvre, G. & Germain, P. Effect of nitrogen/carbon ratio on the specific production rate of spiramycin by Streptomyces ambofaciens. Proc. Biochem. 31, 13–20 (1996b).
Colombié, V., Bideaux, C., Goma, G. & Uribelarrea, J. L. Effects of glucose limitation on biomass and spiramycin production by Streptomyces ambofaciens. Bioprocess. Biosyst. Eng. 28, 55–61 (2005).
Chater, K. F. Streptomyces inside out: a new perspective on the bacteria that provide us with antibiotics. Philos. Trans. R. Soc. Lon. B Siol. Sci. 361, 761–768 (2006).
Saier, Jr M. H. Multiple mechanisms controlling carbon metabolism in bacteria. Biotechnol. Bioeng. 58, 170–174 (1998).
Stülke, J. & Hillen, W. Carbon catabolite repression in bacteria. Curr. Opin. Microbiol. 2, 195–201 (1999).
Titgemeyer, F. et al. Identification and characterization of phosphoenolpyruvate: fructose phosphotransferase systems in three Streptomyces species. Microbiology 141, 51–58 (1995).
Butler, M. J. et al. Analysis of a ptsH homologue from Streptomyces coelicolor A3(2). FEMS Microbiol. Lett. 177, 279–288 (1999).
Parche, S., Schmid, R. & Titgemeyer, F. The phosphotransferase system (PTS) of Streptomyces coelicolor identification and biochemical analysis of a histidine phosphocarrier protein HPr encoded by the gene ptsH. Eur. J. Biochem. 265, 308–317 (1999).
Wang, F., Xiao, X., Saito, A. & Schrempf, H. Streptomyces olivaceoviridis possesses a phosphotransferase system that mediates specific, phosphoenolpyruvate-dependent uptake of N-acetylglucosamine. Mol. Genet. Genomics 268, 344–351 (2002).
van Wezel, G. P. et al. GlcP constitutes the major glucose uptake system of Streptomyces coelicolor A3(2). Mol. Microbiol. 55, 624–636 (2005).
Nothaft, H. et al. The permease gene nagE2 is the key to N-acetylglucosamine sensing and utilization in Streptomyces coelicolor and is subject to multi-level control. Mol. Microbiol. 75, 1133–1144 (2010).
Rigali, S. et al. Feast or famine: the global regulator DasR links nutrient stress to antibiotic production by Streptomyces. EMBO Rep. 9, 670–675 (2008).
Rigali, S. et al. The sugar phosphotransferase system of Streptomyces coelicolor is regulated by the GntR-family regulator DasR and links N-acetylglucosamine metabolism to the control of development. Mol. Microbiol. 61, 1237–1251 (2006).
Seo, J.- W., Ohnishi, Y., Hirata, A. & Horinouchi, S. ATP-binding cassette transport system involved in regulation of morphological differentiation in response to glucose in Streptomyces griseus. J. Bacteriol. 184, 91–103 (2002).
Colson, S. et al. The chitobiose-binding protein, DasA, acts as a link between chitin utilization and morphogenesis in Streptomyces coelicolor. Microbiology 154, 373–382 (2008).
Jahreis, K., Pimentel-Schmitt, E. F., Brückner, R. & Titgemeyer, F. Ins and outs of glucose transport systems in eubacteria. FEMS Microbiol. Rev. 32, 891–907 (2008).
Nothaft, H. et al. The phosphotransferase system of Streptomyces coelicolor is biased for N-acetylglucosamine metabolism. J. Bacteriol. 185, 7019–7023 (2003).
Champness, W. C. New loci required for Streptomyces coelicolor morphological and physiological differentiation. J. Bacteriol. 170, 1168–1174 (1988).
Nguyen, J. The regulatory protein Reg1 of Streptomyces lividans binds the promoter region of several genes repressed by glucose. FEMS Microbiol. Lett. 175, 51–58 (1999).
Kim, D. W., Chater, K., Lee, K. J. & Hesketh, A. Changes in the extracellular proteome caused by the absence of the bldA gene product, a developmentally significant tRNA, reveal a new target for the pleiotropic regulator AdpA in Streptomyces coelicolor. J. Bacteriol. 187, 2957–2966 (2005).
