
Abstract
Nitrospira are chemolithoautotrophic nitrite-oxidizing bacteria that catalyze the second step of nitrification in most oxic habitats and are important for excess nitrogen removal from sewage in wastewater treatment plants (WWTPs). To date, little is known about their diversity and ecological niche partitioning within complex communities. In this study, the fine-scale community structure and function of Nitrospira was analyzed in two full-scale WWTPs as model ecosystems. In Nitrospira-specific 16S rRNA clone libraries retrieved from each plant, closely related phylogenetic clusters (16S rRNA identities between clusters ranged from 95.8% to 99.6%) within Nitrospira lineages I and II were found. Newly designed probes for fluorescence in situ hybridization (FISH) allowed the specific detection of several of these clusters, whose coexistence in the WWTPs was shown for prolonged periods of several years. In situ ecophysiological analyses based on FISH, relative abundance and spatial arrangement quantification, as well as microautoradiography revealed functional differences of these Nitrospira clusters regarding the preferred nitrite concentration, the utilization of formate as substrate and the spatial coaggregation with ammonia-oxidizing bacteria as symbiotic partners. Amplicon pyrosequencing of the nxrB gene, which encodes subunit beta of nitrite oxidoreductase of Nitrospira, revealed in one of the WWTPs as many as 121 species-level nxrB operational taxonomic units with highly uneven relative abundances in the amplicon library. These results show a previously unrecognized high diversity of Nitrospira in engineered systems, which is at least partially linked to niche differentiation and may have important implications for process stability.
Similar content being viewed by others

Introduction
Nitrification, the sequential oxidation of ammonia to nitrite to nitrate, is a key nitrogen cycling process in oxic ecosystems and an important step of biological wastewater treatment. Nitrification is a two-step process carried out by chemolithoautotrophic ammonia-oxidizing bacteria and archaea (AOB, AOA) and nitrite-oxidizing bacteria (NOB).
In the environment and in wastewater treatment plants (WWTPs), the most diverse and often predominant known NOB are mainly uncultured members of the genus Nitrospira (Burrell et al., 1998; Juretschko et al., 1998; Daims et al., 2001a; Freitag et al., 2005; Off et al., 2010). Within this genus, six phylogenetic main lineages have been described that occur in a wide range of habitats (Daims et al., 2001a; Lebedeva et al., 2008; Off et al., 2010). In addition, we recently detected an immense diversity of coexisting Nitrospira from these six and several novel lineages in various soils by amplicon pyrosequencing the nxrB gene, which codes for subunit beta of nitrite oxidoreductase and is a useful functional and phylogenetic marker for Nitrospira (Pester et al., 2013).
Most Nitrospira in WWTPs are affiliated to the main lineages I or II (Daims et al., 2001a; Maixner et al., 2006). These two lineages can coexist in the same full-scale WWTPs or lab-scale reactors (Schramm et al., 1998; Maixner et al., 2006; Park and Noguera, 2008) apparently without competitive exclusion and based on ecological niche partitioning. Past research showed that lineage I Nitrospira preferred a higher nitrite concentration than lineage II (Maixner et al., 2006), but lineage II Nitrospira were better adapted to elevated levels of dissolved oxygen (Park and Noguera, 2008). However, it remains unclear whether the main lineages represent the functionally relevant scale of Nitrospira diversity. If these lineages comprise an additional diversity of more closely related clades, which still differ in ecophysiological traits and can coexist by niche partitioning, a higher phylogenetic resolution than the main Nitrospira lineages may be needed to understand the biology of nitrite oxidation.
For other bacterial groups, highly diverse communities of closely related, coexisting members were reported based on marker gene or metagenomic sequence analyses (e.g., Acinas et al., 2004; Cuadros-Orellana et al., 2007; He et al., 2007; Woebken et al., 2008; Denef et al., 2010; Albertsen et al., 2012; Ngugi and Stingl, 2012). However, the functional differentiation of closely related uncultured microbes has not been investigated directly in the source communities by in situ techniques. Here, we tackled this issue by combining phylogenetic analyses of Nitrospira 16S rRNA genes with single-cell in situ tools. This cultivation-independent approach allowed us to investigate the fine-scale community structure and ecophysiology of uncultured Nitrospira, below the phylogenetic level of the main lineages, in two full-scale nitrifying WWTPs. Additionally, the Nitrospira diversity in one WWTP was explored in great depth by amplicon pyrosequencing the nxrB gene.
Materials and methods
Sampling and fixation of activated sludge
Activated sludge was sampled from an aerated, nitrifying activated sludge basin (tank no. 2) of the full-scale WWTP of the University of Veterinary Medicine, Vienna, Austria (‘WWTP Vetmed’) in March, April and July 2004, May and June 2007, November 2008, February and October 2009 and in July and October 2010. This WWTP consists of two continuously operated activated sludge tanks, each with a volume of 254 m3. The influent wastewater composition, including the N-load, varies with the amounts of animal feces and other sewage. The effluent of WWTP Vetmed is subsequently treated by the main municipal WWTP of the city of Vienna. Nitrifying activated sludge samples were also taken from a sequencing batch reactor, which treats reject water from sludge dewatering after anaerobic digestion, at the municipal full-scale WWTP of Ingolstadt (Germany) in October 2007, November 2008, October 2009 and March 2011. The reject water treatment stage of this WWTP consists of three sequencing batch reactors, which together treat up to 450 m3 wastewater per day with an ammonium load of 450 kg NH4+-N per day. Each sequencing batch reactor is intermittently aerated for N-elimination by nitrification and denitrification. All samples were taken from the same sequencing batch reactor.
Activated sludge was collected by centrifugation (11000 r.p.m., 10 min, 4 °C) and frozen at −20 °C until DNA extraction. Additional sludge aliquots were fixed immediately upon arrival in the lab in 3% (v v−1) formaldehyde in 1 × phosphate-buffered saline for 2 h at 4 °C and were stored in 96% ethanol:1 × phosphate-buffered saline (1:1) at −20 °C. For fluorescence in situ hybridization-microautoradiography (FISH-MAR) analyses, sludge was shipped to the laboratory at 4 °C and immediately processed upon arrival.
DNA extraction, clone library construction, Sanger sequencing and phylogenetic analyses
DNA was extracted from activated sludge sampled in February 2009 and October 2010 from WWTP Vetmed and in March 2011 from WWTP Ingolstadt using the Powersoil DNA Isolation Kit (MO BIO Laboratories, Carlsbad, CA, USA) and following the manufacturer’s instructions.
Nitrospira partial 16S rRNA genes were PCR-amplified using primers 8F (Hicks et al., 1992; Juretschko et al., 1998) and Ntspa1158R (Maixner et al., 2006) according to the protocol of Maixner et al. (2006) with the following modifications: initial denaturation for 5 min at 95 °C, primer annealing at 58 °C and final elongation for 10 min at 72 °C. PCR products were cloned by using the TOPO TA cloning system (Invitrogen, Carlsbad, CA, USA), and Sanger sequencing was performed by using the BigDye Terminator Cycle Sequencing Kit v.3.1 and an ABI 3130xl Genetic Analyzer (Applied Biosystems, Foster City, CA, USA) according to the manufacturer’s instructions.
