
Abstract
Altered expression of the multifunctional protein WRAP53β (WD40 encoding RNA Antisense to p53), which targets repair factors to DNA double-strand breaks and factors involved in telomere elongation to Cajal bodies, is linked to carcinogenesis. While loss of WRAP53β function has been shown to disrupt processes regulated by this protein, the consequences of its overexpression remain unclear. Here we demonstrate that overexpression of WRAP53β disrupts the formation of and impairs the localization of coilin to Cajal bodies. At the same time, the function of this protein in the repair of DNA double-strand breaks is enhanced. Following irradiation, cells overexpressing WRAP53β exhibit more rapid clearance of phospho-histone H2AX (γH2AX), and more efficient homologous recombination and non-homologous end-joining, in association with fewer DNA breaks. Moreover, in these cells the ubiquitylation of damaged chromatin, which is known to facilitate the recruitment of repair factors and subsequent repair, is elevated. Knockdown of the ubiquitin ligase involved, ring-finger protein 8 (RNF8), which is recruited to DNA breaks by WRAP53β, attenuated this effect, suggesting that overexpression of WRAP53β leads to more rapid repair, as well as improved cell survival, by enhancing RNF8-mediated ubiquitylation at DNA breaks. Our present findings indicate that WRAP53β and RNF8 are rate-limiting factors in the repair of DNA double-strand breaks and raise the possibility that upregulation of WRAP53β may contribute to genomic stability in and survival of cancer cells.
Similar content being viewed by others

Main
We previously identified the RNA produced from the WRAP53 (WD40 encoding RNA Antisense to p53) gene as an antisense transcript (WRAP53α) that stabilizes the tumor suppressor p53.1 In addition, this gene encodes the WRAP53β protein (also referred to as WRAP53 or WDR79 or TCAB1), which does not regulate p53 but instead is involved in the regulation of telomere elongation and repair of DNA double-strand breaks by recruiting telomerase to nuclear Cajal bodies and the repair factor RNF8 to these break, respectively.2, 3 The role played by WRAP53β in the repair of DNA double-strand breaks is independent of p53, as WRAP53β regulates DNA repair also in cells that lack p53 expression.3, 4 WRAP53β also directs coilin, the survival of motor neuron (SMN) protein and small Cajal body-associated (sca) RNAs to Cajal bodies.2, 5, 6
Several lines of evidence indicate that WRAP53β itself also acts as a tumor suppressor. For example, mutations that attenuate its nuclear localization and telomere function cause dyskeratosis congenita, which enhances the risk for developing cancer.7, 8 These mutations also prevent binding to the DNA repair factor γH2AX required for the accumulation of WRAP53β at DNA breaks,9 indicating that disturbed DNA repair may contribute to the pathogenesis of dyskeratosis congenita. Furthermore, loss of nuclear WRAP53β or single-nucleotide polymorphisms in the WRAP53 gene is correlated with shorter survival of patients with head and neck, breast and ovarian cancer.4, 10, 11, 12, 13, 14, 15 In addition, attenuated expression of this protein correlates with disruption of the DNA damage response in ovarian tumors,4 as well as with resistance of head and neck cancer to radiotherapy.14 Accordingly, altered DNA repair may be the underlying cause of cancers associated with abnormalities in WRAP53β and influence the response of such tumors to treatment.
At the same time, overexpression of WRAP53β is observed in a variety of cancer cell lines compared with non-transformed cells.16 WRAP53β is also overexpressed in primary nasopharyngeal carcinoma,17 esophageal squamous cell carcinoma,18 non-small-cell lung cancer19 and rectal cancer20 and knockdown of this protein in cancer cells subsequently grafted into mice reduces the size of the tumors formed.17, 19 In esophageal squamous cell carcinoma, overexpression of WRAP53β was significantly correlated with tumor infiltration depth, clinical stage and lymph node metastasis.18 However, for none of the studies mentioned above significant associations between WRAP53β overexpression and patient survival were demonstrated. Therefore, although WRAP53β is clearly overexpressed in some tumor types, the clinical relevance of such overexpression remains unclear.
Thus, while loss of WRAP53β function impairs DNA repair and telomere maintenance, which enhances genomic instability and carcinogenesis, the role of WRAP53β overexpression in connection with carcinogenesis is poorly understood. Here, we examined the potential influence of such overexpression on the DNA damage response.
Results
Overexpression of WRAP53β disrupts Cajal bodies and the overexpressed protein is mainly soluble
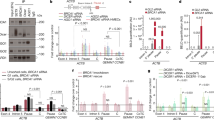
The WRAP53β protein is highly enriched in Cajal bodies, and to examine whether this localization is altered upon overexpression, the total protein lysate from human U-2 osteosarcoma (U2OS) cancer cells that stably overexpress Flag-tagged WRAP53β was analyzed with both rabbit α-WRAP53-C2 antibody, which detects both the nuclear and cytoplasmic forms, and the mouse monoclonal α-WDR79 clone 1F12, which detects the nuclear variant, and is the only antibody that can be used to visualize this protein in repair foci.3, 4 Examination of western blots with either of these antibodies revealed elevated expression of WRAP53β (Figure 1a).

