
Abstract
Background:
Hypoxia and the subsequent activation of hypoxia-inducible factor-2α (HIF2α) contribute to the progression of a variety of cancers. However, their role in the generation of renal cell carcinoma-derived stem cells has not been fully addressed.
Methods:
A sphere formation assay, cell proliferation, RT–PCR, western blot, FACS, immunohistochemistry and tumour xenograft were used to study the role of HIF2α.
Results:
Propagation of four renal cell carcinoma (RCC) cell lines (Caki-1, Caki-2, 786-O, 769-P) in anchorage-independent floating spheres led to the expansion of cells bearing the CXCR4 (CD184) surface marker. Inhibition of the CXCR4 pathway reduced sphere expansion. The enhanced self-renewal activity of the CXCR4-positive spheres was preceded by the upregulation of HIF2α. Knockdown of HIF2α abrogated CXCR4 expression and sphere formation. Finally, RCC-derived spheres showed an undifferentiated phenotype in vivo and formed subcutaneous tumours that highly expressed HIF2α and CXCR4. Inhibition of HIF2α abolished tumour growth in animal models.
Conclusions:
These results suggest that the generation of RCC-derived CSCs involves the activation of HIF2α and may provide a foundation for the development of new strategies to prevent the induction of CSCs in RCC.
Similar content being viewed by others

Main
Renal cell carcinoma (RCC) is the most common epithelial malignancy of human adult kidneys (McLaughlin and Lipworth, 2000; Siegel et al, 2012; Khan et al, 2013). Approximately 30% of RCC patients present metastatic disease with a 5-year survival probability of 10%. This is owing to the high incidence of toxic effects and resistance to therapies (Ljungberg 2004; Siegel et al, 2012). Therefore, identifying novel targets is required for future therapeutic interventions.
Increasing evidence suggests that the presence of a small minority of cells within tumours termed tumour-initiating cells or cancer stem cells (CSCs) might be a reason for this resistance (Reya et al, 2001; Zhou et al, 2009; Vermeulen et al, 2012; Khan et al, 2013). This small subset of cancer cells has been identified in many solid tumours, including the brain (Singh et al, 2003; Clarke et al, 2006), breast (Al-Hajj et al, 2003), and colon (Ricci-Vitiani et al, 2007; Kaur et al, 2014). The CSCs show a high self-renewal capacity, as demonstrated by the growth of tumour spheres in vitro and the tumorigenic and metastatic potential following inoculation into non-obese diabetic/severe combined immunodeficiency (NOD/SCID) mice (Pan et al, 2010).
The CSCs have also been identified and isolated from RCC patients and cell lines using several stem cell markers, including the chemokine receptor CXCR4 (Zagzag et al, 2005; Gelmini et al, 2008; Gassenmaier et al, 2013), CD105 (Bussolati et al, 2008) and aldehyde dehydrogenase 1 (ALDH1) (Ueda et al, 2013). Among these, CXCR4-positive cells exhibited great resistance to tyrosine kinase inhibitors and expressed other ‘stemness’-associated genes. Notably, surgical biopsies of RCC patients confirmed that the high expression of CXCR4 has significant prognostic value and therapeutic importance (D’Alterio et al, 2010). However, further characterisation of CSCs is required to fully assess the role of these cells in RCC genesis and their impact on cancer therapy.
Recent findings showed that the hypoxia-inducible factors (HIFs), HIF1α and HIF2α in particular, could promote the generation of CSCs (Heddleston et al, 2009). HIF2α has been shown to activate Oct4 and control primordial germ cells (Covello et al, 2006). HIF1α can also regulate the stem cell properties of colon and breast cancer cells (Chiavarina et al, 2012). In RCCs, clinical data indicated that HIF2α overexpression was associated with disease progression and mortality whereas HIF1α expression was silenced secondary to von Hippel–Lindau (VHL) mutations that occurred in over 90% of clear cell RCCs (Kaelin, 2008; Keith et al, 2012). However, how HIFs contribute to CSCs generation in RCC is not fully understood.