Nitrospira 16S rRNA sequences were phylogenetically analyzed by using the ARB software (Ludwig et al., 2004) and a sequence database containing 345 almost full-length (>1350 nucleotides) reference sequences from the phylum Nitrospirae, which had been retrieved from GenBank. Sequence alignments generated by ARB were manually refined. To check for chimeric sequences, each sequence was separated into two equal parts. The 5′ and 3′ parts were independently added to a Nitrospira reference tree by using the parsimony interactive tool of ARB without changing the overall tree topology, and sequences showing placement of the two parts in different Nitrospira lineages were discarded as probably chimeric. Phylogenetic trees of Nitrospira lineages I and II were calculated using maximum likelihood inference (AxML) (Stamatakis et al., 2002), neighbor joining (ARB implementation, Felsenstein distance correction model) and maximum parsimony within the PHYLIP package (Felsenstein 1989) with a 50% conservation filter for the phylum Nitrospirae that included 1409 unambiguously aligned nucleotide positions.
Probe design, FISH, microscopy and digital image analysis
To design 16S rRNA-targeted oligonucleotide probes specific for single phylogenetic clusters within Nitrospira lineage I or II, we screened the Nitrospira 16S rRNA sequences from the two WWTPs for probe target regions with cluster-specific single-nucleotide polymorphisms. The new probes should distinguish the Nitrospira clusters, which occurred in the same WWTP, from each other rather than from Nitrospira in other habitats or from other bacteria. To compensate for a possible lack of specificity outside the genus Nitrospira, these probes were combined with established specific probes targeting the entire Nitrospira lineages I or II or the whole genus, respectively, which were labeled with a different fluorochrome (Supplementary Table S1). The optimal hybridization and washing conditions for each new probe were inferred from probe dissociation curves with the targeted 16S rRNA molecules as described elsewhere (Daims et al., 1999) by using Clone-FISH (Schramm et al., 2002) and the image analysis software daime (Daims et al., 2006a). The specificity of the new probes within the Nitrospira communities was confirmed in FISH experiments with non-target Nitrospira 16S rRNA (also obtained by Clone-FISH) with one nucleotide mismatch at the probe binding sites. Probes were 5' labeled with the fluorochromes Cy3, Cy5 or FLUOS. Unlabeled competitor probes (Supplementary Table S1) and double-labeled probes (to increase fluorescence intensity) (Stoecker et al., 2010) were used when necessary. All probes were obtained from Thermo Hybaid (Ulm, Germany).
FISH was performed according to Daims et al. (2005) with hybridization times of 2–16 h (Supplementary Table S1). Images of probe-conferred fluorescence were recorded by using a confocal laser scanning microscope (510 Meta; Zeiss, Oberkochen, Germany). The relative abundance (biovolume fraction) of specific Nitrospira clusters (compared with all Nitrospira detected by probe Ntspa662, to all lineage II Nitrospira detected by probe Ntspa1151 or to all lineage I Nitrospira detected by Ntspa1431) was measured by quantitative FISH as described elsewhere (Daims and Wagner, 2007) and by using the daime software. In situ spatial arrangement patterns of Nitrospira and AOB were quantified as described previously (Daims et al., 2006a). Briefly, the pair cross-correlation function was estimated, by image analysis, for a range of μm distances between two FISH probe-defined target groups. For each distance, the pair cross-correlation function indicates the likelihood that the two analyzed groups are within this distance compared with a random arrangement of cells. Further details of FISH and image analysis are provided in Supplementary Text S1.
FISH and microautoradiography
Substrate uptake experiments were performed with living activated sludge from WWTP Ingolstadt sampled in March 2011. To explore the use of organic substrates by Nitrospira, sludge was incubated with one of the substrates [14C]glucose, [14C]formate, [14C]acetate, [14C]pyruvate and [14C]L-valine (Hartmann Analytic, Braunschweig, Germany) in the presence of 1 mM nitrite or 1 mM ammonium or without the addition of any inorganic electron donor. Autotrophic CO2 fixation was studied by incubating sludge with H14CO3− (Hartmann Analytic, Braunschweig, Germany) in the presence of 1 or 0.1 mM nitrite, 1 mM ammonium or without the addition of any inorganic electron donor. Incubations with dead, formaldehyde-fixed sludge were performed to check for adsorption of the labeled substrates to Nitrospira. Further details are provided in Supplementary Text S1. Incubated sludge was fixed in 1.5% (v v−1) formaldehyde in 1 × phosphate-buffered saline overnight at 4 °C. FISH-MAR was performed as described previously (Lee et al., 1999). For each experiment, at least 30 (up to 164) microcolonies of each analyzed Nitrospira phylogenetic cluster were inspected for the presence or absence of a MAR signal and counted. χ2 Tests of significance were performed between incubations involving the same carbon source and Nitrospira cluster by using IBM SPSS Statistics 21.0. We could not exclude the possibility that environmental parameters such as temperature and pH, which were kept constant in all incubations, were suboptimal for some of the studied Nitrospira phylogenetic clusters and thus caused different levels of the metabolic activity of these NOBs. Therefore, the results of the different incubations (fractions of MAR-positive microcolonies) were compared only for the same Nitrospira cluster but not between different clusters.
Long-term incubation
Activated sludge from WWTP Vetmed (sampled in November 2010) was diluted 1:4 in filter-sterilized (0.2 μm) sludge supernatant. Sludge aliquots of 50 ml were incubated in 200 ml glass bottles sealed with aluminum caps with a polytetrafluoroethylene butyl septum (Carl Roth, Karlsruhe, Germany) in the dark at 20 °C and in the presence of 0.05 or 1 mM nitrite. These nitrite concentrations support the growth of Nitrospira isolates and enrichments (Ehrich et al., 1995; Spieck et al., 2006), and in preliminary experiments using these nitrite concentrations and H14CO3−, we observed MAR-positive Nitrospira microcolonies in sludge from WWTP Vetmed (data not shown). All incubations were performed in duplicates. During the 51 days of incubation, the nitrite concentration was frequently checked colorimetrically (Supplementary Text S1) and nitrite was replenished regularly by using peristaltic pumps. To avoid carbon depletion, 10 mM of NaHCO3 was added on days 27 and 47. The pH was measured every 1–2 days and adjusted to 7.6 by adding HCl when necessary. To prevent nitrate accumulation and possible trace element or cofactor depletion, culture supernatant was replaced by filter-sterilized activated sludge supernatant every 7–10 days. Air was pumped into the headspace for 2 min every 4 h by using an aquarium pump (Eheim 106001; Eheim, Deizisau, Germany), and the cultures were continuously shaken at 90 r.p.m. to ensure sufficient aeration. Samples were taken at five time points and were formaldehyde-fixed as described above.
Amplicon pyrosequencing of Nitrospira nxrB
The nxrB genes of Nitrospira were PCR amplified from WWTP Ingolstadt-activated sludge (sampled in November 2008). DNA extraction, PCR with primer set nxrB169f/nxrB638r, 454 amplicon sequencing and bioinformatics analyses were performed as described previously (Pester et al., 2013). Approximate species-level operational taxonomic units (OTUs) were formed by clustering the nxrB reads with 95% nucleotide sequence identity as cutoff (Pester et al., 2013). Representative sequences from selected OTUs were added to an nxrB reference tree (Pester et al., 2013) by using the parsimony interactive tool of ARB or the maximum likelihood-based evolutionary placement algorithm (Berger et al., 2011), respectively, without changing the overall tree topology. The reference tree was based on the tree of Pester et al. (2013) but contained additionally the two paralogous nxrB copies of Nitrospina gracilis (Lücker et al., 2013) as outgroup.
Accession numbers
The 16S rRNA gene sequences obtained in this study have been deposited at GenBank under accession numbers KJ480818–KJ480936. The nxrB sequences were submitted to the Sequence Read Archive (SRA) at GenBank under the accession number SRA047303.