Overexpressed WRAP53β disrupts Cajal bodies and the overexpressed protein is mainly soluble. (a) Western blotting of the levels of WRAP53β in Mock (endogenous WRAP53β) or 1xFlag-WRAP53β (overexpressing WRAP53β) U2OS cells with WRAP53-C2 or WRAP53-1F12 antibodies. β-Actin was used as a loading control. Asterisk indicates unspecific bands. (b) Mock or Flag-WRAP53β U2OS cells were immunostained for WRAP53β (with WRAP53-C2 or WRAP53-1F12 antibodies) and coilin (a marker for Cajal bodies). In all immunofluorescent stainings, nuclei were stained with DAPI. (c) The soluble proteins in Mock and Flag-WRAP53β U2OS cells were removed by extraction before fixation with paraformaldehyde and immunostained for WRAP53β and coilin. (d) Western blotting of the soluble and chromatin proteins of Mock and Flag-WRAP53β U2OS cells. Equal volumes from each fraction were loaded onto the gels. HSP90 and histone 4 were employed as markers for the soluble and chromatin fractions, respectively. The slower migration of the chromatin proteins of WRAP53β on the SDS gel compared to its soluble counterpart may be due to additional modifications of this protein when bound to chromatin. * indicates unspecific bands or bands of unknown origin
Immunostaining of WRAP53β and its interaction partner coilin (a marker for Cajal bodies) in control cells expressing endogenous WRAP53β revealed enrichment of both of these factors in Cajal bodies, as expected (Figure 1b).5 In contrast, no Cajal bodies were observed in the cells overexpressing WRAP53β, where this protein and coilin were distributed throughout the nucleoplasm (Figure 1b), in agreement with previous findings.5 Nor did reduction of background nucleoplasmic staining through pre-extraction of soluble proteins uncover any accumulation of WRAP53β or coilin in Cajal bodies in the cells overexpressing WRAP53β, although this treatment reduced total WRAP53β staining, potently indicating that most of the overexpressed protein is soluble (Figure 1c). Indeed, western blotting of the soluble and chromatin proteins of WRAP53β-overexpressing cells confirmed that most of the overexpressed WRAP53β was soluble, although the amount of this protein bound to chromatin also is increased (Figure 1d). Taken together, these findings demonstrate that overexpression of WRAP53β impairs accumulation of both this protein itself and coilin in Cajal bodies, with most of the overexpressed WRAP53β being in soluble form.
Overexpression of WRAP53β does not influence its recruitment to sites of DNA damage
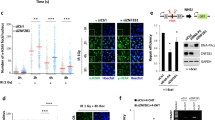
Next, we explored the localization of WRAP53β following exposure of cells to ionizing radiation (IR), which causes DNA double-strand breaks. WRAP53β localized rapidly to these breaks, to an extent of 84% in overexpressing cells in comparison with only 77% in the control cells (Figure 2a), which may simply reflect better visualization because of higher amounts of WRAP53β in the repair foci of the former. These foci also contained γH2AX, a marker for sites of DNA damage known to interact with WRAP53β (Figure 2a).9

WRAP53β accumulates at DNA double-strand breaks in cells overexpressing this protein. (a) Mock or Flag-WRAP53β cells were left untreated or irradiated (IR) with 6 Gy, and 1 h later, their soluble proteins were removed by pre-extraction and the cells were then immunostained for WRAP53β (with WRAP53-1F12 antibody) and γH2AX. The numbers represent the percentage of 100–200 cells counted whose nuclei contained >10 WRAP53β or γH2AX foci. Means±S.D. are shown, n=3. (b and c) After irradiation and pre-extraction as above, Flag-WRAP53β cells were immunostained for the proteins indicated. The FK2 antibody recognizes conjugated ubiquitin
Ubiquitylation of damaged chromatin, a process regulated by the ubiquitin ligases RNF8 and RNF168 in conjunction with WRAP53β,3 has an important role in the downstream recruitment of repair factors.3, 21, 22, 23, 24, 25 Repair foci containing WRAP53β were also enriched in conjugated ubiquitin (detected using the FK2 (antibody clone recognizing K29-, K48- and K63-linked polyubiquitylated and monoubiquitylated protein) antibody) (Figure 2b), as well as in tumor protein p53 binding protein 1 (53BP1) (Figures 2b and c), a downstream repair factor that requires ubiquitin signaling for its recruitment to DNA breaks.3 Thus, we conclude that WRAP53β-mediated targeting of repair factors to DNA double-strand breaks is not disturbed in cells that overexpress WRAP53β.
Repair of double-strand breaks in DNA is enhanced in cells that overexpress WRAP53β
Interestingly, in the cells overexpressing WRAP53β clearance of γH2AX-containing foci formed in response to irradiation was more rapid than in control cells (Figure 3a), indicating faster DNA repair. Moreover, comet assays revealed that the cells overexpressing WRAP53β contained fewer DNA breaks both 1 and 4 h after irradiation (Figure 3b).

Overexpression of WRAP53β enhances the repair of double-strand breaks. (a) The percentage of 200 Mock and Flag-WRAP53β cells counted, irradiated with 6 Gy and then fixed at the time-points indicated, whose nuclei contained >10 γH2AX foci. (b) Alkaline Comet assay of Mock and Flag-WRAP53β cells, either untreated (control) or exposed to 20 Gy IR, and then allowed to recover for 0, 1 or 4 h. The tail moment of 200 comets were analyzed for each experiment, n⩾3. (c) FACS analysis of HR and NHEJ efficiency following transient transfection of U2OS cells with the 3xFlag-Empty or 3xFlag-WRAP53β vectors for 48 h. In all cases, the values presented are means±S.D., n=3. *P<0.05, **P<0.01 and ***P<0.001, as determined by Student’s t-test
To examine whether more efficient DNA repair could explain these differences, we measured the efficiency of homologous recombination (HR) and non-homologous end-joining (NHEJ), repair pathways involved in the repair of double-strand breaks, using green fluorescent protein (GFP)-reporter assays.26, 27 In the case of HR, this assay involves U2OS cells carrying the direct repeat–GFP recombination construct, in which expression of exogenous I-SceI introduces a single double-strand break and repair of this break by HR produces a functional GFP. The NHEJ assay works in a similar manner, except that the GFP reporter (EJ5-GFP) contains two I-SceI sites, which upon cleavage generate two incompatible DNA ends repaired by NHEJ. Strikingly, transient overexpression of Flag-WRAP53β enhanced the efficiency of HR and NHEJ by ~4-fold (Figure 3c). Thus, we conclude that in cells overexpressing WRAP53β, HR and NHEJ are more efficient and consequently repair of DNA breaks more rapid.
