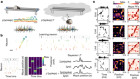
- NEWS AND VIEWS
Models of Parkinson’s disease revisited
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 557, 169-170 (2018)
doi: https://doi.org/10.1038/d41586-018-02589-8
References
DeLong, M. R. Trends Neurosci. 13, 281–285 (1990).
Parker, J. G. et al. Nature 557, 177–182 (2018).
Kravitz, A. V. et al. Nature 466, 622–626 (2010).
Freeze, B. S., Kravitz, A. V., Hammack, N., Berke, J. D. & Kreitzer, A. C. J. Neurosci. 33, 18531-18539 (2013).
DeLong, M. & Wichmann, T. Clin. EEG Neurosci. 41, 61–67 (2010).
Mink, J. W. Progr. Neurobiol. 50, 381-425 (1996).
Nambu, A. Curr. Opin. Neurobiol. 18, 595-604 (2008).
Bocarsly, M. E. et al. Biomed. Opt. Express 6, 4546-4556 (2015).
Klaus, A. et al. Neuron 95, 1171–1180 (2017).
Barbera, G. et al. Neuron 92, 202-213 (2016).
Tecuapetla, F., Matias, S., Dugue, G. P., Mainen, Z. F. & Costa, R. M. Nature Commun. 5, 4315 (2014).
Tecuapetla, F., Jin, X., Lima, S. Q. & Costa, R. M. Cell 166, 703-715 (2016).
Cui, G. et al. Nature 494, 238-242 (2013).
Hikosaka, O., Kim, H. F., Yasuda, M. & Yamamoto, S. Annu. Rev. Neurosci. 37, 289-306 (2014).
Turner, R. S. & Desmurget, M. Curr. Opin. Neurobiol. 20, 704-716 (2010).
McIver, E., Atherton, J., Surmeier, D. & Bevan, M. D. In Neuroscience 2015 Abstr. 708.711 (Soc. Neurosci., 2015).
Wang, J. et al. Brain Stimul. 9, 609-617 (2016).
Adamchic, I. et al. Mov. Disord. 29, 1679-1684 (2014).
Villalba, R. M. & Smith, Y. J. Neural Transm. 125, 431–447 (2017).
 Read the paper: Diametric neural ensemble dynamics in parkinsonian and dyskinetic states
Read the paper: Diametric neural ensemble dynamics in parkinsonian and dyskinetic states
 Vivid views of the PINK1 protein
Vivid views of the PINK1 protein
 Neural synchronization in Alzheimer’s disease
Neural synchronization in Alzheimer’s disease