
Abstract
The inhibitory effect of parvovirus H-1 on the colony-forming ability in vitro of QGY -7703, a cultured human hepatoma cell line, and on the formation and growth of its tumors in nude mice was studied. With higher multiplicity of infection (MOI) of H-1 given, survival of the QGY -7703 cells was found to be decreased. H-1 DNA amplification level at 30 h postinfection (p.i.) was detected to be 7.4 times higher than that at 2 h by dispersed cells assay, while the cells were delayed to enter into S phase. Plaques were formed in the indicator cells (new-born human kidney cell line, NBK) by progeny H-1 virus particles released from the infected QGY -7703 cells by infectious cell center assay. The formation of tumors in nude mice by QGY -7703 cells which were injected s c at 2 h postinfection was observed to be prevented in 2 groups with given MOI 25 and 50. The tumor growth of MOI 10 group occurred at a lower exponential rate than that of control, after a 20 d latent period. It was evident that parvovirus H-1 exhibited a direct inhibitory effect on the formation and growth of human hepatoma cells in vivo as well as in vitro.
Similar content being viewed by others

Introduction
The non-enveloped virion of parvoviruses has a diameter of 20-26 nm and contains a linear, single strand DNA genome of approximately 5 kilobases encapsidated with a simple icosahedral protein coat. Parvoviruses are divided into 2 groups: (1) Adeno-associated viruses, which need a coinfection with a helper virus, such as adenovirus or herpesvirus for a productive infection of host cells; (2) Autonomous parvoviruses1, which require no coinfection and rely on proliferation and differential states of host cells for a productive life cycle2. Autonomus parvoviral DNA does not integrate into its host genome3. H-1, an osteolytic virus of hamsters, belongs to the autonomous parvoviruses4.
Many papers have ldentified an oncosuppressive effect of parvoviruses. The viruses lowered the incidences of some spontaneous and induced tumors5, 6. Parvoviruses were able to kill many kinds of transformed cells. Minute virus of mice (MVM) infection decreased markedly the viability of simian virus 40 (SV40) transformed MVM-resistant BALB/3T3 variant as measured by cell colony-efficiency on plastic plates7. H-1 virus suppressed the growth of human fibroblast transformed by SV40 8 and human keratinocytes transformed by Ha-ras oncogene9. Our previous studies have documented that H-1 virus displayed a selective antineoplastic activity on human gastric cancer or hepatoma tissues cultured directly after surgical operation, while the killing activity on their juxta-cancerous tissues could not be observed10, 11. There was a report about the suppression of H-1 to the formation and growth of tumors in nude mice by transformed mammary epithelial cells12.
The mechanism of the preferentially oncosuppressive activity of parvovirus has not yet been completely elucidated. However, at earlier stages of viral life cycle there were no detectable differences whether in normal cells or in their transformed counterparts, while at later stages, significant differences did appear. In most transformed cells, amplification of replicating form DNA (RF DNA) maintained at a much higher level than that in their normal parent cells. Thus, RF DNA amplification level seemed to be a good mark to ascertain whether host cells have transformed or not8. It has been documented that the nonstructural protein type 1 (NS-1) of MVM displayed a cytotoxic activity13, and that NS-1 and NS-2 might act synergistically in the cytopathogenicity of parvovirus MVM14. The amount of NS-1 accumulated in transformed cells was several times higher than that in the normal. This might be a useful index to imply that the tumors or the transformed cells were inhibited15, 16.
Our previous studies have reported the preferentially cytolytic effect by parvovirus H-1 in 4 kinds of cultured hepatoma cell lines10. To elucidate the mechanisms, we assessed quantitatively the killing effect, H-1 DNA amplification and host cell DNA synthesis, and investigated this effect on the heterotransplanted human hepatoma in nude mice.
Materials and Methods
1. Cell line and virus
Human hepatoma cell line QGY -770317 was cultured in RPMI 1640 medium supplemented with 10% fetal calf serum (FCS), in 5% CO2 and saturated vapor at 37 °C. Tumorigenic characteristics of the cells was determined with high colony-forming efficiency (46%) in 0.3% soft agar and 100% tumorigenicity in nude mice after s c injection or transplantation.
H-1 virus was propagated in NB-K cells (a SV40-transformed new-born human kidney cell line) and purified according to the published method18. Virus titration by plaque assay was mearsured as described by Ledinko19. Virus infection was performed as the published method20, and determined with multiplicity of infection (MOI), the ratio of plaque-forming unit (PFU) to the amount of infected cells.