Overexpression of WRAP53β enhances DNA repair by promoting RNF8-mediated ubiquitylation of histones
WRAP53β orchestrates repair of DNA double-strand breaks by recruiting the critical ubiquitin ligase RNF8 to DNA lesions.3 Ubiquitylation of histone H2AX, a known target of RNF8 at sites of DNA damage, was clearly elevated in cells overexpressing WRAP53β following irradiation (Figure 4a) and knockdown of RNF8 attenuated this effect (Figure 4b), suggesting that overexpression of WRAP53β stimulates RNF8-regulated ubiquitylation of damaged chromatin. In agreement with this idea, overexpression of RNF8 itself enhanced the efficiency of both HR and NHEJ repair to an extent similar to overexpression of WRAP53β (Figure 4c). Altogether, these findings indicate that overexpression of WRAP53β facilitates repair of DNA breaks by promoting RNF8-mediated ubiquitylation of associated histones, a process important for the assembly of downstream repair factors.

Overexpression of WRAP53β enhances histone ubiquitylation by RNF8. (a) Mock and Flag-WRAP53β cells were irradiated with 6 Gy and then allowed to recover for 1 h, after which western blotting for WRAP53β, H2AX and β-actin was performed. (b) Mock or Flag-WRAP53β cells were treated with the siRNAs indicated for 48 h, then irradiated with 6 Gy and allowed to recover for 1 h, after which western blotting for WRAP53β, RNF8, H2AX and β-actin was performed. (c) FACS analysis of HR and NHEJ efficiency following transient transfection of U2OS cells with the 1xFlag-Empty, 1xFlag-WRAP53β or 1xFlag-RNF8 vectors for 48 h. The values presented are means±S.D., n=3. **P<0.01 and ***P<0.001, as determined by Student’s t-test
Overexpression of WRAP53β promotes cells' survival following DNA damage
Tumors exhibiting altered responses to DNA damage can be either hypersensitive or resistant to genotoxic drugs. Interestingly, 70% of the control cells, but only 23% of those overexpressing WRAP53β, were apoptotic 5 days after irradiation (Figure 5a). The latter were also less sensitive to other genotoxic agents, including ultraviolet (UV) rays (42–47% reduction in the extent of apoptosis), mitomycin C (MMC; 68–72% decrease), hydroxyurea (HU; 59–68% decrease) and camptothecin (CPT; 66–73% decrease) (Figure 5b).

Cells that overexpress WRAP53β are less sensitive to apoptosis induced by DNA damage. (a) Mock and Flag-WRAP53β cells were irradiated (10 Gy), harvested 96 or 120 h later and analyzed by FACS for positive staining with Annexin V-PI, an indicator of apoptosis. (b) Mock and Flag-WRAP53β cells were treated with UV (30 J/m2), MMC (1 μg/ml), HU (500 μM), CPT (50 nM) or nothing (NT control) for 96 or 120 h, harvested and analyzed for Annexin V-PI positivity by FACS. (c) Mock and Flag-WRAP53β cells were left untreated (NT), irradiated (10 Gy) or treated with CPT (50 nM), harvested 72 h later and analyzed by western blotting for indicated proteins. (d) A total of 10 000 cells per ml of Mock and Flag-WRAP53β cells were seeded in 6-well plates, grown for 48 or 96 h, harvested and counted. The values presented are cells per ml. (e) Mock and Flag-WRAP53β cells were seeded, grown for 48 or 96 h, harvested, stained with PI and subjected to flow cytometry. The numbers indicate the % of cells in each cell cycle phase. Asterisk indicates the significance between the values in Flag-WRAP53β cells compared to the corresponding value in control cells. Percentages of sub-G1 and super-G2 are not shown. In all cases, the values presented are means±S.D., n=3. *P<0.05, **P<0.01 and ***P<0.001, as determined by Student’s t-test
Examination of the apoptotic response following DNA damage in these cells revealed reduced proapoptotic signaling (p53, PUMA, BAX) in cells overexpressing WRAP53β compared with control cells and upregulation of the antiapoptotic protein Bcl-2 in the former (Figure 5c), indicating that an altered apoptotic program may contribute to the resistance of WRAP53β-overexpressing cells to genotoxic agents.
Furthermore, we examined the growth rate of these cells, as DNA-damaging drugs most effectively eliminate rapidly dividing cells, while quiescent or slow-cycling cells are less susceptible.28 Interestingly, cells overexpressing WRAP53β grow slower than control cells (Figure 5d). The lower amount of cells overexpressing WRAP53β at 48 or 96 h after seeding compared with control cells was not because of apoptosis (Figure 5b – NT fraction) or cell cycle arrest (Figure 5e). However, the cell cycle distribution was slightly different between cells overexpressing WRAP53β and control cells, with more cells in the G1 phase and less in the S phase for the former, whereas the amount of cells in the G2 phase was the same for both cell lines. Taken together, our results demonstrate that overexpression of WRAP53β results in faster DNA repair, an altered apoptotic response and slower proliferation/cycling, thereby promoting cell survival following DNA damage.