Kelemen, G. H. & Buttner, M. J. Initiation of aerial mycelium formation in Streptomyces. Curr. Opin. Microbiol. 1, 656–662 (1998).
Pope, M. K., Green, B. & Westpheling, J. The bldB gene encodes a small protein required for morphogenesis, antibiotic production, and catabolite control in Streptomyces coelicolor. J. Bacteriol. 180, 1556–1562 (1998).
Eccleston, M., Ali, R. A., Seyler, R., Westpheling, J. & Nodwell, J. Structural and genetic analysis of the BldB protein of Streptomyces coelicolor. J. Bacteriol. 184, 4270–4276 (2002).
Bignell, D. R. D., Warawa, J. L., Strap, J. L., Chater, K. F. & Leskiw, B. K. Study of the bldG locus suggests that an anti-anti-sigma factor and an anti-sigma factor may be involved in Streptomyces coelicolor antibiotic production and sporulation. Microbiol. 146, 2161–2173 (2000).
Chouayekh, H. et al. Phosphoinositides are involved in control of the glucose-dependent growth resumption that follows the transition phase in Streptomyces lividans. J. Bacteriol. 189, 741–749 (2007).
Hodgson, D. A. Glucose repression of carbon source uptake and metabolism in Streptomyces coelicolor A3(2) and its perturbation in mutants resistant to 2-deoxyglucose. J. Gen. Microbiol. 128, 2417–2430 (1982).
Ramos, I. et al. Glucose kinase alone cannot be responsible for carbon source regulation in Streptomyces peucetius var. caesius. Res. Microbiol. 155, 267–274 (2004).
Borodina, I. et al. Antibiotic overproduction in Streptomyces coelicolor A3(2) mediated by phosphofructokinase deletion. J. Biol. Chem. 283, 25186–25199 (2008).
Ikeda, H., Seno, E. T., Bruton, C. J. & Chater, K. F. Genetic mapping, cloning and physiological aspects of the glucose kinase gene of Streptomyces coelicolor. Mol. Gen. Genet. 196, 501–507 (1984).
Angell, S., Schwarz, E. & Bibb, M. J. The glucose kinase gene of Streptomyces coelicolor A3(2): its nucleotide sequence, transcriptional analysis and role in glucose repression. Mol. Microbiol. 6, 2833–2844 (1992).
Kwakman, J.H.J.M. & Postma, P. W. Glucose kinase has a regulatory role in carbon catabolite repression in Streptomyces coelicolor. J. Bacteriol. 176, 2694–2698 (1994).
Saito, A., Fugii, T., Yoneyama, T. & Miyashita, K. glkA is involved in glucose repression of chitinase production in Streptomyces lividans. J. Bacteriol. 180, 2911–2914 (1998).
Seno, E. T. & Chater, K. F. Glycerol catabolic enzymes and their regulation in wild-type and mutant strains of Streptomyces coelicolor A3(2). J. Gen. Microbiol. 129, 1403–1413 (1983).
Angell, S., Lewis, C. G., Buttner, M. J. & Bibb, M. L. Glucose repression in Streptomyces coelicolor A3(2) a likely regulatory role for glucose kinase. Mol. Gen. Genet. 244, 135–143 (1994).
Pope, M. K., Green, B. D. & Westpheling, J. The bld mutants of Streptomyces coelicolor are defective in the regulation of carbon utilization, morphogenesis and cell-cell signalling. Mol. Microbiol. 19, 747–756 (1996).
van Wezel, G. P., White, J., Young, P., Postma, P. W. & Bibb, M. J. Substrate induction and glucose repression of maltose utilization by Streptomyces coelicolor A3(2) is controlled by malR, a member of the lacI-galR family of regulatory genes. Mol. Microbiol. 23, 537–549 (1997).
Mahr, K. et al. Glucose kinase of Streptomyces coelicolor A3(2): large-scale purification and biochemical analysis. Anton. Leeuw. Int. J.G. 78, 253–261 (2000).
van Wezel, G. P. et al. A new piece of an old jigsaw: glucose kinase is activated posttranslationally in a glucose transport-dependent manner in streptomyces coelicolor A3(2). J. Mol. Microbiol. Biotechnol. 12, 67–74 (2007).
