
- NEWS AND VIEWS
Previously unknown human species found in Asia raises questions about early hominin dispersals from Africa
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 568, 176-178 (2019)
doi: https://doi.org/10.1038/d41586-019-01019-7
References
Détroit, F. et al. Nature 568, 181–186 (2019).
Ferring, R. et al. Proc. Natl Acad. Sci. USA 108, 10432–10436 (2011).
Fleagle, J. G. et al. (eds) Out of Africa I: the First Hominin Colonization of Eurasia (Springer, 2010).
Hershkovitz, I. et al. Science 359, 456–459 (2018).
Dennell, R. & Roebroeks, W. Nature 438, 1099–1104 (2005).
Zhu, Z. et al. Nature 559, 608–612 (2018).
Brown, P. et al. Nature 431, 1055–1061 (2004).
Mijares, A. S. et al. J. Hum. Evol. 59, 123–132 (2010).
Kivell, T. L. et al. Nature Commun. 6, 8431 (2015).
Morwood, M. J. et al. Nature 437, 1012–1017 (2005).
van den Bergh, G. D. et al. Nature 534, 245–248 (2016).
Argue, D., Groves, C. P., Lee, M. S. Y. & Jungers, W. L. J. Hum. Evol. 107, 107–133 (2017).
Brumm, A. et al. Nature 441, 624–628 (2006).
Ingicco, T. et al. Nature 557, 233–237 (2018).
Morwood, M. J. & Jungers, W. L. J. Hum. Evol. 57, 640–648 (2009).
 These bones belong to a new species of human
These bones belong to a new species of human
 Read the paper: A new species of Homo from the Late Pleistocene of the Philippines
Read the paper: A new species of Homo from the Late Pleistocene of the Philippines
 Dating of hominin discoveries at Denisova
Dating of hominin discoveries at Denisova
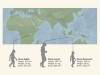 The dawn of Homo floresiensis
The dawn of Homo floresiensis