Discussion
Although high levels of WRAP53β, a scaffold protein involved in the intracellular trafficking of factors to Cajal bodies and DNA breaks,3, 5, 29 are present in various cancers,16, 17, 18, 19, 20 it remains unclear how such overexpression may alter its functions. Here we report that overexpression of WRAP53β disturbs its recruitment and maintenance of factors to Cajal bodies, while at the same time enhancing the repair of DNA double-strand breaks. In cancer cells stably overexpressing this protein, WRAP53β was mislocalized in the nucleoplasm, in agreement with previous findings.5 Coilin, a marker protein for the Cajal body, demonstrated similar mislocalization, and thus no Cajal bodies appeared to have been formed. It is important to note that the overexpression in our cells was relatively high and that cells overexpressing WRAP53β at lower levels are still capable of forming Cajal bodies.5 Moreover, these cells only overexpress the open-reading frame of the protein and not the sequence of the WRAP53α transcript involved in regulating p53.
Intriguingly, in contrast to its absence from Cajal bodies, WRAP53β still localized to sites of DNA damage sites in overexpressing cells. Most of the overexpressed WRAP53β was soluble and may have competed with endogenous soluble WRAP53β for factors important for localization of this protein and associated factors. If soluble WRAP53β controls targeting of factors to Cajal bodies and the chromatin-bound fraction participates in DNA repair, this could explain why only its function related to the Cajal body are disrupted by overexpression.
Indeed, WRAP53β translocates certain components, including the SMN complex, from the cytoplasm to Cajal bodies,5 suggesting the involvement of a more mobile form of WRAP53β. Moreover, Cajal bodies alternate between passive diffusion within the nucleoplasm and transient immobilization through association with chromatin. In contrast, WRAP53β is recruited to sites of DNA damage from the nuclear pool alone, that is, no cytoplasmic shuttling is involved.4 In addition, many repair factors are bound to chromatin even before damage, allowing rapid redistribution to damaged sites when needed.
Our present observations also show that overexpression of WRAP53β enables more rapid repair of DNA double-strand breaks by HR and NHEJ. An open chromatin configuration allows more efficient repair of DNA double-strand breaks, providing greater resistance to such damage.30 Ubiquitylation of damaged chromatin is rate limiting for the accumulation of downstream repair factors,3, 21, 22, 23, 24 and, indeed, we found that overexpression of WRAP53β elevated ubiquitylation of H2AX. Knockdown of the ubiquitin ligase involved, RNF8, abrogated this effect in agreement with our previous report that WRAP53β targets RNF8 to DNA breaks.3
Rapid exchange of RNF8 at sites of DNA damage and the small number of copies of this protein in repair foci have been proposed to reflect its rate-limiting role in repair.31 Overexpression of WRAP53β may increase the level of RNF8 in repair foci by tethering it to damaged chromatin, thus slowing down its otherwise rapid turnover and promoting ubiquitylation. Our finding that overexpression of RNF8 itself enhanced HR and NHEJ repair supports the suggestion that elevating its local concentration may boost its action at DNA breaks. Moreover, RNF8 promotes decondensation of damaged chromatin by recruiting the chromatin-remodeling factor CHD4 (chromodomain helicase DNA-binding 4)32 and it is possible that WRAP53β aids in this decondensation, thereby enhancing chromatin accessibility to DNA repair. However, further investigation of the potential role of WRAP53β in the organization of chromatin is required.
Overexpression of WRAP53β also enhances cells survival following exposure to radiation and other DNA-damaging agents. We showed earlier that knockdown of WRAP53β in cancer cell lines, including the U2OS cells examined here, triggers mitochondria-dependent apoptosis and that overexpression of Bcl-2 protects against this apoptosis.16 Thus, Bcl-2 and/or other antiapoptotic proteins may be upregulated in cells that overexpress WRAP53β, protecting them from apoptosis. Indeed, we observed higher expression of Bcl-2 in these cells as well as diminished proapoptotic signaling. Moreover, we reveal that cells overexpressing WRAP53β proliferate significantly slower than cells with endogenous expression of this protein. Thus, it appears like cells overexpressing WRAP53β have acquired multiple resistance mechanisms, including increased DNA repair, overexpression of the antiapoptotic protein Bcl-2 and slow cycling, rendering them less sensitive to initiation of apoptosis by DNA damage.
The contradictory findings concerning the contribution of WRAP53β to cancer may involve timing. Inactivation could help initiate tumor development by impairing DNA repair and telomere maintenance and causing genomic instability. Indeed, inactivation of WRAP53β as a result of inherited mutations leads to the cancer susceptibility-disorder dyskeratosis congenita.7 In contrast, overexpression could be a late event in tumor progression, enhancing DNA repair in transformed cells suffering from replication stress and thereby promoting their maintenance and survival. Alternatively, high expression of WRAP53β observed in primary tumors could reflect accumulation of misfolded inactive WRAP53β, as mutations in this protein disrupt its folding by the chaperonin TRiC, which results in cytoplasmic accumulation and lost nuclear activity of mutated WRAP53β.9, 33 Similarly, p53 was initially believed to be an oncogene because of its overexpression in cancer, but later on found to be a tumor suppressor frequently inactivated in cancer, where the mutations cause overexpression of inactive p53 owing to reduced degradation of the protein.34 In any case, understanding the function of WRAP53β in health and diseases, such as cancer, may open novel therapeutic strategies.