2. Plasmid
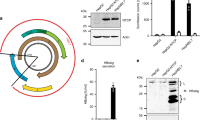
Plasmid pULB 3514 (a gift of Prof J Rommelaere, Universite Libre de Bruxelles) was constructed with BamHI fragment of pUC12 and Bg1II fragment of H-1 DNA. The construct of the plasmid was outlined in Fig 1. Amplification, purification and identification were performed as described in “Molecular Cloning”21.

Construct of plasmid pULB3514
3. Colony-forming efficiency in soft agar
For measurement of anchorage-independent growth, 500 QGY -7703 cells in 4 ml of RPMI 1640 medium containing 10% FCS and 0.3% agar were seeded onto each 6 cm diameter plastic dish (Nunc) overlayed previously with 4 ml of RPMI 1640 medium supplemented with 10% FCS and 0.6% agar. Colonies were counted 6-7 d later by microscopic examination.
4. Survival of the cells after infection
The 500 QGY -7703 cells infected with H-1 virus of mock infected in 4 ml of RPMI 1640 medium were plated onto each 6 am-diameter Nunc dish. After 6-7 days culture, cells were fixed with Carnoy solution and stained with Giemsa and colonies were counted under light microscope. Colony-forming efficiency of the infected cells in RPMI 1640 medium containing 0.3% agar was also examined according to Method 3.
The cell survival rate was calculated as follows:

5. H-1 virus DNA amplification and host cellular DNA synthesis
Total viral DNA within host cells was measured by dispersed cell assay22. Plasmid pULB 3514 was labled with α-32 P -d CTP by nick translation. Labeled plasmid was separated from unincorporated precursors by chromatography with Sephadex G-50 column. An amount of 1 × 106 cells infected with H-1 virus or the mock infected cells was dispersed onto nitrocellulose membrane (Schleider and Schuell BA85, 0.45 μm, ф 25 mm) and hybridized with labeled plasmid. Radioactivity was measured with a liquid scintillator. In the determination of viral DNA synthesis, the background radioactivity inherent in the hybridization method and the input radioactivity by uptake of viral DNA were measured with samples uninfected and harvested at 2 h postinfection, respectively; while the peak of viral DNA synthesis was shown at 30 h postinfection 8, when viral DNA had completed the conversion from double-strand monomers to multimers23. Viral DNA amplification level was obtained from the ratio of intracellular H-1 DNA contents at 30 h vs 2 h postinfection.

For measurement of the effect of H-1 virus on cell cycle, especially on the percentage of S-phase cells, QGY -7703 cells infected with H-1 virus at MOI 5 were harvested at various time intervals and fixed immediately with 70% ethanol solution. Then, the fixed cells were treated with 1% Triton X -100 solution , digested by RNase, stained with propidium iodide solution, finally filtered with 200-net nylon membrane. After the above procedures, the percentages of difierent cell phase were determined with an EPICS -751 fluorescence analyzer24.
6. Progeny virus in cells
Progeny virus was detected by infectious cell center assay. After 5 h incubation p.i, cells were washed with PBS 3 times, then plated onto the indicating NB-K cells, the most susceptible cells to H-1 infection. After 1 wk culturing, plaques which indicated cells releasing infectious progeny virions were formed and counted.
7. Inhibitory effect of H-1 on the formation and growth of tumors by QGY-7703 cells in nude mice
Cells were infected with H-1 virus at various MOIs, 10, 25, 50 for 1 h, then incubated with RPMI 1640 medium plus 10% FCS for one more hour. The infected cells were trypsinized and suspended in RPMI 1640 medium, then 1×107 cells were injected s c at the flanked fat pad of BALB / C nude mice aged 4-6 wk old. Cell viability was determined with trypan blue under the same condition as in in vitro assay. Three dimesions of tumors (L, W, H) formed in nude mice were measured twice weekly after injection. The volume of tumors was caculated as follow:
V= 4/3 π × (L/2 × W/2 × H/2) 25
Results
A. Effects on QGY -7703 cells by H-1 virus
1. The survival rate of QGY -7703 cells decreased due to H-1 infection
The survival rate became lower at a higher MOI given, either in RPMI 1640 medium (Fig 2A) or in RPMI 1640 medium containing 0.3% agar (Fig 2B), both showing the relation between the amount of virus used and growth inhibition of QGY -7703 cells.