Flores, M. E., Ponce, E., Rubio, M. & Huitrón, C. Glucose repression of α-amylase in Streptomyces kanamyceticus and isolation of deregulated mutants. Biotechnol. Lett. 15, 595–600 (1993).
Segura, D. et al. Streptomyces mutants insensitive to glucose repression showed deregulation of primary and secondary metabolism. ASPA J. Mol. Biol. Biotechnol. 4, 30–36 (1996).
Guzmán, S. et al. Pleiotropic effect of the sco2127 gene on the glucose uptake, glucose kinase activity and carbon catabolite repression in Streptomyces peucetius var. caesius. Microbiology 151, 1717–1723 (2005a).
Guzmán, S. et al. Sugar uptake and sensitivity to carbon catabolite regulation in Streptomyces peucetius var. caesius. Appl. Microbiol. Biotechnol. 69, 200–206 (2005b).
Chávez, A., Garcia-Huante, Y., Ruiz, B., Rodriguez-Sanoja, R. & Sanchez, S. Cloning and expression of the sco2127 gene from Streptomyces coelicolor M145. J. Ind. Microbiol. Biotechnol. 36, 649–654 (2009).
Gersch, D., Romer, W., Bocker, H. & Thrum, H. Variations in cyclic adenosine 3′,5′-monophosphate and cyclic guanosine 3′,5′-monophosphate in antibiotic producing strains of Streptomyces hygroscopicus. FEMS Microbiol. Lett. 3, 39–41 (1978).
Satoh, A., Ogawa, H. & Satomura, Y. Regulation of N-acetylkanamycin amidohydrolase in the idiophase in kanamycin fermentation. Agr. Biol. Chem. 40, 191–196 (1976).
Tata, M. & Menawat, A. S. Cyclic AMP regulation of tylosin biosynthesis and secondary metabolism in Streptomyces fradiae. Biotechnol. Bioeng. 44, 283–290 (1994).
Süsstrunk, U., Pidoux, J., Taubert, S., Ullmann, A. & Thompson, C. J. Pleiotropic effects of cAMP on germination, antibiotic biosynthesis, and morphological development in Streptomyces coelicolor. Mol. Microbiol. 30, 33–46 (1998).
Kang, S. G., Jin, W., Bibb, M. & Lee, K. J. Actinorhodin and undecylprodigiosin production in wild-type and relA mutant strains of Streptomyces coelicolor A3(2) grown in continuous culture. FEMS Microbiol. Lett. 168, 221–226 (1998).
Derouaux, A. et al. Deletion of a cyclic AMP receptor protein homologue diminishes germination and affects morphological development of Streptomyces coelicolor. J. Bacteriol. 186, 1893–1897 (2004).
Li, M., Kim, T., Kwon, H. & Suh, J. Effects of extracellular ATP on the physiology of Streptomyces coelicolor A3(2). FEMS Microbiol. Lett. 286, 24–31 (2008).
Rajkarnikar, A., Kwon, H. J. & Suh, J. W. Role of adenosine kinase in the control of Streptomyces differentiations: loss of adenosine kinase suppresses sporulation and actinorhodin biosynthesis while inducing hyperproduction of undecylprodigiosin in Streptomyces lividans. Biochem. Biophys. Res. Comm. 363, 322–328 (2007).
Oliynyk, M. et al. Complete genome sequence of the erythromycin-producing bacterium Saccharopolyspora erythraea. Nat. Biotechnol. 25, 428–429 (2007).
Challis, G. & Hopwood, D. Synergy and contingency as driving forces for the evolution of multiple secondary metabolite production by Streptomyces species. Proc. Natl Acad.Sci.USA 100, 14555–14561 (2003).
Escalante, L., Lopez, H., Mateos, R. C., Lara, F. & Sanchez, S. Transient repression of erythromycin formation in Streptomyces erythraeus. J. Gen. Microbiol. 128, 2011–2015 (1982).
