- NEWS AND VIEWS
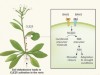
Plant-thickening mechanisms revealed
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 565, 433-435 (2019)
doi: https://doi.org/10.1038/d41586-018-07880-2
References
Lucas, W. J. et al. J. Integr. Plant Biol. 55, 294–388 (2013).
Bar-On, Y. M., Phillips, R. & Milo, R. Proc. Natl Acad. Sci. USA 115, 6506–6511 (2018).
Miyashima, S. et al. Nature 565, 490–494 (2019).
Smetana, O. et al. Nature 565, 485–489 (2019).
Jouannet, V., Brackmann, K. & Greb, T. Curr. Opin. Plant Biol. 23, 54–60 (2015).
Sanio, C. J. Wiss. Bot. 9, 50–128 (1873).
Greb, T. & Lohmann, J. U. Curr. Biol. 26, R816–R821 (2016).
Evert, R. F. Esau’s Plant Anatomy 3rd edn, 323–355 (Wiley, 2006).
Sibout, R., Plantegenet, S. & Hardtke, C. S. Curr. Biol. 18, 458–463 (2008).
Zhang, J., Elo, A. & Helariutta, Y. Curr. Opin. Biotechnol. 22, 293–299 (2011).
 Read the paper: Mobile PEAR transcription factors integrate positional cues to prime cambial growth
Read the paper: Mobile PEAR transcription factors integrate positional cues to prime cambial growth
 Read the paper: High levels of auxin signalling define the stem-cell organizer of the vascular cambium
Read the paper: High levels of auxin signalling define the stem-cell organizer of the vascular cambium
 A cellular passage to the root interior
A cellular passage to the root interior
 Peptide signal alerts plants to drought
Peptide signal alerts plants to drought