Results
Nitrospira 16S rRNA gene diversity in two WWTPs
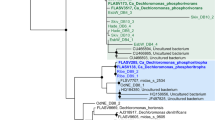
By cloning and Sanger sequencing, partial sequences of Nitrospira 16S rRNA genes (1126–1132 bp) were retrieved from WWTP Vetmed in February 2009 (n=40) and October 2010 (n=22) and from WWTP Ingolstadt in March 2011 (n=58) (Supplementary Table S2). Based on these sequence data, phylogenetic clusters within the main Nitrospira lineages I and II were defined. The criterion used for clustering was that all members of the same cluster must form a monophyletic group in at least one phylogenetic tree (ideally in all trees calculated by different treeing methods). The smallest identified monophyletic groups were defined as separate clusters to maximize the phylogenetic resolution of the grouping. Sequences within the same cluster were highly similar to each other, with identities ranging from 98.6% to 99.9%, but high sequence identities were also observed between different clusters (ranging from 95.8% to 99.6%). In total, 11 clusters were defined, some of which contained only sequences from one of the two studied WWTPs (Figure 1). One lineage I (Ingolstadt clone 43) and one lineage II sequence (Vetmed clone 09-19) did not group with other sequences from this study (Figure 1). Several clusters contained also Nitrospira sequences retrieved in other studies from WWTPs, bioreactors and iron-oxidizing biofilm (Figure 1 and Supplementary Table S2).

Phylogenetic analysis of partial Nitrospira 16S rRNA gene sequences retrieved from WWTPs Vetmed and Ingolstadt. The neighbor joining tree depicts the identified phylogenetic clusters from Nitrospira lineages I and II, with clusters found in WWTP Vetmed colored blue, clusters found in WWTP Ingolstadt colored red and clusters found in both WWTPs colored blue and red, respectively. Clusters also containing Nitrospira sequences from other environments are marked with black in addition to the colors, whereas groups that fully consist of sequences from other sources are drawn completely black. Numbers within parentheses behind cluster names indicate the numbers of sequences in the clusters. The names of FISH probes used to detect the different clusters in situ are indicated together with the name of the respective WWTP and in the matching color. Horizontal gray bars, which link the clusters and probe names, illustrate that each probe was designed for and applied to only one Nitrospira community (in the respective WWTP). Circles on tree branches indicate the support by other treeing methods of the respective monophyletic clusters. Sequences of Nitrospira lineage IV were used as outgroup. The scale bar indicates 1% estimated sequence divergence. The accession numbers of the sequences in each cluster are listed in Supplementary Table S2.
In situ detection of closely related Nitrospira
To confirm that the Nitrospira sequence clusters represented autochthonous community members in the two WWTPs and did not result from transiently present cells, naked DNA or PCR and sequencing errors, FISH experiments were carried out to detect the Nitrospira clusters in situ. For this purpose, new 16S rRNA-targeted oligonucleotide probes were designed based on the sequences in each cluster (Supplementary Table S1). Owing to the high 16S rRNA sequence identities between the different Nitrospira clusters (see above), the design of cluster-specific probes had to rely on SNPs at the probe binding sites to distinguish the clusters. As unique SNPs were rare, we could not design a universally specific probe for each cluster. Instead, the new probes specifically distinguished the clusters found in the same WWTP but might have been unspecific if applied to the respective other WWTP or to samples from other environments. Therefore, each new probe was applied to only one of the WWTPs analyzed in this study (Figure 1) and use of these probes on other samples is not recommended.
In WWTP Vetmed, Nitrospira clusters Id, Ie, IIa, IIb and IIc were specifically detected by FISH (Figures 2a–c), demonstrating that these Nitrospira indeed coexisted in the WWTP. Notably, probe Ntspa1012 targeting cluster Ie has no base mismatch to Vetmed clone 09-2, which is outside cluster Ie (Supplementary Table S2) but could thus not be distinguished from cluster Ie by FISH using this probe. In WWTP Ingolstadt, clusters Ia, Ib and Ig were detected by the respective FISH probes (Figures 2d and e). Cluster IIa was detected in WWTP Ingolstadt by probe Ntspa1151 targeting Nitrospira lineage II (no specific probe was applied because cluster IIa were the only lineage II-related Nitrospira found in this WWTP; Figure 1). The other Nitrospira clusters (Figure 1) were not detected by FISH (for details see Supplementary Text S1). All Nitrospira clusters identified in situ hybridized also to the Nitrospira genus-specific probe Ntspa662 and, except cluster Ig, to the respective group-specific probes targeting Nitrospira lineages I or II (Supplementary Table S1 and Figure 2).

Closely related Nitrospira clusters detected by FISH in WWTPs Vetmed and Ingolstadt. All scale bars depict 10 μm. (a) Clusters IIa (yellow) and IIc (cyan) in WWTP Vetmed (probe Ntspa195 in blue, Ntspa256 in red and Ntspa1151 in green). (b) Clusters IIa and IIc (both cyan) and IIb (yellow) in WWTP Vetmed (probe Ntspa195 and Ntspa256 in blue, Ntspa175 in red and Ntspa1151 in green). (c) Clusters Id (purple) and Ie (yellow) in WWTP Vetmed (probe Ntspa1002 in blue, Ntspa1012 in green and Ntspa1431 in red). (d) Clusters Ia (yellow) and Ib (purple) in WWTP Ingolstadt (probe Ntspa1131 in blue, Ntspa194 in green and Ntspa1431 in red). (e) Cluster Ig (purple) in WWTP Ingolstadt (probe Ntspa451 in red, Ntspa1431 and Ntspa1151 in green, and Ntspa662 in blue). Note that cluster Ig was missed by the lineage I-specific probe Ntspa1431.
All Nitrospira formed spherical or irregularly shaped microcolonies as typically observed for these NOB in activated sludge (Juretschko et al., 1998; Daims et al., 2001a) (Figure 2). As none of these microcolonies hybridized to more than one of the cluster-specific probes in each WWTP (Figure 2), we can largely exclude the possibility that the sequence clusters (Figure 1) might represent multiple 16S rRNA gene copies of very few Nitrospira strains. This result of FISH is in agreement with the presence of only one rrn operon in the genomes of ‘Ca. N. defluvii’ (Lücker et al., 2010) and N. moscoviensis (unpublished data).
Stable coexistence of closely related Nitrospira: an indication of niche partitioning?
FISH using the newly designed high-resolution probes revealed that in each WWTP, the closely related Nitrospira clusters apparently coexisted during prolonged periods for which a sparse set of activated sludge samples was available (∼6 years for WWTP Vetmed and ∼4 years for WWTP Ingolstadt) (data not shown). Interestingly, the co-occurrence of the lineage II Nitrospira clusters in the samples from WWTP Vetmed was accompanied by pronounced shifts in the lineage II Nitrospira community composition (Supplementary Text S1 and Supplementary Figure S1).
The detection of a diverse community of stably coexisting, closely related Nitrospira raised the question of whether these organisms are functionally different and coexist by ecological niche partitioning. To address this question, we took advantage of the unique possibility to detect, distinguish and quantify the closely related Nitrospira in situ and to monitor their substrate utilization by FISH-MAR.
Preference for different nitrite concentrations
We carried out a long-term incubation experiment with activated sludge to test the effects of two ambient nitrite concentrations (0.05 and 1 mM) on different Nitrospira clusters. This experiment was conducted with sludge from WWTP Vetmed, which contained different clusters within both Nitrospira lineages I and II (Figure 1).