Materials and Methods
Cells and culture conditions
Mock and Flag-WRAP53β U2OS cells were maintained in McCoy’s 5A medium (HyClone, Thermo Scientific, Stockholm, Sweden), selected with 10 μg/ml blasticidine S (InvivoGen), and DR-GFP and EJ5-GFP U2OS cells were maintained in high glucose DMEM (HyClone), supplemented with 10% fetal bovine serum (Hyclone) and 2.5 μg/ml plasmocin (InvivoGen) at 37 °C in 5% CO2 humidified chamber. Flag-WRAP53β cells overexpress the open-reading frame of the protein tagged with 1xFlag.
Antibodies
Primary
Rabbit α-WRAP53-C2 (cat. no. PA-2020-100; Innovagen AB, Lund, Sweden), mouse monoclonal α-WDR79 (clone 1F12, cat. no. H00055135-M04; Abnova, Stockholm, Sweden), mouse α-coilin (cat. no. sc-56298; Santa Cruz Biotechnology, Heidelberg, Germany), rabbit α-coilin (cat. no. sc-32860; Santa Cruz Biotechnology), mouse α-γH2AX (cat. no. 05-636; Millipore, Solna, Sweden), rabbit α-γH2AX (cat. no. 2577; Cell Signaling, Danvers, MA, USA), mouse α-conjugated ubiquitin (FK2) (cat. no. ST1200; Calbiochem, Millipore), rabbit α-53BP1 (cat. no. NB100-904; Novus Biologicals), rabbit α-H2AX (cat. no. ab11175; Abcam), mouse α-RNF8 (cat. no. sc-271462; Santa Cruz Biotechnology), rabbit α-H4 (cat. no. ab10158; Abcam), mouse α-p53 (cat. no. sc-126; Santa Cruz), rabbit α-PUMA (cat. no. ab9643; Abcam), rabbit α-BAX (cat. no. ab32503; Abcam), mouse α-Bcl-2 (cat. no. M0887; Dako), rabbit α-p21 (cat. no. ab109199; Abcam) and mouse heat-shock protein 90 (HSP90) α/β (cat. no. sc-13119; Santa Cruz Biotechnology).
Secondary
Goat α-rabbit HRP (cat. no. 7074; Cell Signaling), horse α-mouse HRP (cat. no. 7076; Cell Signaling), goat α-rabbit Alexa Fluor 488 (cat. no. A11008; Invitrogen, Thermo Scientific), goat α-mouse Alexa Fluor 488 (cat. no. A11029, Invitrogen, Thermo Scientific), donkey α-rabbit Alexa Flour 594 (cat. no. A21207; Invitrogen) and donkey α-mouse Alexa Fluor 594 (cat. no. A21203; Invitrogen).
Immunofluorescence microscopy
Cells grown on sterilized coverslips were fixed with 4% paraformaldehyde for 15 min at room temperature. They were then permeabilized with 0.1% Triton X-100 for 5 min at room temperature, followed by 30 min of blocking in blocking buffer (2% bovine serum albumin (BSA), 5% glycerol, 0.2% Tween-20, 0.1% NaN3). The coverslips were subsequently incubated for 1 h in primary antibody, followed by 40 min in secondary antibody, both diluted in blocking buffer, and finally mounted with Vectashield mounting medium containing DAPI (4′,6-diamidino-2-phenylindole; Vector Laboratories, Burlingame, CA, USA). Images were acquired with an LSM700 confocal microscope (Zeiss, Stockholm, Sweden), mounted on Axio observer.Z1 (Zeiss) equipped with Plan-Apochromat x63/1.4 oil immersion lens and processed with using Zen 2012 Black (Zeiss).
Pre-extraction
The cells were first washed with PBS and then incubated for 3 min at room temperature with cytoskeleton buffer (CSK) (10 mM Pipes, pH 7.0, 100 mM NaCl, 300 mM sucrose, 3 mM MgCl2 and 0.7% Triton X-100) and thereafter for another 3 min with the same CSK buffer supplemented with 0.3 mg/ml RNase A (CSK+R). Following these treatments, the cells were washed once again with PBS and then fixed in 4% paraformaldehyde.
Western blotting
Cells were harvested, washed and lysed in ice-cold lysis buffer (100 mM Tris-HCl, pH 8, 150 mM NaCl, 1% NP-40, 1% PMSF, 1% protease inhibitor cocktail) for 30 min on ice followed by sonication. The lysates obtained were centrifuged at 14 000 r.p.m. for 15 min at 4 °C and their protein concentrations determined with the Bradford assay (Bio-Rad). Thereafter, western blotting was performed by standard procedures.
Cell fractionation
Cells were harvested, washed in PBS and lysed in low salt buffer (10 mM HEPES, pH 7.4, 10 mM KCl, 0.05% Nonidet P-40, 1 mM DTT, 1% protease inhibitor cocktail) for 5 min on ice, followed by centrifugation at 3600 × g for 5 min at 4 °C. The supernatants were centrifuged again at 13 000 × g for 10 min at 4 °C. The supernatant (soluble proteins) was transferred to a new Eppendorf tube. The pellet from the first centrifugation were washed two times in low salt buffer with centrifugation at 3000 × g for 5 min at 4 °C between each wash. The remaining pellet were lysed in nuclease buffer supplemented with micrococcal nuclease (150 nM NaCl, 5 mM MgCl2, 1x micrococcal nuclease buffer, BSA, 2000 gel units micrococcal nuclease (cat. no. M0247S; New England Biolabs, Ipswich, MA, USA) for 10 min at 37 °C, followed by centrifugation at 13 000 × g for 10 min at 4 °C and transferred to a new Eppendorf tube (chromatin proteins). The soluble and chromatin fractions were adjusted to equal volumes. In Figure 1d, equal volumes of each fraction were loaded on an SDS-PAGE gel, corresponding to ~50 μg of soluble proteins and 3 μg of chromatin proteins.