Bermudez, O., Padilla, P., Huitron, C. & Flores, M. E. Influence of carbon and nitrogen source on synthesis of NADP super(+)-isocitrate dehydrogenase, methylmalonyl-coenzyme A mutase, and methylmalonyl-coenzyme A decarboxylase in Saccharopolyspora erythraea CA340. FEMS Microbiol. Lett. 164, 77–82 (1988).
Reeves, A. R., Brikun, I. A., Cernota, W. H., Leach, B. I., Gonzalez, M. C. & Weber, J. M. Engineering of the methylmalonyl-CoA metabolite node of Saccharopolyspora erythraea for increased erythromycin production. Metab. Eng. 9, 293–303 (2007).
Reeve, L. M. & Baumberg, S. Physiological controls of erythromycin production by Saccharopolyspora erythraea are exerted at least in part at the level of transcription. Biotechnol. Lett. 20, 585–589 (1998).
El-Enshasy, H. A., Mohamed, N. A., Farid, M. A. & El-Diwany, A. L. Improvement of erythromycin production by Saccharopolyspora erythraea in molasses based medium through cultivation medium optimization. Biores. Technol. 99, 4263–4268 (2008).
Chng, C., Lum, A. M., Vroom, J. A. & Kao, C. M. A key developmental regulator controls the synthesis of the antibiotic erythromycin in Saccharopolyspora erythraea. Proc. Natl Acad. Sci. USA 105, 11346–11351 (2008).
Elliot, M. A., Bibb, M. J., Buttner, M. J. & Leskiw, B. K. BldD is a direct regulator of key developmental genes in Streptomyces coelicolor A3(2). Mol. Microbiol. 40, 257–269 (2001).
Elliot, M. A., Locke, T. R., Galibois, C. M. & Leskiw, B. K. BldD from Streptomyces coelicolor is a non-essential global regulator that binds its own promoter as a dimer. FEMS Microbiol. Lett. 225, 35–40 (2003).
Cortes, J., Castro, J. M., Liras, P. & Martin, J. F. Glucose regulation of cephamycin biosynthesis in Streptomyces lactamdurans. Proc. 3rd Eur. Congr. Biotechnol. München, Germany 1, I 21–I 27 (1984).
El-Banna, N. M. Effect of carbon source on the antimicrobial activity of Corynebacterium kutscheri and Corynebacterium xerosis. Afr. J. Biotechnol. 5, 833–835 (2006).
Parche, S., Thomae, A. W., Schlicht, M. & Titgemeyer, F. Corynebacterium diphtheriae: a PTS view to the genome. J. Mol. Microbiol. Biotechnol. 3, 415–422 (2001).
Papagianni, M. Fungal morphology and metabolite production in submerged mycelial processes. Biotechnol. Adv. 22, 189–259 (2004).
Jia, Z., Zhang, X. & Cao, X. Effects of carbon sources on fungal morphology and lovastatin biosynthesis by submerged cultivation of Aspergillus terreus. Asia Pac. J. Chem. Eng. 4, 672–677 (2009).
Brakhage, A. A. et al. Aspects on evolution of fungal β-lactam biosynthesis gene clusters and recruitment of trans-acting factors. Phytochemistry 70, 1801–1811 (2009).
Gutiérrez, S. et al. Transcription of the pcbAB, pcbC and penDE genes of Penicillium chrysogenum AS-P-78 is repressed by glucose and the repression is not reversed by alkaline pHs. Microbiology 145, 317–324 (1999).
Gidijala, L. et al. Production of functionally active Penicillium chrysogenum isopenicillin N synthase in the yeast Hansenula polymorpha. BMC Biotechnol. 19, 8–29 (2008).
Brakhage, A. A. et al. Regulation of penicillin biosynthesis in filamentous fungi. Adv. Biochem. Eng. Biotechnol. 88, 45–90 (2004).
Litzka, O., Bergh, K. T. & Brakhage, A. A. Analysis of the regulation of the Aspergillus nidulans penicillin biosynthesis gene aat (penDE), which encodes acyl coenzyme A:6-aminopenicillanic acid acyltransferase. Mol. Gen. Genet. 249, 557–569 (1995).
Espeso, E. A. & Peñalva, M. A. In vitro binding of the two-finger repressor CreA to several consensus and non-consensus sites at the ipnA upstream region is context dependent. FEBS Lett. 342, 43–48 (1994).