Interestingly, quantitative FISH revealed distinct effects of the two nitrite concentrations on the fine-scale community structure within lineage I. Throughout the experiment, the relative abundance of cluster Ie was significantly higher in the incubations with 1 mM nitrite (Figure 3a). The situation was reversed for cluster Id, whose relative abundance was higher in the incubations with 0.05 mM nitrite between days 14 and 31 of the experiment. Subsequently, cluster Id declined in all incubations (Figure 3b).

Relative abundance shifts of Nitrospira lineage I clusters from WWTP Vetmed during incubation with two different nitrite concentrations. All incubations were performed in duplicates (biological replicates 1 and 2), and two technical replicates per time point were analyzed by qFISH. (a) Relative abundance (biovolume fraction) of cluster Ie, compared with total Nitrospira lineage I, at different time points during the incubation. The relative abundance of cluster Ie was significantly higher (Mann–Whitney U-test: P<0.001; t-test: P<0.001) in samples from the high nitrite incubations taken between days 14 and 51. Symbols are as in panel b. (b) Biovolume fraction of cluster Id, relative to total Nitrospira lineage I, at different time points during the incubation. The relative abundance of cluster Id was significantly higher (Mann–Whitney U-test: P<0.001; t-test: P<0.001) in samples from the low nitrite incubations taken between days 14 and 31.
Observation by non-quantitative FISH revealed no obvious effect of the nitrite concentrations on the different lineage II clusters. During all incubations and in the original sludge sampled for this experiment, cluster IIc was predominant and easily detectable by FISH, whereas clusters IIa and IIb were encountered only sporadically in some of the inspected fields of view (data not shown). As the low abundance of clusters IIa and IIb was close to the detection limit of FISH, quantitative FISH would not be precise enough for measuring small abundance shifts of these extremely rare Nitrospira.
Autotrophic and mixotrophic carbon utilization
Another potentially niche-defining property of Nitrospira could be the utilization of simple organic substrates in addition to nitrite and CO2 (mixotrophy) (Watson et al., 1986; Daims et al., 2001a; Lücker et al., 2010). To test whether closely related Nitrospira differed in their mixotrophic potential and substrate spectrum, the uptake by lineage I clusters of radiolabeled formate, acetate, pyruvate, L-valine and glucose was monitored by FISH-MAR. Based on the genomic inventory of ‘Ca. N. defluvii’ (a lineage I member), we hypothesized that these substrates might be used by lineage I Nitrospira (Lücker et al., 2010). These experiments were carried out with sludge from WWTP Ingolstadt as lineage I was more diverse in this WWTP than in WWTP Vetmed (Figure 1). Additional incubations with 14C-labeled bicarbonate instead of organics tested the autotrophic activity of the different lineage I Nitrospira. All incubations were performed in the presence or absence of an inorganic energy source (nitrite or ammonium). The experiments with ammonium were included because Nitrospira might be best adapted to the (unknown) local nitrite concentrations close to nitrite-producing AOB (Maixner et al., 2006).
All in situ detectable Nitrospira lineage I clusters incorporated bicarbonate and formate, but the fractions of MAR-positive microcolonies varied substantially between clusters and conditions. Cluster Ib was most active with the highest fraction of MAR-positive microcolonies (P<0.001) in all experiments. The autotrophic activity of clusters Ia and Ib was not influenced by the incubation conditions, whereas the percentage of MAR-positive cluster Ig microcolonies was significantly higher in the presence of 1 mM nitrite than in the other incubations with 14C-bicarbonate (Figure 4). No Nitrospira assimilated bicarbonate in the absence of nitrite and ammonium (data not shown).

Incorporation of 14C-labeled bicarbonate and formate by Nitrospira lineage I clusters from WWTP Ingolstadt. Fractions of MAR-positive FISH-identified microcolonies (n⩾30 for each cluster) were counted after the different incubation experiments. Significantly different fractions (according to χ2-tests on values obtained for one cluster and carbon source) are indicated by *P<0.05 or ***P<0.001.
Interestingly, formate utilization was more heterogeneous than CO2 fixation. When incubated with 1 mM nitrite, formate incorporation by cluster Ib was significantly lower than with 1 mM ammonium or formate alone (Figure 4). In contrast, the fraction of formate-labeled cluster Ig microcolonies was highest with 1 mM nitrite (Figure 4). Cluster Ia used formate most actively in the incubation amended with ammonium (Figure 4). Finally, only cluster Ib used formate efficiently as the sole substrate (Figure 4 and Supplementary Figure S2).
No Nitrospira lineage I cluster in WWTP Ingolstadt utilized pyruvate, glucose, acetate or valine, whereas uptake of the latter three substrates by other bacteria was observed. The lack of uptake by Nitrospira may be due to genetic differences of the analyzed lineage I clusters compared with ‘Ca. N. defluvii’ and awaits explanation until more Nitrospira genomes become available.
Spatial arrangement patterns
In activated sludge flocs and sessile biofilms, Nitrospira coaggregate with AOB because nitrifiers live in mutualistic symbiosis where ammonia oxidizers produce nitrite, which is consumed and detoxified by NOB (Juretschko et al., 1998; Stein and Arp, 1998; Daims et al., 2001a; Almstrand et al., 2013). Previously, we showed that lineage I Nitrospira occurred closer to AOB microcolonies than members of lineage II, probably because small-scale nitrite concentration gradients formed around AOB, and Nitrospira lineages I and II settled along those gradients according to their different nitrite optima (Maixner et al., 2006). Here we extended our earlier study, which had analyzed Nitrospira only at the level of main lineages, by quantifying the in situ spatial arrangement of coexisting lineage I members relative to AOB and to each other. Different localization patterns might indicate niche-separating features of these closely related Nitrospira. These analyses were carried out with sludge from WWTP Ingolstadt, where the different Nitrospira clusters were more equally abundant and AOB detected by available AOB-targeted FISH probes were much more abundant than in WWTP Vetmed (data not shown). As the AOB-targeted probes (Supplementary Table S1) unexpectedly showed inconsistent hybridization patterns (Supplementary Text S1), only the signal of the AOB-targeted probe Ncom1025 was evaluated.
We detected a strong and highly specific tendency of Nitrospira cluster Ig to coaggregate with the Ncom1025-targeted AOB over a distance range from 9 to 54 μm (Figure 5a). Intriguingly, a completely different pattern was observed for Nitrospira clusters Ia and Ib that were either randomly distributed relative to these AOB (cluster Ia; Figure 5b) or were even negatively correlated with Ncom1025-targeted AOB up to a distance of 42 μm (cluster Ib; Figure 5c). Different localization patterns were also detected by comparing the three Nitrospira lineage I clusters with each other (Supplementary Text S1 and Supplementary Figure S3).

(a–c) Spatial arrangement of Nitrospira lineage I clusters relative to probe Ncom1025-targeted AOB in WWTP Ingolstadt. The pair cross-correlation is plotted as a function of distance between the NOB and AOB. Pair cross-correlation values above one (log2=0, horizontal line) indicate coaggregation, values below one (log2=0) indicate a negative spatial correlation and values not significantly different from one (log2=0) indicate random distribution at the respective distance. Continuous lines depict means calculated from all analyzed images, and error bars show 95% confidence intervals.
The spatial distribution between Nitrospira targeted by probe Ntspa1151 (specific for Nitrospira lineage II; Supplementary Table S1) and the Ncom1025-targeted AOB was also quantified, but no coaggregation was observed (data not shown).