Ionizing radiation
γ-Irradiation was performed with a 137Cs source (Scanditronix, Uppsala, Sweden) at the Karolinska Institutet (Stockholm, Sweden), at a photon dose rate of 0.5 Gy/min. Dosimetry was carried out with an ionization chamber as well as with ferro sulfate.
Comet assay
The alkaline Comet assay was performed using the method described earlier with some modifications.35 Briefly, cells were irradiated with 20 Gy. After 0, 1 or 4 h, cells were harvested with trypsin and washed. A total of 150 000 cells were diluted in 300 μl PBS, of which 100 μl were mixed with 500 μl low-melting-point agarose (1.2% (w/v) in PBS) to a final concentration of 1%. Of the resulting suspension, 100 μl was layed on the top of a previously prepared normal-melting-point agarose (1% (w/v) in PBS) on fully-frosted slides and the suspension was covered with a 22x22 mm2 coverslip. The slides were put on ice for 10 min and then immersed in lysis buffer (2.5 M NaCl, 100 mM sodium-EDTA, 10 mM Tri-HCl, pH 10, 10% DMSO, 1% Triton X-100) for overnight at 4 °C in the dark. After lysis, slides were placed in alkaline electrophoresis buffer (0.3 M NaOH, 1 mM sodium-EDTA) for 30 min at room temperature to denature DNA and express alkali-labile sites. Electrophoresis was carried out at room temperature for 30 min at 25 V, 300 mA in a Comet Assay Tank (Thistle Scientific, Glasgow, UK). The slides were then washed two times in neutralizing buffer (0.4 M Tris-HCl, pH 7.4) for 30 min. DNA was stained with 40 μl CybrGOLD diluted 1 : 1000 (Thermo Scientific). Two slides were run and analyzed for each experiment. On each slide, 100 comets were quantified at × 10 magnification using the Comet Assay IV Image analysis system (Perceptive Instruments, Bury Saint Edmunds, UK) in live-video mode on a Zeiss Axiovert 35 fluorescent microscope (Zeiss). Tail moment (arbitrary unit) is defined as the product of the tail length and the fraction of total DNA in the tail. Images were acquired using the same software. For the statistical analysis, a two-way ANOVA analysis was performed in GraphPad Prism software (GraphPad Software, La Jolla, CA, USA).
Drug treatment
In all, 1 μg/ml MMC (cat. no. 10107409001; Roche, Stockholm, Sweden), 500 μM HU (cat. no. H8627; Sigma-Aldrich), 50 nM CPT (cat. no. C9911; Sigma-Aldrich, Stockholm, Sweden) were added to cells for 96 or 120 h.
FACS analysis
For the analysis of apoptosis, cells were exposed to genotoxic agents and harvested at the indicated time-points with trypsin. Next, the samples were incubated with incubation buffer (10 mM HEPES (pH 7.4), 140 mM NaCl, 2.5 mM CaCl2) supplemented with Annexin-V-FLUOS (cat. no. 11828681001; Roche) and 0.1% propidium Iodide (PI) solution (cat. no. P4864; Sigma-Aldrich) for 15 min at room temperature. The cells were then dissolved in incubation buffer and analyzed for active Annexin V-PI staining by flow cytometry on a FACS Calibur (Becton Dickinson, Erembodegem, Belgium) using the Cell Quest software (Becton Dickinson). The sum of the Annexin V+-P− (early apoptosis) and Annexin V+-PI+ (late apoptosis) population of cells were accounted as apoptotic cells.
For the analysis of the cell cycle, cells were seeded and 48 or 96 h later harvested with trypsin, washed in PBS and fixed in 60% ethanol overnight at 4 °C. After removal of the ethanol, the samples were incubated with a solution of RNase A/PI for 30 min at 37 °C and subsequently analyzed with a NovoCyte apparatus (ACEA Biosciences Inc., San Diego, CA, USA).
HR and NHEJ assays
A total of 300 000 cells were seeded into 6-well plates. After 24 h, cells were transfected with an I-SceI vector together with a vector expressing Flag-Empty, Flag-WRAP53β or Flag-RNF8 using Lipofectamine 2000 (Invitrogen). 1xFlag vector corresponds to pCMV-Tag2 vector (Invitrogen) and 3xFlag vector to p3xFlag-CMV-9 (Sigma-Aldrich). The next day, media were changed, and 24 h after this, cells were harvested by trypsination, washed with PBS and the GFP signal arising from the recombination event was measured by flow cytometry on a FACS Calibur (as described above), with fluorescence detected in the FL1-H channel (logarithmic scale). The frequency of repair in cells transfected with the various plasmids was calculated relative to cells transfected with the empty plasmid. Each data point represents the mean±S.D. from three independent experiments.
Cell count analysis
Cells were seeded and 48 or 96 h later harvested and counted using a Bürker chamber Brand (Sigma-Aldrich).
siRNA transfections
Ten nanomoles of small interference RNA (siRNA), siRNF8 (cat. no. L-006900-00-0005; Dharmacon, Karlskoga, Sweden) or siControl (cat. no. 1027280, Qiagen), was transfected into cells using HiPerfect (Qiagen, Sollentuna, Sweden) transfection reagent in accordance with the supplier’s recommendations.