H-1 effect on survival of QGY -7703 cells. Survival of QGY -7703 cells infected (• - •) with different MOI 1,5,10 of H-1 and mock infected (○) by colony-formed ability assay (A), and anchorage-independence assay (B). Average valued from 3 experiments, bars represented SEM.
2. H-1 DNA amplified in QGY -7703 cells and cellular DNA synthesis decreased
Three kinds of cell samples were collected, including 2 postinfection samples, i e, at 2 h to determine the uptake of viral DNA, and 30 h to indicate the peak of viral DNA amplification and the uninfected control sample. In infected cells, H-1 DNA amplification at 30 h postinfection was 7.4 times higher than that at 2 h (Tab 1). These results showed that QGY -7703 cells supported the viral DNA amplification. Meanwhile, H-1 infection greatly reduced the percentage of S-phses cells in the cell cycle (Tab 2) and cells were hindered in G1 or Go phase, indicating that cellular DNA synthesis was inhibited. The reduction of S-phase cells maintained from 2 h p.i. to 30 h p.i. Apparently, parvoviral DNA replication exhausted DNA replication enzyme system of host cells on a large scale of amplification, and cellular DNA synthesis itself was thus disordered and inhibited.
3. Percentage of cells releasing infectious progeny virions increased at higher MOI
Infection progeny virions released from QGY -7703 cell formed a plaque on the NB-K monolayer, that stood for an infectious cell center. At a higher MOI given, the percentage of cells releasing progeny virions increased (Tab 3). These results showed that higher MOI infection improved the lytic efficacy to QGY -7703 cells.
B. Effect of H-1 on the formation and growth of tumors by QGY -7703 cells in nude mice
1. Cell viability in treated and control groups remained at a high level within 2 h postinfection
The QGY -7703 cells were injected into nude mice just 2 h p.i.. The viability of the cells at this time obtained from trypan blue staining did not show any obvious differences among the control and 3 treated groups (Tab 4). Hence, any difference of tumor formation and growth in nude mice among 4 groups could not be due to the culturing and viral infecting steps in vitro.
2. H-1 virus inhibited the formation and growth of hepatoma by QGY-7703 cells in nude mice
Tumors grew much more rapidly in control group than those in the 3 treated groups (Fig 3). In the control group, tumors expanded at such an exponential rate that the overlying skin became denuded with tumors burgeoning on d 31 after trasplantation. Thus, mice in the control group were killed on that day, with an average tumor weight of 2.3 g. In the MOI 10 group, tumors remained non-palpable throughout the first 20 d, then regrew noticeably at a much lower exponential rate than those in the control group (Fig 3). Tumors did not appear at all in other 2 treated groups, MOI 25 and MOI 50. These results suggested that H-1 virus harbored an obvious oncosuppression in vivo as well as in vitro.

Tumor growth curve of QGY-7703 cells in nude mice.
1 × 107 of QGY-7703 cells, mock-infected (• -•) and infected with H-1 at MOI 10 (○ - ○) , were injected s c at flanked fat pad of nude mice. The size of tumors was measured twice weekly after transplantation. Each point averaged from 4 values, bars represented SEM.
Higher MOIs of H-1 virus exhibited a more significant inhibitory effect on tumorigenicity as well as on growth. Within the 3 treated groups, MOI 25 or 50 tumor fromation was completely suppressed and no growth occurred in the 56 d period of observation after transplantation. It might be concluded that oncosuppressive effect was greatly affected by virus uptake strategy, i e, the MOI chosen, or the total amount of virions for infection.
Discussion
As far as parvoviral inhibition of tumor growth was concerned, there are 3 possibilities. First, suppression occurred during cell transformation. For instance, adenoassociated virus inhibited the transformation of adenovirus or herpes virus26. Second, an indirect effect by triggering the host immune surveillance system against tumors, including production of immunogenic viral proteins or cytotoxic factors, and inducement of interference factors such as the interferon and tumor necrosis factor27, and /or stimulation of new antigens on cell surface so that they could easily be recognized by the host immune system 28. Finally, a direct cytolytic effect. Our previous studies10, 11 showed that H-1 acts cytolytically on various human cancer cells of different tissues (or origins), but has no obvious inhibitory effect on normal cells of the same origin. Moreover, this viral killing effect on malignant cells is only selective. Recently, Lin et al discovered that the human fetal liver cell line, HuL-1, which had spontaneously transformed to (pre)-neoplastic cells through several scores of passages in our lab, became much more sensitive to H-l infection, thus offered an evidence for selective killing of H-1 (data to be published). In this study, a suppressive effect of H-1 virus on tumorigenicity and growth of hepatoma by QGY -7703 cells in nude mice, as well as inhibition to colony formation in vitro have been demonstrated. Parvovirus H-1 exhibited direct killing effect on hapatoma cells whether in vivo or in vitro. This conclusion was supported by another report, which suggested that the recognition of parvovirus-specific antigens on the surface of infected tumor cells as a target for an increased immune rejection almost certainly does not constitute a major component of parvoviral oncosuppression29.