Here, we show that tumour spheres with self-renewal capacity from some RCC cell lines could be a source for enrichment of renal CSCs. Moreover, HIF2α is essential in the induction of CSC-like properties through CXCR4 expression. Altogether, our data suggest that future therapies could combine blockade of the HIF2α signalling pathway with molecular therapies for more effective treatments of metastatic RCC.
Materials and Methods
Antibodies and reagents
Antibodies purchased for these studies included HIF1α (Chemicon MAB5382, Darmstadt, Germany), HIF2α (Origene TA309641, Rockville, MD, USA) and CXCR4 (Biorbyt orb74308, Cambridge, UK). Other purchased reagents included a CXCR4 inhibitor (AMD3100; Sigma A5602, St Louis, MO, USA), biotinylated anti-rabbit IgG (BA-1000), biotinylated anti-goat IgG (BA-5000), and Vectastain ABC Kit (Vector Laboratories, Burlingame, CA, USA), Texas Red Conjugated goat Anti-Rabbit IgG (Thermo Scientific 31506, Waltham, MA, USA) and FITC- rabbit IgG (Sigma F9887).
Cell lines
Human RCC cell lines (786-O, Caki-1, 769-P, and Caki-2) were obtained from the American Type Culture Collection (ATCC). The cell lines 786-O and 769-P were grown in RPMI-1640, whereas Caki-1 and Caki-2 were grown in McCOY’s 5A supplemented with 10% FBS at 37 °C in a humidified 5% CO2-containing atmosphere. To obtain chemical hypoxia, a concentration of 500 μ M cobalt(II) chloride (CoCl2; Sigma 409332) was added to the medium, and the cells were treated for 24 h.
Sphere formation assay
As reported by Zhong et al (2010), single cells were seeded on ultra-low attachment plates (Corning, Corning, NY, USA) at a concentration of 2 × 105 cells ml−1 in serum-free medium (DMEM/F12) supplemented with B27 (Gibco 17504-044), EGF (20 ng ml−1, PeproTech AF-100-15) and FGF (20 ng ml−1, PeproTech 100-18B). The growth factor-responsive cells proliferated and formed floating spheres, whereas most of the differentiated cells rapidly died. The first generation spheres were collected after 7 days of culture. Spheres were dissociated into a single-cell suspension with trypsin and were then cultured again to promote further generations. After 14 and 21 days, we collected the second- and third-generation spheres, respectively, to study self-renewal capacity. The second generation cells were used for RT–PCR and in vitro assays. To analyse the cell viability before each experiment, the number and size distribution of cells were measured with a portable cell counter, Scepter Handheld Automated Cell Counter (PHCC20060 Scepter, Merck Millipore, Billerica, MA USA).
Differentiation assay
For adipogenic differentiation, sphere-derived cells from 786-O, 769-P, Caki-2, and Caki-1 were seeded at 5 × 104 on six-well plates in adipogenic medium containing complete RPMI-1640 or McCOY’s 5A with 2 mM L-glutamine, 100 U ml−1 penicillin, 100 mg ml−1 streptomycin, and 10% FBS supplemented with 0.5 μ M dexamethasone, 5 μg ml−1 insulin, 0.2 mM indomethacin, and 0.45 mM isobutylmethylxanthine (Sigma-Aldrich, St Louis, MO, USA). Adipogenic medium was changed twice a week until the end of the experiment. The cell lines 786-O, 769-P, Caki-2, and Caki-1 were cultured in complete RPMI-1640 and McCOY’s 5A, respectively, or without adipogenic supplements as a control.
Flow cytometry
To analyse the expression of CXCR4 in 786-O and Caki-1 by flow cytometry, cells were washed twice with ice-cold PBS and then incubated with the FITC-conjugated primary antibody for 30 min on ice. Cells were then washed twice with PBS and were analysed using a FACSCalibur flow cytometer (BD Pharmingen, Erembodegem, Belgium).