Espeso, E. A. & Peñalva, M. A. Carbon catabolite repression can account for temporal pattern of expression of a penicillin biosynthetic gene in Aspergillus nidulans. Mol. Microbiol. 6, 1457–1465 (1992).
Roy, P., Lockington, R. A. & Kelly, J. M. CreA-mediated repression in Aspergillus nidulans does not require transcriptional auto-regulation, regulated inracellular localisation or degradation. Fungal Gen. Biol. 45, 657–670 (2008).
Janus, D., Hortschansky, P. & Kück, U. Identification of a minimal cre1 promoter sequence promoting glucose-dependent gene expression in the beta-lactam producer Acremonium chrysogenum. Curr. Genet. 53, 35–48 (2008).
Rauscher, R. et al. Transcriptional regulation of xyn1, encoding xylanase I, in Hypocrea jecorina. Eukaryot Cell 5, 447–456 (2006).
Espeso, E. A., Fernández-Cañón, J. M. & Peñalva, M. A. Carbon regulation of penicillin biosynthesis in Aspergillus nidulans: a minor effect of mutations in creB and creC. FEMS Microbiol. Lett. 126, 63–68 (1995).
Jekosch, K. & Kück, U. Glucose dependent transcriptional expression of the cre1 gene in Acremonium chrysogenum strains showing different levels of cephalosporin C production. Curr. Genet. 37, 388–395 (2000a).
Neveu, B., Belzile, F. & Bélanger, R. R. Cloning of the glyceraldehyde-3-phosphate dehydrogenase gene from Pseudozyma xocculosa and functionality of its promoter in two Pseudozyma species. Anton. Leeuw. Int J. G. 92, 245–255 (2007).
Zhang, J. & Demain, A. L. Regulation of ACV synthetase activity in the beta-lactam biosynthetic pathway by carbon sources and their metabolites. Arch. Microbiol. 158, 364–369 (1992).
Flipphi, M. et al. Onset of carbon catabolite repression in Aspergillus nidulans: parallel involvement of hexokinase and glucokinase in sugar signaling. J. Biol. Chem. 278, 11849–11857 (2003).
Arst, H. N., Tollervey, Jr D., Dowzer, C. E. A. & Kelly, J. M. An inversion truncating the creA gene of Aspergillus nidulans results in carbon catabolite derepression. Mol. Microbiol. 4, 851–854 (1990).
Cubero, B. & Scazzochio, C. Two different, adjacent and divergent zinc finger binding sites are necessary for CreA mediated carbon catabolite repression in the proline gene cluster of Aspergillus nidulans. EMBO J. 13, 407–411 (1994).
Cubero, B., Gomez, D. & Scazzocchio, C. Metabolite repression and inducer exclusion in the proline utilization gene cluster of Aspergillus nidulans. J. Bacteriol. 182, 233–235 (2000).
Todd, R. B., Lockington, R. A. & Kelly, J. M. The Aspergillus nidulans creC gene involved in carbon catabolite repression encodes a WD40 repeat protein. Mol. Gen. Genet. 263, 561–570 (2000).
Lockington, R. A. & Kelly, J. M. Carbon catabolite repression in Aspergillus nidulans involves deubiquitination. Mol. Microbiol. 40, 1311–1321 (2001).
Lockington, R. A. & Kelly, J. M. The WD40-repeat protein CreC interacts with and stabilizes the deubiquitinating enzyme CreB in vivo in Aspergillus nidulans. Mol. Microbiol. 43, 1173–1182 (2002).
Boase, N. A. & Kelly, J. M. A role for creD, a carbon catabolite repression gene from Aspergillus nidulans, in ubiquitination. Mol. Microbiol. 53, 929–940 (2004).
Forment, J. V., Flipphi, M., Ramón, D., Ventura, L. & MacCabe, A. P. Identification of the mstE gene encoding a glucose-inducible, low affinity glucose transporter in Aspergillus nidulans. J. Biol. Chem. 281, 8339–8346 (2006).
Phillips, O. A. & Sharaf, L. H. Pleuromutilin antibacterial agents: patent review 2001–2006. Exp. Op. Therap. Pat. 17, 429–436 (2007).
