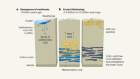
- NEWS AND VIEWS
Proposed early signs of life not set in stone
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 563, 190-191 (2018)
doi: https://doi.org/10.1038/d41586-018-06994-x
References
Nutman, A. P., Bennett, V. C., Friend, C. R. L., Van Kranendonk, M. J. & Chivas, A. R. Nature 537, 535–538 (2016).
Allwood, A. C., Walter, M. R., Kamber, B. S., Marshall, C. P. & Burch, I. W. Nature 441, 714–718 (2006).
Allwood, A. C. et al. Nature 563, 241–244 (2018).
Walter, M. R. (ed.) Stromatolites. Developments in Sedimentology 20 (Elsevier, 1976).
Grotzinger, J. P., Rothman, D. H. Nature 383, 423–425 (1996).
Pope, M. C., Grotzinger, J. P., Schreiber, B. C. J. Sediment. Res. 70, 1139–1151 (2000).
Grotzinger, J. P., Knoll, A. H. Annu. Rev. Earth Planet. Sci. 27, 313–358 (1999).
Bosak, T. et al. Geobiology 10, 384–401 (2012).
Bolhar, R., Kamber, B. S., Moorbath, S., Fedo, C. M., Whitehouse, M. J. Earth Planet. Sci. Lett. 222, 43–60 (2004).
Boak, J. L., Dymek, R. F. Earth Planet. Sci. Lett. 59, 155–176 (1982).
 Read the paper: Reassessing evidence of life in 3,700-million-year-old rocks of Greenland
Read the paper: Reassessing evidence of life in 3,700-million-year-old rocks of Greenland
 Respect for stromatolites
Respect for stromatolites
 Origins of multicellularity
Origins of multicellularity