In-depth analysis of Nitrospira richness in WWTP Ingolstadt
The Nitrospira richness in WWTP Ingolstadt was assessed in more depth by amplicon pyrosequencing of nxrB. In total, 9549 Nitrospira nxrB reads passed the quality screening criteria described in Pester et al. (2013). Clustering of these reads using a nucleotide sequence identity threshold of 95%, which is the suggested species threshold for Nitrospira nxrB (Pester et al., 2013), resulted in 121 Nitrospira species-level OTUs that apparently coexisted in the WWTP. As suggested by a high Good’s coverage (Good, 1953) of 0.996, the Nitrospira richness in the amplicon library was almost completely covered by the applied sequencing approach. The ACE and Chao1 nonparametric richness estimators predicted 159 (95% confidence interval 140–195) and 147 (95% confidence interval 132–180) species-level OTUs, respectively. As typical 454 pyrosequencing errors can cause overestimation of microbial diversity (Kunin et al., 2010), our analysis pipeline combined several approaches for bias reduction including sequence quality screening and filtering, frameshift detection, manual sequence curation and chimera exclusion (Pester et al., 2012, 2013). Thus, the 55 OTUs, which were represented by only one or two quality-controlled 454 reads very likely represent rare Nitrospira species, but we cannot exclude the possibility that some of these rare OTUs result from undetected 454 sequencing errors in single reads.
Only six of the 121 species-level OTUs were high in relative abundance (>5%) and together represented approximately 73% of all nxrB sequence reads (Figure 6). As the genomic copy numbers of nxrB range from 2 to 6 in known Nitrospira species (Pester et al., 2013), the quantitative composition of the amplicon library may be influenced by different nxrB copy numbers in Nitrospira genomes from different OTUs. However, this possible bias would not explain the whole range of observed difference in relative abundance and we conclude that the Nitrospira community composition was highly uneven (Figure 6). Phylogenetic analyses were conducted with representative sequences from the species-level OTUs (Supplementary Table S3 and Supplementary Figure S4). In total, 79 (65%) of the OTUs were closely related to Nitrospira lineages I or II that were originally defined based on 16S rRNA gene phylogenies (Daims et al., 2001a) and are also resolved in nxrB-based phylogenetic trees (Pester et al., 2013). Four OTUs of low abundance were tentatively assigned to a new ‘WWTP Ingolstadt 454 lineage’ based on their position in the genus Nitrospira, but outside established lineages in the nxrB reference tree (Supplementary Figure S4 and Supplementary Table S3). The phylogenetic position of 38 OTUs within the genus Nitrospira was ambiguous (Supplementary Table S3), and some of these unassigned OTUs were abundant in the nxrB amplicon library (Figure 6).

Relative abundances of species-level OTUs detected in WWTP Ingolstadt by Nitrospira nxrB amplicon pyrosequencing. Colors indicate the phylogenetic affiliation of the OTUs with the established Nitrospira lineages I and II, and with the new ‘WWTP Ingolstadt 454 lineage’ (see also Supplementary Table S3 for details).
Discussion
In two full-scale WWTPs, application of the full-cycle rRNA approach revealed an unexpectedly high diversity of closely related Nitrospira that coexisted over several years. Moreover, in one of the WWTPs as many as 121 species-level OTUs of Nitrospira were detected by nxrB pyrosequencing, highlighting the enormous amount of apparently coexisting Nitrospira that escape or avoid competitive exclusion in these man-made systems.
Activated sludge offers microhabitats due to the spatial heterogeneity of flocs that lead to strong gradients of electron donors and acceptors (e.g., Schramm et al., 1999; Maixner et al., 2006). This and temporal variations of environmental parameters during the operation of WWTPs (e.g., Daims et al., 2001a) may support the coexistence of NOB strains with different ecophysiological adaptations (niche partitioning). For various microorganisms other than NOB, ecological niche partitioning of closely related strains has been inferred from covariation of the strains with environmental parameters or other organisms (e.g., Coleman and Chisholm, 2007; Fuhrman and Steele, 2008; Hunt et al., 2008; Koeppel et al., 2008; Zo et al., 2008; Connor et al., 2010; Denef et al., 2010; Brown et al., 2012). Niche partitioning of closely related strains was also suggested by differential pigmentation of phototrophs (e.g., Acinas et al., 2009; Haverkamp et al., 2009), by differences in gene content, protein expression and metabolites (Denef et al., 2010; Pena et al., 2010), by in silico evolutionary simulations (Koeppel et al., 2008; Connor et al., 2010; Becraft et al., 2011) and by comparisons of isolates (e.g., Hunt et al., 2008; Acinas et al., 2009; Malmstrom et al., 2010; Pena et al., 2010; Verbeke et al., 2011). All these approaches yielded important insight into functional traits of closely related microorganisms. However, actual phenotypic differences have mainly been demonstrated for cultured isolates. In contrast, ecophysiological studies of specific, closely related, uncultured microorganisms directly in environmental samples have to our knowledge not been undertaken yet. Filling this research gap is important because isolating a collection of closely related strains of slow-growing microbes from a sample is an enormous challenge, and observations made with pure cultures or artificial mixtures of isolates do not reflect the situation in complex communities. Here, the detection of a high diversity of uncultured Nitrospira by FISH with SNP-discriminating, rRNA-targeted probes paved the way for first in situ experiments to elucidate the ecophysiology and coexistence of closely related Nitrospira.
Niche differentiation of closely related Nitrospira
This study provides multiple lines of evidence for ecological niche partitioning of coexisting lineage I Nitrospira in two WWTPs (Figures 3, 4, 5). In a long-term incubation experiment, the Nitrospira from WWTP Vetmed were exposed to different nitrite concentrations and had to compete under these conditions. Their abundance shifts indicated that cluster Ie was more competitive in the presence of a high nitrite concentration than with a low nitrite level, whereas cluster Id showed the opposite trend (Figure 3). The decline of cluster Id in all replicates toward the end of the experiment (Figure 3b) indicated that these microbes were not well adapted to the incubation conditions (e.g., the intensity of aeration that was identical in all incubations) and were finally outcompeted by other lineage I Nitrospira. This suggests additional ecophysiological differences not identified in our study.
Ecological niche partitioning was also shown by FISH-MAR analyses of three Nitrospira lineage I clusters in WWTP Ingolstadt with respect to carbon assimilation (Figure 4). Interestingly, all three Nitrospira clusters utilized formate, although to a different extent (Figure 4). To the best of our knowledge, this study is the first to demonstrate experimentally the use of formate by Nitrospira. This feature of Nitrospira could be advantageous close to anoxic niches in sludge flocs or biofilm, where formate may be released by fermenting microorganisms, or in nitrifying bioreactors treating sludge liquor from anaerobic digesters. Indeed, the source reactor of these Nitrospira at WWTP Ingolstadt treats sludge liquor. As formate is oxidized to CO2 by formate dehydrogenase and CO2 is then fixed (Lücker et al., 2010), its use as a carbon source is not advantageous compared with autotrophy. However, as electron donor with a much lower redox potential, formate should be a better energy source than nitrite. Nevertheless, clusters Ia and Ig apparently required nitrite (added directly to the medium or provided by ammonia oxidation) for carbon assimilation from formate (Figure 4), in contrast to cluster Ib that seemed to be highly efficient in using formate as carbon and also as energy source. The MAR labelling of each Nitrospira cluster was never completely positive or negative. Instead, different fractions of the counted microcolonies from the same Nitrospira cluster were MAR-positive in the various experiments (Figure 4). This heterogeneity may indicate activity differences between microcolonies or further ecophysiological diversification at finer phylogenetic scales than resolved by our FISH approach.