Hu, C., Zou, Y. & Zhao, W. Effect of soybean oil on the production of mycelial biomass and pleuromutilin in the shake-flask culture of Pleurotus mutilis. World J. Microbiol. Biotechnol. 25, 1705–1711 (2009).
Son, S. W. et al. Bikaverin and fusaric acid from Fusarium oxysporum show antioomycete activity against Phytophthora infestans. J. Appl. Microbiol. 104, 692–698 (2008).
Rodríguez-Ortiz, R., Mehta, B. J., Avalos, J. & Limón, M. C. Stimulation of bikaverin production by sucrose and by salt starvation in Fusarium fujikuroi. Appl. Microbiol. Biotechnol. 85, 1991–2000 (2010).
Adrio, J. L. & Demain, A. L. Genetic improvement of process yielding microbial products. FEMS Microbiol. Rev. 30, 187–214 (2006).
Khetan, A. & Hu, W. S. Metabolic engineering of antibiotic biosynthetic pathways. in Manual of Industrial Microbiology and Biotechnology (eds Demain, A.L., Davies, J.E., Atlas, R.M., Cohen, G., Hershberger, C., Hu, W.S., et al.) 717–724 (ASM Press, Washington, D.C., 1999).
Segura, D., Santana, C., Gosh, R., Escalante, L. & Sanchez, S. Anthracyclines: isolation of overproducing strains by selection and genetic recombination of putative regulatory mutants of Streptomyces peucetius var. caesius. Appl. Microbiol. Biotechnol. 48, 615–620 (1997).
Rokem, J. S., Eliasson, A. & Nielsen, J. Systems biology of antibiotic production by microorganisms. Nat. Prod. Rep. 24, 1262–1287 (2007).
Ryu, Y. G., Butler, J. M., Chater, F. K. & Lee, J. K. Engineering of primary carbohydrate metabolism for increased production of actinorhodin in Streptomyces coelicolor. Appl. Environ. Microbiol. 72, 7132–7139 (2006).
Li, R. F. & Townsend, C. A. Rational strain improvement for enhanced clavulanic acid production by genetic engineering of the glycolytic pathway in Streptomyces clavuligerus. Metab. Eng. 8, 240–252 (2006).
Basch, J. & Chiang, S. Genetic engineering approach to reduce undesirable by-products in cephalosporin C fermentation. J. Ind. Microbiol. Biotechnol. 20, 344–353 (1998).
Velazco, J., Adrio, J. L., Moreno, M. A., Diez, B., Soler, G. & Barredo, J. L. Environmental safe production of 7-aminodeacetoxycephalosporanic acid (7-ADCA) using recombinant strains of Acremonoium chrysogenum. Nat. Biotechnol. 18, 857–861 (2000).
Wei, C. L. et al. Engineering Streptomyces clavuligerus deacetoxycephalosporin C synthase for optimal ring expansion activity toward penicillin G. Appl. Environ. Microbiol. 69, 2306–2312 (2003).
Kim, D. J. et al. Accumulation of S-adenosyl-L-methionine enhances production of actinorhodin but inhibits sporulation in Streptomyces lividans TK23. J. Bacteriol. 185, 592–600 (2003).
Okamoto, S., Lezhava, A., Hosaka, T., Okamoto-Hosoya, Y. & Ochi, K. Enhanced expression of S-adenosylmethionine synthetase causes overproduction of actinorhodin in Streptomyces coelicolor A3(2). J. Bacteriol. 185, 601–609 (2003).
Huh, J. H. et al. Widespread activation of antibiotic biosynthesis by S-adenosylmethionine in Streptomycetes. FEMS Microbiol. Lett. 238, 439–447 (2004).
Oh, T. J., Niraula, N. P., Liou, K. & Sohng, J. K. Identification of the duplicated genes for S-adenosyl-L-methionine synthetase (metK1-sp and metK2-sp) in Streptomyces peucetius var caesius ATCC 27952. J. Appl. Microb 108, 398–407 (2010).
Kim, W. S., Wang, Y., Fang, A. & Demain, A. L. Methionine interference in rapamycin production involves repression of demethylrapamycin methyltransferase and S-adenosylmethionine synthetase. Antimicrob. Agents Chemother. 44, 2908–2910 (2000).