Abbreviations
- WRAP53:
-
WD40 encoding RNA antisense to p53
- HR:
-
homologous recombination
- NHEJ:
-
non-homologous end-joining
- IR:
-
ionizing irradiation
- DAPI:
-
4′,6-diamidino-2-phenylindole
- CSK:
-
cytoskeleton buffer
- siRNA:
-
small interference RNA
- γH2AX:
-
phospho-histone H2AX
- RNF8:
-
ring-finger protein 8
- 53BP1:
-
tumor protein p53 binding protein 1
- FK2:
-
antibody clone recognizing K29-, K48- and K63-linked polyubiquitylated and monoubiquitylated protein
- SMN:
-
survival of motor neuron
- UV:
-
ultraviolet
- MMC:
-
mitomycin C
- HU:
-
hydroxyurea
- CPT:
-
camptothecin
- PI:
-
propidium iodide
- HSP90:
-
heat-shock protein 90
- U2OS:
-
U-2 osteosarcoma
- BSA:
-
bovine serum albumin
References
Mahmoudi S, Henriksson S, Corcoran M, Mendez-Vidal C, Wiman KG, Farnebo M . Wrap53, a natural p53 antisense transcript required for p53 induction upon DNA damage. Mol Cell 2009; 33: 462–471.
Venteicher AS, Abreu EB, Meng Z, McCann KE, Terns RM, Veenstra TD et al. A human telomerase holoenzyme protein required for Cajal body localization and telomere synthesis. Science 2009; 323: 644–648.
Henriksson S, Rassoolzadeh H, Hedstrom E, Coucoravas C, Julner A, Goldstein M et al. The scaffold protein WRAP53beta orchestrates the ubiquitin response critical for DNA double-strand break repair. Genes Dev 2014; 28: 2726–2738.
Hedstrom E, Pederiva C, Farnebo J, Nodin B, Jirstrom K, Brennan DJ et al. Downregulation of the cancer susceptibility protein WRAP53beta in epithelial ovarian cancer leads to defective DNA repair and poor clinical outcome. Cell Death Dis 2015; 6: e1892.
Mahmoudi S, Henriksson S, Weibrecht I, Smith S, Soderberg O, Stromblad S et al. WRAP53 is essential for Cajal body formation and for targeting the survival of motor neuron complex to Cajal bodies. PLoS Biol 2010; 8: e1000521.
Tycowski KT, Shu MD, Kukoyi A, Steitz JA . A conserved WD40 protein binds the Cajal body localization signal of scaRNP particles. Mol Cell 2009; 34: 47–57.
Zhong F, Savage SA, Shkreli M, Giri N, Jessop L, Myers T et al. Disruption of telomerase trafficking by TCAB1 mutation causes dyskeratosis congenita. Genes Dev 2011; 25: 11–16.
Zhong FL, Batista LF, Freund A, Pech MF, Venteicher AS, Artandi SE . TPP1 OB-fold domain controls telomere maintenance by recruiting telomerase to chromosome ends. Cell 2012; 150: 481–494.
Rassoolzadeh H, Coucoravas C, Farnebo M . The proximity ligation assay reveals that at DNA double-strand breaks WRAP53beta associates with gammaH2AX and controls interactions between RNF8 and MDC1. Nucleus 2015; 6: 417–424.
Garcia-Closas M, Kristensen V, Langerod A, Qi Y, Yeager M, Burdett L et al. Common genetic variation in TP53 and its flanking genes, WDR79 and ATP1B2, and susceptibility to breast cancer. Int J Cancer 2007; 121: 2532–2538.
Medrek K, Magnowski P, Masojc B, Chudecka-Glaz A, Torbe B, Menkiszak J et al. Association of common WRAP 53 variant with ovarian cancer risk in the Polish population. Mol Biol Rep 2013; 40: 2145–2147.
Schildkraut JM, Goode EL, Clyde MA, Iversen ES, Moorman PG, Berchuck A et al. Single nucleotide polymorphisms in the TP53 region and susceptibility to invasive epithelial ovarian cancer. Cancer Res 2009; 69: 2349–2357.
Lan Q, Zhang L, Shen M, Jo WJ, Vermeulen R, Li G et al. Large-scale evaluation of candidate genes identifies associations between DNA repair and genomic maintenance and development of benzene hematotoxicity. Carcinogenesis 2009; 30: 50–58.
Garvin S, Tiefenbock K, Farnebo L, Thunell LK, Farnebo M, Roberg K . Nuclear expression of WRAP53beta is associated with a positive response to radiotherapy and improved overall survival in patients with head and neck squamous cell carcinoma. Oral Oncol 2015; 51: 24–30.
Silwal-Pandit L, Russnes H, Borgen E, Skarpeteig V, Moen Vollan HK, Schlichting E et al. The sub-cellular localization of WRAP53 has prognostic impact in breast cancer. PLoS One 2015; 10: e0139965.
Mahmoudi S, Henriksson S, Farnebo L, Roberg K, Farnebo M . WRAP53 promotes cancer cell survival and is a potential target for cancer therapy. Cell Death Dis 2011; 2: e114.
Sun CK, Luo XB, Gou YP, Hu L, Wang K, Li C et al. TCAB1: a potential target for diagnosis and therapy of head and neck carcinomas. Mol Cancer 2014; 13: 180.