Moreover, the three Nitrospira lineage I clusters in WWTP Ingolstadt showed different localization patterns relative to probe Ncom1025-targeted AOB, indicating niche separation in addition to the differential use of formate. Only cluster Ig coaggregated with these AOB (Figure 5), suggesting that the symbiosis among nitrifiers can be selective at the phylogenetic level studied here. We cannot exclude that additional specific coaggregation existed also between any of the Nitrospira clusters and other AOB, but could not verify this due to the inconsistent hybridization patterns of the AOB-targeted probes 6a192 and NEU (Supplementary Text S1). The coaggregation of cluster Ig with AOB indicates that these Nitrospira may prefer high nitrite levels found in the close vicinity of AOB microcolonies (Maixner et al., 2006). Consistently, FISH-MAR showed a strong increase in the CO2 fixation by cluster Ig in the presence of a high nitrite concentration (1 mM) (Figure 4). However, a specific spatial distribution might also reflect other AOB–NOB interactions such as an exchange of excreted organics or the utilization of siderophores produced by the respective partner (Chain et al., 2003). Similarly, a negative spatial correlation as observed for cluster Ib and the Ncom1025-targeted AOB (Figure 5c) could indicate for example competition for oxygen or limited co-factors required by both groups. Recently, distinct cocorrelation patterns were described for terrestrial Nitrospira and AOA (Pester et al., 2013). Thus, specific interactions among nitrifiers of different phylogenetic lineages could be a widespread and interesting phenomenon that awaits further analysis. Alternatively, the positive and negative spatial correlation of Nitrospira clusters Ig and Ib with Ncom1025-targeted AOB may be caused by preferences for similar or dissimilar environmental conditions within sludge flocs rather than by specific interactions. This would indicate distinct habitat requirements of clusters Ia, Ib and Ig. The lack of coaggregation between AOB and lineage II Nitrospira contrasts previous results obtained for other WWTPs (Maixner et al., 2006) and could be caused by similar effects as discussed for lineage I clusters (see above) or might even indicate that these Nitrospira did not rely on nitrite as energy source.
In summary, our results show that ecological niche differentiation has a role for the coexistence of closely related Nitrospira in WWTPs. The impact of other factors, such as microbial predation (Dolinšek et al., 2013), on Nitrospira community structures remains to be determined.
Function of a diverse Nitrospira community
The continuous presence of several closely related members with different ecophysiology may have interesting implications for Nitrospira community function. The resulting more efficient use of the available niche space might improve the performance of nitrite oxidation, as shown for other functionally redundant groups and processes (Tilman et al., 1997; Loreau et al., 2001; Cardinale, 2011). Furthermore, a diverse Nitrospira community may be advantageous for the stability of nitrite oxidation due to a favorable diversity–stability relationship (Yachi and Loreau, 1999; Daims et al., 2001b; Elmqvist et al., 2003; Tilman et al., 2006; Eisenhauer et al., 2012). With few Nitrospira OTUs being highly abundant at a particular time, a large diversity of less abundant Nitrospira (Figure 6) may act as seed bank for compensatory growth after disturbances such as changes in the wastewater composition in a WWTP or phage attack. As closely related strains can show high genomic plasticity (e.g., Thompson et al., 2005; Cuadros-Orellana et al., 2007; Denef et al., 2010; Albertsen et al., 2012), a high Nitrospira diversity may also represent a large pool of potentially advantageous genes or alleles that could be exchanged by horizontal transfer within the Nitrospira community (e.g., Cuadros-Orellana et al., 2007; Simmons et al., 2008; Cordero et al., 2012) or homologous recombination (Eppley et al., 2007; Denef and Banfield, 2012), respectively.
Implications for future nitrification research
While past studies almost exclusively analyzed Nitrospira in WWTPs at the level of the whole genus or its major lineages (e.g., Schramm et al., 1998; Daims et al., 2001a; Maixner et al., 2006; Park and Noguera, 2008), our results imply that a higher phylogenetic resolution is needed to understand the ecophysiological diversification and abundance dynamics of these organisms. Still, it remains to be determined which units of diversity comprise ecologically cohesive Nitrospira populations. For example, all analyzed Nitrospira clusters contained highly similar but not identical 16S rRNA sequences (Figure 1 and Supplementary Table S2). These sequences might represent Nitrospira with further functional differentiation or just allelic diversity of the 16S rRNA gene among ecophysiologically identical Nitrospira. Moreover, as we analyzed only two relatively highly conserved marker genes coding for 16S rRNA and nxrB, the detected Nitrospira diversity may only be the tip of an iceberg. A next step could be genome-wide comparative analyses of coexisting Nitrospira complemented by physiological experiments to validate genome-based hypotheses on metabolic functions. With single-cell genomics (Blainey, 2013) and methods to study single-cell ecophysiology in situ (Wagner et al., 2006; Wagner, 2009), the required tools would in principle be available. If ammonia oxidizers are also included, nitrifiers in WWTPs can become model organisms for studying functional and evolutionary aspects of bacterial diversity with a high phylogenetic resolution, also in the context of symbiosis, in an easily accessible but complex model ecosystem (Daims et al., 2006b).
References
Acinas SG, Klepac-Ceraj V, Hunt DE, Pharino C, Ceraj I, Distel DL et al. (2004). Fine-scale phylogenetic architecture of a complex bacterial community. Nature 430: 551–554.
Acinas SG, Haverkamp TH, Huisman J, Stal LJ . (2009). Phenotypic and genetic diversification of Pseudanabaena spp. (cyanobacteria). ISME J 3: 31–46.
Albertsen M, Hansen LB, Saunders AM, Nielsen PH, Nielsen KL . (2012). A metagenome of a full-scale microbial community carrying out enhanced biological phosphorus removal. ISME J 6: 1094–1106.
Almstrand R, Daims H, Persson F, Sorensson F, Hermansson M . (2013). New methods for analysis of spatial distribution and coaggregation of microbial populations in complex biofilms. Appl Environ Microbiol 79: 5978–5987.
Becraft ED, Cohan FM, Kuhl M, Jensen SI, Ward DM . (2011). Fine-scale distribution patterns of Synechococcus ecological diversity in microbial mats of Mushroom Spring, Yellowstone National Park. Appl Environ Microbiol 77: 7689–7697.
Berger SA, Krompass D, Stamatakis A . (2011). Performance, accuracy, and Web server for evolutionary placement of short sequence reads under maximum likelihood. Syst Biol 60: 291–302.
Blainey PC . (2013). The future is now: single-cell genomics of bacteria and archaea. FEMS Microbiol Rev 37: 407–427.
Brown MV, Lauro FM, DeMaere MZ, Muir L, Wilkins D, Thomas T et al. (2012). Global biogeography of SAR11 marine bacteria. Mol Syst Biol 8: 595.
Burrell PC, Keller J, Blackall LL . (1998). Microbiology of a nitrite-oxidizing bioreactor. Appl Environ Microbiol 64: 1878–1883.
Cardinale BJ . (2011). Biodiversity improves water quality through niche partitioning. Nature 472: 86–89.
Chain P, Lamerdin J, Larimer F, Regala W, Lao V, Land M et al. (2003). Complete genome sequence of the ammonia-oxidizing bacterium and obligate chemolithoautotroph Nitrosomonas europaea. J Bacteriol 185: 2759–2773.
Coleman ML, Chisholm SW . (2007). Code and context: Prochlorococcus as a model for cross-scale biology. Trends Microbiol 15: 398–407.
Connor N, Sikorski J, Rooney AP, Kopac S, Koeppel AF, Burger A et al. (2010). Ecology of speciation in the genus Bacillus. Appl Environ Microbiol 76: 1349–1358.