Demain, A. L. & Adrio, J. L. Strain improvement for production of pharmaceuticals and other microbial metabolites by fermentation. Prog. Drug Res. 65, 252–289 (2008).
Brown, D. D. et al. Regulation of tryptophan metabolism and its relationship to actinomycin D-synthesis. in Genetics of Industrial Microorganisms (eds Ikeda, Y. & Beppu, T.) 85–91 (Kodansha, Tokyo, 1983).
Bhatnagar, R. K., Doull, J. L. & Vining, L. C. Role of the carbon source in regulating chloramphenicol production by Streptomyces venezuelae: studies in batch and continuous cultures. Can. J. Microbiol. 34, 1217–1223 (1988).
Sankaran, L. & Pogell, P. M. Biosynthesis of puromycin in Streptomyces alboniger: regulation and properties of O-demethylpuromycin O-methyltransferase. Antimicrob. Agents Chemother. 8, 721–732 (1975).
Erban, V., Novotná, J., Bìhal, V. & Hoštálek, Z. Growth rate, sugar consumption, chlortetracycline production and anhydrotetracycline oxygenase activity in Streptomyces aureofaciens. Folia Microbiol. 28, 262–267 (1983).
Vu-Trong, K., Bhuwapathanapun, S. & Gray, P. P. Metabolic regulation in tylosin-producing Streptomyces fradiae: regulatory role of adenylate nucleotide pool and enzymes involved in biosynthesis of tylonolide precursors. Antimicrob. Agents Chemother. 17, 519–525 (1980).
Jekosch, K. & Kuck, U. Loss of glucose repression in an Acremonium chrysogenum β-lactam producer strain and its restoration by multiple copies of the cre1 gene. Appl. Microbiol. Biotechnol. 54, 556–563 (2000b).
Theilgaard, H.B., Kristiansen, K. N., Henriksen, C. M. & Nielsen, J. Purification and characterization of delta-(L-alpha-aminoadipyl)-L-cysteinyl-D-valine synthetase from Penicillium chrysogenum. Biochem. J. 327, 185–191 (1997).
Acknowledgements
We are indebted to MA Ortíz and L Escalante for their assistance during the elaboration of this work. We thank GP van Wezel for reviewing a section of this paper. This work was supported in part by the grants IN 209210 from PAPIIT, Dirección General de Asuntos del Personal Académico, UNAM and CB2008-100564-IIBO from Consejo Nacional de Ciencia y Tecnología, Mexico.
Author information
Authors and Affiliations
Corresponding author
Additional information
This work was written to honor Professor Arnold L Demain, on the 60th anniversary of his very productive scientific and personal life. During this time, he has been strongly devoted to the study of secondary metabolism and has guided more than 100 students throughout the industrial microbiology and biotechnology world. In 1995, most of these students gathered to celebrate the First Arny's Army and Friends Symposium at MIT in the United States, an academic event that has been repeated every 2 years for the past 15 years.
Rights and permissions
About this article
Cite this article
Sánchez, S., Chávez, A., Forero, A. et al. Carbon source regulation of antibiotic production. J Antibiot 63, 442–459 (2010). https://doi.org/10.1038/ja.2010.78
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ja.2010.78
Keywords
This article is cited by
-
Utilizing Waste from Palm Oil Mills as a Substrate to Produce Antimicrobial Agents Using Fermentation by Streptomyces philanthi RM-1-138: Effectiveness, Stability, and Compound Identification
Waste and Biomass Valorization (2024)
-
Exploring the biosynthetic gene clusters in Brevibacterium: a comparative genomic analysis of diversity and distribution
BMC Genomics (2023)
-
From waste to health-supporting molecules: biosynthesis of natural products from lignin-, plastic- and seaweed-based monomers using metabolically engineered Streptomyces lividans
Microbial Cell Factories (2023)
-
Functional and structural diversification of incomplete phosphotransferase system in cellulose-degrading clostridia
The ISME Journal (2023)
-
Effects of Glucose and Mannose on Nutrient Absorption and Fruit Quality in Peach (Prunus persica L.)
Journal of Soil Science and Plant Nutrition (2023)