RT–PCR analysis
Total RNA was isolated from spheres and adherent cells by the Qiagen RNase Extraction Kit (Qiagen 74104, Milan, Italy). We used 500 ng of total RNA from each sample for cDNA synthesis by reverse transcription (RT) with the Fermentas kit (#K1631) in a total volume of 20 μl. We used 0.5 μl of RT product for real-time PCR using the SYBR Green method. As a housekeeping gene, β-actin was used. The genes studied were CXCR4, SDF-1, VEGF, Nanog, CD44, CD105, Oct4, CD133, and ALDH with the following primers: CXCR4 forward 5′-TCTTCCTGCCCACCATCTAC-3′; reverse 5′-CAGCCTGTACTTGTCCGTCAT-3′; SDF-1 forward 5′-CGTCAGCCTG AGCTACAGAT-3′ reverse 5′-AGCCGGGCTA CAATCTGAAG-3′; VEGF forward 5′-CCGAAACCATGAACTTTCTGC-3′ reverse 5′-CCATGAACTTCACCACTTCGT-3′; Nanog forward 5′-GATTTGTGGGCCTGAAGAAA-3′ reverse 5′-AAGTGGGTTGTTTGCCTTTG-3′; CD44 forward 5′-CCTGCAGGTATGGGTTCATAG-3′ reverse 5′-GTGTCATACTGGGAGGTGTTG-3′; ALDH forward 5′-TCCTGGTTATGGGCCTACAG-3′ reverse 5′-CTGGCCCTGGTGGTAGAATA-3′; CD133 forward 5′-GGTGCATCCATCAAGTGAAACC-3′ reverse 5′-GGATTGATAGCCCTGTTGGAC-3′; OCT4 forward 5′-AGTGAGAGGCAACCTGGAGA-3′ reverse 5′-ACACTCGGACCACATCCTTC-3′; CD105 forward 5′-AAGCAAAATGGCACCTGGC-3′ reverse 5′-GCACTTGGCC TACAATTCCA-3′.
Western blot
Whole-cell lysates from spheres and adherent cells from 786-O and Caki-1 were obtained with RIPA buffer containing a protease inhibitor cocktail. Proteins were resolved on 10% SDS-polyacrylamide gels and electroblotted onto nitrocellulose membranes. After transfer, blocking of unspecific binding sites was achieved by incubation in PBS-Tween containing 5% skimmed milk. For the incubation steps with primary and HRP-conjugated secondary antibodies, the antibody concentration was used as recommended by the manufacturer. Detection was performed using an ECL kit (Merck Millipore).
Transfection with shRNA plasmids
Cells were grown on six-well plates in normal growth medium without antibiotics, and the X-tremeGENE HP (Roche, Milan, Italy) transfection reagent at a ratio 3:1 (μl reagent per μg DNA) was used as suggested by the manufacturer. HIF2α-shRNA was obtained from OriGene TG315484; we used 1 μg. The efficacy of transfection was analysed 24 h post transfection and was confirmed by GFP fluorescence. Seventy-two hours after transfection, we added puromycin (1 μg ml−1) to obtain a stable culture.
Tumour induction in NOD/SCID mice
Before subcutaneous injection in NOD/SCID mice, spheres were dissociated to single cells through enzymatic and mechanical dissociation and were then resuspended in PBS. We performed two limited dilutions in in vivo experiments: in the right flank of two different mice, we injected 5 × 104 and 3 × 106 sphere-derived cells, and in the other flank, we injected the same number of adherent cells (both 786-O and Caki-1). In a second experiment, we injected 3 × 106 786-O sh-Empty cells in the right flank, and we injected the same amount of 786-O sh-HIF2α in the other side. In the last in vivo experiment, we injected 5 × 104 786-O sh-Empty sphere-derived cells in the right flank, and we injected the same number of 786-O sh-HIF2α sphere-derived cells in the other flank. Injection was performed in mice that were anaesthetised with 2,2,2-tribromoethanol (Sigma 90710) 97%. Tumour growth was monitored weekly, and tumour size was measured using a digital calliper; the volume was calculated as 4/3 πr3.