The mechanism of oncosuppression of H-1 virus indeed needs further study. However, it would become clearer at the DNA replication level. On the one hand, viral DNA amplification was supported by host cells, as was shown above. On the other hand, cellular DNA synthesis deregulated to a much lower level several hours after infection. H-1 virus seemed to interfere with the cellular DNA replication by exhausting the replication enzyme system for its own DNA amplification. This might be one aspect of the so-called direct killing effect. With regard to the viral gene expression in hepatoma cells, recent studies showed that NS-1 protein was accumulated to a higher level in hepatoma cells than that in other non-permissive cells (Lin et al., unpublished data). It seemed that NS-1 protein was cytotoxic, though the metablic mechanisms of this protein on tumor cells remained to be explored.
Analyzing the results from nude mouse experiment, it was interesting to note that the exponential growth rate in the MOI 10 group was much lower than that in the control group. What is its mechanism? We might suppose that cells uninfected or infected beyond S phase may regrow after a latent period; that the progeny virions may attack the hepatoma cells successively; or through a combined effect of both. Further studies on the distribution of virus in tumors and main tissues might be more fruitful.
H-1 virus did not show any potent ability to act as a carcinogen5, 30. Studies on the oncosuppression of H-1 virus might lead to some new approaches in human hepatoma prevention and therapy.
References
Tartersall P, Ward DC, eds. Replication of mammalian parvoviruses. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York 1978:3–12.
Cotmore SF, Tattersall P . The autonomously replicating parvoviruses of vertebrates. Adv Virus Res 1987; 33:91–169.
Hirt B, Parvoviruses and Cancer. In: Celie J, Graessmann A, eds. Cell Transformation. NATO ASI Series, Sery A: Life Sciences 1985; Plenum Press, New York. 94:175–83.
Hauswirth WW, In: The Parvoviruses. Plenum Press, New York and London 1984; 129–52.
Toolan HW . Lack of oncogenic effect of the H-1 viruses for hamster. Nature 1967; 214:1036.
Toolan HW . Inhibition of 7, 12-dimethylbenzanthracene induced tumors in Syrian hamster by prior infection with parvovirus H-1. Cancer Res 1982; 42:2552–5.
Mousset S, Rommelaere J . Minute virus of mice inhibits cell transformation by Simian virus 40. Nature 1982; 300:537–9.
Chen YQ, de Foresta F, Hertoghs J, Avalosse B, Cornelis J, Rommelaere J . Selected killing of Simian virus 40-transformed human fibroblasts by parvovirus H-1. Cancer Res 1986; 46:3574–9.
Chen YQ, Taynder MC, Cornelis J, Boupcamp P, Fusenig NE, Rommelaere J . Sensitization of human keratinocytes to killing by parvovirus H-1 takes palce during their malignant transformation but does not require them to be tumorigenic. Carcinogenesis 1989; 10:163–7.
Luo ZY, Su ZZ, Guo LP, Li JZ, Liu YL . The inhibitory effect of parvovirus H-1 on cultured human cancer cell or transformed cells. Cerutti PA, eds. Anticarcinogenesis and Radiation Protection. Plemun Press, New York 1988:451–5.
Lin WM, Cui YZ, Yu BG, Luo ZY . Preliminary comparison of sensitivity toward parvovirus H-1 of human hepatoma cells and parahepatoma tissue. Chin J Cancer Res. 1989; 1:15–20.
Dupressoir T, Vanacker JM, Cornelis J, Duponchel N, Rommelaere J . Inhibition by parvovirus H-1 of the formation of tumors in nude mice and colonies in vitro by transformed human mammary epithelial cells. Cancer Res 1989; 49:3203–8.
Caillet-Fauquet P, Perros M, Brandenburger A, Spegelaere P, Rommelaere J . Programmed killing of human cell by means of an inducible clone of parvovirus gene encoding NS protein. EMBO J 1990; 9:2989–96.