Cordero OX, Wildschutte H, Kirkup B, Proehl S, Ngo L, Hussain F et al. (2012). Ecological populations of bacteria act as socially cohesive units of antibiotic production and resistance. Science 337: 1228–1231.
Cuadros-Orellana S, Martin-Cuadrado AB, Legault B, D'Auria G, Zhaxybayeva O, Papke RT et al. (2007). Genomic plasticity in prokaryotes: the case of the square haloarchaeon. ISME J 1: 235–245.
Daims H, Brühl A, Amann R, Schleifer KH, Wagner M . (1999). The domain-specific probe EUB338 is insufficient for the detection of all Bacteria: development and evaluation of a more comprehensive probe set. Syst Appl Microbiol 22: 434–444.
Daims H, Nielsen JL, Nielsen PH, Schleifer KH, Wagner M . (2001a). In situ characterization of Nitrospira-like nitrite-oxidizing bacteria active in wastewater treatment plants. Appl Environ Microbiol 67: 5273–5284.
Daims H, Purkhold U, Bjerrum L, Arnold E, Wilderer PA, Wagner M . (2001b). Nitrification in sequencing biofilm batch reactors: lessons from molecular approaches. Water Sci Technol 43: 9–18.
Daims H, Stoecker K, Wagner M . (2005). Fluorescence in situ hybridization for the detection of prokaryotes. In: Osborn AM, Smith CJ (eds) Advanced Methods in Molecular Microbial Ecology. Bios-Garland: Abingdon, UK, pp 213–239.
Daims H, Lücker S, Wagner M . (2006a). daime, a novel image analysis program for microbial ecology and biofilm research. Environ Microbiol 8: 200–213.
Daims H, Taylor MW, Wagner M . (2006b). Wastewater treatment: a model system for microbial ecology. Trends Biotechnol 24: 483–489.
Daims H, Wagner M . (2007). Quantification of uncultured microorganisms by fluorescence microscopy and digital image analysis. Appl Microbiol Biotechnol 75: 237–248.
Denef VJ, Kalnejais LH, Mueller RS, Wilmes P, Baker BJ, Thomas BC et al. (2010). Proteogenomic basis for ecological divergence of closely related bacteria in natural acidophilic microbial communities. Proc Natl Acad Sci USA 107: 2383–2390.
Denef VJ, Banfield JF . (2012). In situ evolutionary rate measurements show ecological success of recently emerged bacterial hybrids. Science 336: 462–466.
Dolinšek J, Lagkouvardos I, Wanek W, Wagner M, Daims H . (2013). Interactions of nitrifying bacteria and heterotrophs: identification of a Micavibrio-like putative predator of Nitrospira spp. Appl Environ Microbiol 79: 2027–2037.
Ehrich S, Behrens D, Lebedeva E, Ludwig W, Bock E . (1995). A new obligately chemolithoautotrophic, nitrite-oxidizing bacterium, Nitrospira moscoviensis sp. nov. and its phylogenetic relationship. Arch Microbiol 164: 16–23.
Eisenhauer N, Scheu S, Jousset A . (2012). Bacterial diversity stabilizes community productivity. PLoS One 7: e34517.
Elmqvist T, Folke C, Nystrom M, Peterson G, Bengtsson J, Walker B et al. (2003). Response diversity, ecosystem change, and resilience. Front Ecol Environ 1: 488–494.
Eppley JM, Tyson GW, Getz WM, Banfield JF . (2007). Genetic exchange across a species boundary in the archaeal genus ferroplasma. Genetics 177: 407–416.
Felsenstein J . (1989). PHYLIP—Phylogeny Inference Package (Version 3.2). Cladistics 5: 164–166.
Freitag TE, Chang L, Clegg CD, Prosser JI . (2005). Influence of inorganic nitrogen management regime on the diversity of nitrite-oxidizing bacteria in agricultural grassland soils. Appl Environ Microbiol 71: 8323–8334.
Fuhrman JA, Steele JA . (2008). Community structure of marine bacterioplankton: patterns, networks, and relationships to function. Aquat Microb Ecol 53: 69–81.
Good IJ . (1953). The population frequencies of species and the estimation of population parameters. Biometrika 40: 237–264.
Haverkamp TH, Schouten D, Doeleman M, Wollenzien U, Huisman J, Stal LJ . (2009). Colorful microdiversity of Synechococcus strains (picocyanobacteria) isolated from the Baltic Sea. ISME J 3: 397–408.
He S, Gall DL, McMahon KD . (2007). ‘Candidatus Accumulibacter’ population structure in enhanced biological phosphorus removal sludges as revealed by polyphosphate kinase genes. Appl Environ Microbiol 73: 5865–5874.
Hicks RE, Amann RI, Stahl DA . (1992). Dual staining of natural bacterioplankton with 4',6-diamidino-2-phenylindole and fluorescent oligonucleotide probes targeting kingdom-level 16S rRNA sequences. Appl Environ Microbiol 58: 2158–2163.
Hunt DE, David LA, Gevers D, Preheim SP, Alm EJ, Polz MF . (2008). Resource partitioning and sympatric differentiation among closely related bacterioplankton. Science 320: 1081–1085.
Juretschko S, Timmermann G, Schmid M, Schleifer KH, Pommerening-Röser A, Koops HP et al. (1998). Combined molecular and conventional analyses of nitrifying bacterium diversity in activated sludge: Nitrosococcus mobilis and Nitrospira-like bacteria as dominant populations. Appl Environ Microbiol 64: 3042–3051.
Koeppel A, Perry EB, Sikorski J, Krizanc D, Warner A, Ward DM et al. (2008). Identifying the fundamental units of bacterial diversity: a paradigm shift to incorporate ecology into bacterial systematics. Proc Natl Acad Sci USA 105: 2504–2509.
Kunin V, Engelbrektson A, Ochman H, Hugenholtz P . (2010). Wrinkles in the rare biosphere: pyrosequencing errors can lead to artificial inflation of diversity estimates. Environ Microbiol 12: 118–123.
Lebedeva EV, Alawi M, Maixner F, Jozsa PG, Daims H, Spieck E . (2008). Physiological and phylogenetic characterization of a novel lithoautotrophic nitrite-oxidizing bacterium, ‘Candidatus Nitrospira bockiana’. Int J Syst Evol Microbiol 58: 242–250.
Lee N, Nielsen PH, Andreasen KH, Juretschko S, Nielsen JL, Schleifer KH et al. (1999). Combination of fluorescent in situ hybridization and microautoradiography—a new tool for structure–function analyses in microbial ecology. Appl Environ Microbiol 65: 1289–1297.
Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime JP, Hector A et al. (2001). Biodiversity and ecosystem functioning: current knowledge and future challenges. Science 294: 804–808.
Lücker S, Wagner M, Maixner F, Pelletier E, Koch H, Vacherie B et al. (2010). A Nitrospira metagenome illuminates the physiology and evolution of globally important nitrite-oxidizing bacteria. Proc Natl Acad Sci USA 107: 13479–13484.
Lücker S, Nowka B, Rattei T, Spieck E, Daims H . (2013). The genome of Nitrospina gracilis illuminates the metabolism and evolution of the major marine nitrite oxidizer. Front Microbiol 4: 27.
Ludwig W, Strunk O, Westram R, Richter L, Meier H, Yadhukumar et al. (2004). ARB: a software environment for sequence data. Nucleic Acids Res 32: 1363–1371.
Maixner F, Noguera DR, Anneser B, Stoecker K, Wegl G, Wagner M et al. (2006). Nitrite concentration influences the population structure of Nitrospira-like bacteria. Environ Microbiol 8: 1487–1495.