Immunohistology
We sequentially incubated 4-μm sections in xylene (5 min twice), 100% ethanol (5 min), 95% ethanol (5 min), 70% ethanol (5 min), and then water. Subsequently, the sections were antigen-retrieved using 0.25 mM EDTA buffer (pH 6.0 Dako) in a steamer for 20 min and then cooled to ambient temperature. Sections were then washed with PBS and quenched with 3% hydrogen peroxide in methanol for 10 min, blocked for avidin/biotin activity and incubated with primary antibodies as follows: for HIF1α (1:500), HIF2α (1:500), and CXCR4 (1:1000), the sections were incubated at 4 °C overnight. After primary antibody incubation, the sections were washed with PBS, incubated with biotinylated anti-rabbit or biotinylated anti-goat IgG (1:200) for 30 min and then washed and incubated with ABC-horseradish peroxidase. Antibody binding was visualised with diaminobenzidine and counterstained with haematoxylin. Finally, the sections were dehydrated through graded alcohol, cleared in xylene, and cover-slipped.
Analysis of expression data of HIF2α and CXCR4 in human renal cancer
For the human gene expression data, we took advantage of the publicly available gene expression data sets in the NextBio Research bioinformatics tool. We searched for gene names in the ‘Disease Atlas’ section and used a filter to select RNA selection studies in human kidney cancers.
Statistical analysis
The data are expressed as the mean±s.d. Significant differences between groups were analysed using Student’s t-test. The difference was considered statistically significant at a P value<0.05. To obtain comparable results, the data of at least three mice from each group were used in the statistical analysis.
Results
Characterisation of a panel of human RCC cell lines grown in non-adherent conditions
We used a panel of clear cell-RCC (CC-RCC) cell lines, including Caki-1, 769-P, 786-O, and Caki-2 (Shinojima et al, 2007). As described in the Sanger Institute COSMIC database, Caki-1 cells were wild type, whereas the other cell lines were mutated for the VHL gene. Moreover, Caki-1 cells showed HIF1α expression only when they were treated with the hypoxia-mimetic agent cobalt(II) chloride (CoCl2), whereas 769-P and Caki-2 cells showed basal expression of HIF1α (data not shown). No expression of HIF1α was observed in 786-O, which was consistent with a report showing that 786-O is a VHL-deficient RCC cell line that constitutively expressed only HIF2α (Soeda et al, 2008; Supplementary Figure S1).
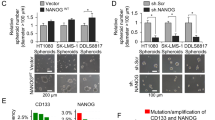
We analysed the self-renewal capacities of these cells by performing a sphere-forming assay (Keith and Simon, 2007). We found that 786-O and Caki-1 cell lines have sphere-forming abilities compared with 769-P and Caki-2 which are not able to form the three-dimensional structures that are ascribable to spheres (Figure 1A). To test whether the sphere-derived cells have stem/progenitor properties, we performed a differentiation assay based on cultivating cells for 10 days in differentiating conditions, such as the presence of adipogenic medium. Sphere-derived cells from the Caki-1 and 786-O cell lines were able to differentiate into adipocytes, in contrast to 769-P and Caki-2 cells (Figure 1B), which suggests that Caki-1- and 786-O-derived spheres showed a CSC-like phenotype that was independent of VHL and HIF1α status.

Characterisation of 786-O, Caki-1, Caki-2, and 769-P cell lines grown in non-adherent conditions. (A) 786-O and Caki-1 cell lines were able to form spheres in non-adherent conditions compared with the Caki-2 and 769-P cell lines. Qualitative and quantitative representation of this phenomenon. (B) 786-O and Caki-1 sphere-derived cells were able to differentiate into adipocytes. (C) Real-time PCR with stem cell markers on the CSC-like subpopulation of 786-O and Caki-1 cell lines compared with adherent cells. (D) Reduction of the mean number of spheres after treatment with the CXCR4 inhibitor (AMD3100). *P<0.0038, **P<0.0001.
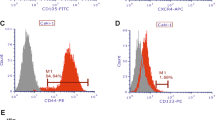
To further characterise the phenotype of the CSC-like subpopulation, we performed real-time PCR analysis of sphere-derived mRNAs with a series of markers associated with stem/progenitor and cancer cells. Propagation in non-adherent condition led to an increase in the number of cells bearing CXCR4 and its ligand SDF-1 in both 786-O- and Caki-1-derived spheres, with an increase in Nanog-expressing cells observed only in 786-O-derived spheres (Figure 1C). No significantly increased expression in stem cell markers was observed in the Caki-2 and 769-P cell lines (data not shown). Flow cytometry (FACS) analysis confirmed the enhanced proportion of CXCR4-positive cells in both 786-O- and Caki-1-derived spheres compared with the same cell lines cultured in adhesion (Supplementary Figure S2). These results showed that the sphere-forming assay preferentially expands a CXCR4-positive subpopulation of RCC cells with CSC-like properties.