Brandenburger A, Legendre D, Avalosse B, Rommelaere J . NS-1 and NS-2 proteins may act synergistically in the cyteopathogenicity of parvovirus MVMp. Virology 1990; 174:576–84.
Cornelis JJ, Chen YQ, Spruyt N, et al. Susceptiblity of human cells to killing by the parvovirus H-1 and minute virus of mice correlates with viral gene transcription. J Virol 1990; 64:2537–44.
Avalosse BA, Berrijal S, Chen YQ, Cassiman JJ, Rommelaere J . Identification of a transformation sensitive nuclear protein from normal fibroblasts that specifically interacts with minute virus of mice DNA and correlates with cell resistaace to the parvovirus. Mol Carcinog 1989; 2:245–52.
Wang JB, Zhu DH, Ye XZ, Chen MR . Establishment and some characteristics of a human hepatoma cell line (QGY -7703). Chin J Oncol 1981; 3:241–4.
Tattesall P, Cawte PJ, Shatkin AJ, Ward DC . Three structural polypeptides coded for by minute virus of mice, a parvovirus. J Virol 1976; 20:273–89.
Ledinko N, Toolan HW . Plaque assay of the effect of cytosine arabinoside and 5-iodo-2′ deoxyuridine on the synthesis of H-1 virus particles. Nature 1967; 214:1346–7.
Guo LP, Su ZZ, Luo ZY, Chen TS, Lan Y, Pu XH . Malignant transformation increase cell permissiveness to parvovirus H-1. Chin J Cell Biol 1988; 3:125–9.
Sambrook J, Fritsch EF, Maniatis T, eds. Molecular Cloning, a laboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York 1989; 1.21–1.52.
Lavi S . Carcinogen-mediated amplification of viral DNA sequences in Simian virus 40-transformed Chinese hamster embryo cells. PNAS (USA) 1981; 78:6144–8.
Tatterall P, Ward DC . Rolling hairpin model for replication of parvovirus and linar chromosomal DNA. Nature 1976; 263:106–9.
Crissman HA, Stevenson AP, Kissane RJ, Tobey RA . Techniques for quantitative staining of cellular DNA for flow cytometric analysis. In: Melamed MR, Mullaney PF and Mendelson ML eds. Flow cytometry and sorting. John Willey Inc. New York 1979; 243–61.
Looney WB, Longerbeam HA, Hopkins HA, Carter WH . Solid tumor models for the assessment of different treatment modelities. Cancer (Philadel.) 1983; 51:1012–20.
Ostrove JM, Duckworth DH, Berns KI . Inhibition of adenovirus-transformed cell oncogenicity by adeno-associated virus. Virology 1981;113:521–33.
Rood PA, Lorence RM, Kelley KW . Serum protease inhibitor abrogation of Newcastle disease virus enhancement of cytolysis by recombinant tumor necrosis factors alpha and beta. J Nat Cancer Inst 1990; 82:213–7.
Kelloff G . An approach to C-type virus immunoprevention of spontaneously occurring tumor in laboratory mice. Cancer Res 1976; 36:622–30.
Rommelaere J, Tattersall P . Oncosuppression by parvovirus. In: Tijssen P. ed. Handbook of Parvoviruses. CRC Press, Boca Raton. 1990; 12:21–57.
Luo ZY, Chen TS, Yan SJ, et al. Protein sequence homology comparison of viral proteins and oncoproteins as evidence for the lack of oncogenic potency in parvoviruses. J Fudan Univ (Natural Science) 1991;4:406–12.
Acknowledgements
The authors thank Prof J Rommelaere for providing plasmids. Dr SY Zhang for establishing human hepatoma model in nude mice, Ms ZB Qian for technical help in nude mice experiments, Drs YH Xu, ZZ Su and WM Lin for critically reading the manuscript. This study was supported by National Natural Sciences Foundation of China, National Commission on Education of China, Science and Technology Commission of Shanghai and Scientific Funds of Fudan University.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Yan, S., Ma, C., Chen, X. et al. Inhibitory effect of parvovirus H-1 on the formation of colonies of human hepatoma cell line in vitro and its tumors in nude mice. Cell Res 4, 47–56 (1994). https://doi.org/10.1038/cr.1994.5
Received:
Revised:
Accepted:
Issue Date:
DOI: https://doi.org/10.1038/cr.1994.5
Keywords
This article is cited by
-
Research and development of cancer-targeting vectors: An exploration on oncotropism and oncosuppression of autonomous parvoviruses
Chinese Journal of Cancer Research (2000)