Malmstrom RR, Coe A, Kettler GC, Martiny AC, Frias-Lopez J, Zinser ER et al. (2010). Temporal dynamics of Prochlorococcus ecotypes in the Atlantic and Pacific oceans. ISME J 4: 1252–1264.
Ngugi DK, Stingl U . (2012). Combined analyses of the ITS loci and the corresponding 16S rRNA genes reveal high micro- and macrodiversity of SAR11 populations in the Red Sea. PLoS One 7: e50274.
Off S, Alawi M, Spieck E . (2010). Enrichment and physiological characterization of a novel Nitrospira-like bacterium obtained from a marine sponge. Appl Environ Microbiol 76: 4640–4646.
Park HD, Noguera DR . (2008). Nitrospira community composition in nitrifying reactors operated with two different dissolved oxygen levels. J Microbiol Biotechnol 18: 1470–1474.
Pena A, Teeling H, Huerta-Cepas J, Santos F, Yarza P, Brito-Echeverria J et al. (2010). Fine-scale evolution: genomic, phenotypic and ecological differentiation in two coexisting Salinibacter ruber strains. ISME J 4: 882–895.
Pester M, Rattei T, Flechl S, Grongroft A, Richter A, Overmann J et al. (2012). amoA-based consensus phylogeny of ammonia-oxidizing archaea and deep sequencing of amoA genes from soils of four different geographic regions. Environ Microbiol 14: 525–539.
Pester M, Maixner F, Berry D, Rattei T, Koch H, Lücker S et al. (2013). NxrB encoding the beta subunit of nitrite oxidoreductase as functional and phylogenetic marker for nitrite-oxidizing Nitrospira. Environ Microbiol e-pub ahead of print 10 October 2013 doi:10.1111/1462-2920.12300.
Schramm A, de Beer D, Wagner M, Amann R . (1998). Identification and activities in situ of Nitrosospira and Nitrospira spp. as dominant populations in a nitrifying fluidized bed reactor. Appl Environ Microb 64: 3480–3485.
Schramm A, Santegoeds CM, Nielsen HK, Ploug H, Wagner M, Pribyl M et al. (1999). On the occurrence of anoxic microniches, denitrification, and sulfate reduction in aerated activated sludge. Appl Environ Microbiol 65: 4189–4196.
Schramm A, Fuchs BM, Nielsen JL, Tonolla M, Stahl DA . (2002). Fluorescence in situ hybridization of 16S rRNA gene clones (Clone-FISH) for probe validation and screening of clone libraries. Environ Microbiol 4: 713–720.
Simmons SL, Dibartolo G, Denef VJ, Goltsman DS, Thelen MP, Banfield JF . (2008). Population genomic analysis of strain variation in Leptospirillum group II bacteria involved in acid mine drainage formation. PLoS Biol 6: e177.
Spieck E, Hartwig C, McCormack I, Maixner F, Wagner M, Lipski A et al. (2006). Selective enrichment and molecular characterization of a previously uncultured Nitrospira-like bacterium from activated sludge. Environ Microbiol 8: 405–415.
Stamatakis AP, Ludwig T, Meier H, Wolf MJ . (2002). AxML: a fast program for sequential and parallel phylogenetic tree calculations based on the maximum likelihood method. Proc IEEE Comput Soc Bioinform Conf 1: 21–28.
Stein LY, Arp DJ . (1998). Loss of ammonia monooxygenase activity in Nitrosomonas europaea upon exposure to nitrite. Appl Environ Microbiol 64: 4098–4102.
Stoecker K, Dorninger C, Daims H, Wagner M . (2010). Double labeling of oligonucleotide probes for fluorescence in situ hybridization (DOPE-FISH) improves signal intensity and increases rRNA accessibility. Appl Environ Microbiol 76: 922–926.
Thompson JR, Pacocha S, Pharino C, Klepac-Ceraj V, Hunt DE, Benoit J et al. (2005). Genotypic diversity within a natural coastal bacterioplankton population. Science 307: 1311–1313.
Tilman D, Lehman CL, Thomson KT . (1997). Plant diversity and ecosystem productivity: theoretical considerations. Proc Natl Acad Sci USA 94: 1857–1861.
Tilman D, Reich PB, Knops JM . (2006). Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 441: 629–632.
Verbeke TJ, Dumonceaux TJ, Wushke S, Cicek N, Levin DB, Sparling R . (2011). Isolates of Thermoanaerobacter thermohydrosulfuricus from decaying wood compost display genetic and phenotypic microdiversity. FEMS Microbiol Ecol 78: 473–487.
Wagner M, Nielsen PH, Loy A, Nielsen JL, Daims H . (2006). Linking microbial community structure with function: fluorescence in situ hybridization-microautoradiography and isotope arrays. Curr Opin Biotechnol 17: 83–91.
Wagner M . (2009). Single-cell ecophysiology of microbes as revealed by Raman microspectroscopy or secondary ion mass spectrometry imaging. Annu Rev Microbiol 63: 411–429.
Watson SW, Bock E, Valois FW, Waterbury JB, Schlosser U . (1986). Nitrospira marina gen. nov. sp. nov.: a chemolithotrophic nitrite-oxidizing bacterium. Arch Microbiol 144: 1–7.
Woebken D, Lam P, Kuypers MM, Naqvi SW, Kartal B, Strous M et al. (2008). A microdiversity study of anammox bacteria reveals a novel Candidatus Scalindua phylotype in marine oxygen minimum zones. Environ Microbiol 10: 3106–3119.
Yachi S, Loreau M . (1999). Biodiversity and ecosystem productivity in a fluctuating environment: the insurance hypothesis. Proc Natl Acad Sci USA 96: 1463–1468.
Zo YG, Chokesajjawatee N, Arakawa E, Watanabe H, Huq A, Colwell RR . (2008). Covariability of Vibrio cholerae microdiversity and environmental parameters. Appl Environ Microbiol 74: 2915–2920.
Acknowledgements
We thank Sebastian Lücker for establishing an ARB database of Nitrospira 16S rRNA genes, David Berry and Martin Gruber-Dorninger for help with statistical analyses, Jan Dolinšek, Jeppe Lund Nielsen and Hanna Koch for helpful discussions, and Annika Hennig, Christian Baranyi and Martina Grill for excellent technical assistance. We also thank the operators of the WWTPs Ingolstadt and Vetmed for providing activated sludge samples. This work was supported by the Vienna Science and Technology Fund (WWTF, Grant LS09-40 to HD), an ERC Advanced Grant (Nitricare, Grant 294343, to MW) and the Austrian Science Fund (FWF, Grant P25231-B21 to HD).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on The ISME Journal website
Supplementary information
Rights and permissions
About this article

Cite this article
Gruber-Dorninger, C., Pester, M., Kitzinger, K. et al. Functionally relevant diversity of closely related Nitrospira in activated sludge. ISME J 9, 643–655 (2015). https://doi.org/10.1038/ismej.2014.156
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ismej.2014.156
This article is cited by
-
Succession of bacterial biofilm communities following removal of chloramine from a full-scale drinking water distribution system
npj Clean Water (2023)
-
Wastewater microbial diversity versus molecular analysis at a glance: a mini-review
Brazilian Journal of Microbiology (2023)
-
Differences in distributions, assembly mechanisms, and putative interactions of AOB and NOB at a large spatial scale
Frontiers of Environmental Science & Engineering (2023)
-
Overland flow during a storm event strongly affects stream water chemistry and bacterial community structure
Aquatic Sciences (2022)
-
Nitrospira in wastewater treatment: applications, opportunities and research gaps
Reviews in Environmental Science and Bio/Technology (2022)

{kind=link}