Rao X, Huang D, Sui X, Liu G, Song X, Xie J et al. Overexpression of WRAP53 is associated with development and progression of esophageal squamous cell carcinoma. PLoS One 2014; 9: e91670.
Sun Y, Yang C, Chen J, Song X, Li Z, Duan M et al. Overexpression of WDR79 in non-small cell lung cancer is linked to tumour progression. J Cell Mol Med 2016; 20: 698–709.
Zhang H, Wang DW, Adell G, Sun XF . WRAP53 is an independent prognostic factor in rectal cancer – a study of Swedish clinical trial of preoperative radiotherapy in rectal cancer patients. BMC Cancer 2012; 12: 294.
Kolas NK, Chapman JR, Nakada S, Ylanko J, Chahwan R, Sweeney FD et al. Orchestration of the DNA-damage response by the RNF8 ubiquitin ligase. Science 2007; 318: 1637–1640.
Marteijn JA, Bekker-Jensen S, Mailand N, Lans H, Schwertman P, Gourdin AM et al. Nucleotide excision repair-induced H2A ubiquitination is dependent on MDC1 and RNF8 and reveals a universal DNA damage response. J Cell Biol 2009; 186: 835–847.
Huen MS, Grant R, Manke I, Minn K, Yu X, Yaffe MB et al. RNF8 transduces the DNA-damage signal via histone ubiquitylation and checkpoint protein assembly. Cell 2007; 131: 901–914.
Mailand N, Bekker-Jensen S, Faustrup H, Melander F, Bartek J, Lukas C et al. RNF8 ubiquitylates histones at DNA double-strand breaks and promotes assembly of repair proteins. Cell 2007; 131: 887–900.
Doil C, Mailand N, Bekker-Jensen S, Menard P, Larsen DH, Pepperkok R et al. RNF168 binds and amplifies ubiquitin conjugates on damaged chromosomes to allow accumulation of repair proteins. Cell 2009; 136: 435–446.
Pierce AJ, Johnson RD, Thompson LH, Jasin M . XRCC3 promotes homology-directed repair of DNA damage in mammalian cells. Genes Dev 1999; 13: 2633–2638.
Gunn A, Stark JM . I-SceI-based assays to examine distinct repair outcomes of mammalian chromosomal double strand breaks. Methods Mol Biol 2012; 920: 379–391.
Moore N, Houghton J, Lyle S . Slow-cycling therapy-resistant cancer cells. Stem Cells Dev 2012; 21: 1822–1830.
Henriksson S, Farnebo M . On the road with WRAP53beta: guardian of Cajal bodies and genome integrity. Front Genet 2015; 6: 91.
Murga M, Jaco I, Fan Y, Soria R, Martinez-Pastor B, Cuadrado M et al. Global chromatin compaction limits the strength of the DNA damage response. J Cell Biol 2007; 178: 1101–1108.
Mok MT, Cheng AS, Henderson BR . The ubiquitin ligases RNF8 and RNF168 display rapid but distinct dynamics at DNA repair foci in living cells. Int J Biochem Cell Biol 2014; 57: 27–34.
Luijsterburg MS, Acs K, Ackermann L, Wiegant WW, Bekker-Jensen S, Larsen DH et al. A new non-catalytic role for ubiquitin ligase RNF8 in unfolding higher-order chromatin structure. EMBO J 2012; 31: 2511–2527.
Freund A, Zhong FL, Venteicher AS, Meng Z, Veenstra TD, Frydman J et al. Proteostatic control of telomerase function through TRiC-mediated folding of TCAB1. Cell 2014; 159: 1389–1403.
Oren M, Rotter V . Introduction: p53 – the first twenty years. Cell Mol Life Sci 1999; 55: 9–11.
Sasaki YF, Saga A, Akasaka M, Yoshida K, Nishidate E, Su YQ et al. In vivo genotoxicity of ortho-phenylphenol, biphenyl, and thiabendazole detected in multiple mouse organs by the alkaline single cell gel electrophoresis assay. Mutat Res 1997; 395: 189–198.
Acknowledgements
This work was supported by grants from the Swedish Cancer Society (Cancerfonden), the Swedish Research Foundation (VR), the Strategic Research Programme in Cancer (StratCan), Worldwide Cancer Research (former AICR), the Swedish Childhood Cancer Society (Barncancerfonden), the Cancer Society of Stockholm (Cancerföreningen), Olle Engkvist Byggmästare Foundation and the Karolinska Institutet.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Edited by J Chipuk
Rights and permissions
Cell Death and Disease is an open-access journal published by Nature Publishing Group. This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Rassoolzadeh, H., Böhm, S., Hedström, E. et al. Overexpression of the scaffold WD40 protein WRAP53β enhances the repair of and cell survival from DNA double-strand breaks. Cell Death Dis 7, e2267 (2016). https://doi.org/10.1038/cddis.2016.172
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cddis.2016.172
This article is cited by
-
Small Cajal body-associated RNA 2 (scaRNA2) regulates DNA repair pathway choice by inhibiting DNA-PK
Nature Communications (2022)
-
Suppression of TCAB1 expression induced cellular senescence by lessening proteasomal degradation of p21 in cancer cells
Cancer Cell International (2021)
-
Non-canonical roles of canonical telomere binding proteins in cancers
Cellular and Molecular Life Sciences (2021)
-
Biallelic mutations in WRAP53 result in dysfunctional telomeres, Cajal bodies and DNA repair, thereby causing Hoyeraal–Hreidarsson syndrome
Cell Death & Disease (2020)