We then used the CXCR4 inhibitor (AMD3100) at a concentration of 10 μ M to further investigate the role of CXCR4 in promoting the sphere-forming capacity of both 786-O and Caki-1 RCC cell lines. Inhibition of the CXCR4 pathway markedly inhibited the expansion of CSC-like cells (Figure 1D). These data suggest that inhibiting the CXCR4 signalling pathway may attenuate the expansion of CXCR4-positive RCC cells with CSCs properties.
786-O and Caki-1 sphere-derived cells demonstrate increased tumorigenicity
To analyse the tumorigenicity of the RCC cells that had undergone CSC-like properties, we injected sphere-derived cells in limiting dilutions into subcutaneous sites of NOD/SCID mice. All the mice that were injected with 5 × 104 cells derived from spheres of 786-O cells formed tumours, whereas no masses occurred when an equal number of cells derived from standard culture conditions were injected in the other flank of the same mouse (Table 1). In fact, 3 × 106 of the parental cells (grown under adherent conditions) were required to initiate tumour formation. Similarly, sphere-derived Caki-1 cells at a density of 5 × 104 formed tumours, whereas Caki-2 cells did not (Table 1). Therefore, the sphere-forming assay significantly increased (by approximately two orders of magnitude) the number of tumour-initiating cells. Notably, the histology of the tumours developed from the cells derived from spheres appeared to be similar to that of the tumours formed by the parental cells inoculated at high density. However, tumours derived from 786-O spheres present higher expression of both CXCR4 and HIF2α (Supplementary Figure S3) than tumours derived from adherent cells where we found a not detectable or very low expression of both. It can be owing to the strong dilution of CSC subpopulation in 786-O adherent cell-derived tumours.
CSCs-like RCC cells expressed HIF2α
Because HIFs are important transcription factors that are involved in cancer progression, we hypothesised that RCC cell lines may acquire CSCs-like properties and the expression of CXCR4 by stabilising HIF2α expression. In tumour spheres derived from Caki-1 and 786-O cells, we found a high expression of HIF2α by western blotting (Figure 2A). Interestingly, there was no difference in HIF1α expression. Moreover, because vascular endothelial growth factor (VEGF) is an important target gene that is involved in the angiogenic response to HIFs, we determined the VEGF mRNA level in sphere-derived cells by real-time PCR. The VEGF mRNA level was nearly threefold higher in sphere-derived cells than in cells cultured under adherent conditions (Figure 2B), suggesting HIF2α stabilisation and activation in tumour spheres.

CSCs-like RCC cells express HIF2 α. (A) Western blot showing a high expression of HIF2α in 786-O and Caki-1 cell lines cultured in adherent and sphere conditions. No expression was reported for HIF1α in both cell lines. (B) Real-time PCR for relative VEGF mRNA expression in 786-O and Caki-1 cell lines in adherent and sphere conditions. In both the 786-O and Caki-1 cell lines, VEGF expression was higher in spheres compared with adherent cells. *P<0.0003 and **P<0.0001. (C) Immunohistochemical analysis of HIF1α and HIF2α expression on 786-O and Caki-1 sphere-derived tumours. Low HIF1α expression (right panels) favoured intense expression of HIF2α (left panels).
The different involvement of HIFs was also confirmed by histological analysis of subcutaneous tumours as previously described. As reported in Figure 2C, in both 786-O and Caki-1 sphere-derived tumours, we observed an absent or low expression of HIF1α (right panels) in favour of the intense expression of HIF2α (left panels).
HIF2α is important for tumour formation
We used a stable knockdown approach to further investigate the role of HIF2α in promoting the sphere-forming capacity of CSCs-like RCC cells. The transfection of 786-O cells with a plasmid containing a short hairpin RNA for HIF2α (sh-HIF2α) abrogated the expression of HIF2α as analysed by western blotting (Figure 3A). However, sh-HIF2α-transfected 786-O cells showed a progressive reduction in the number of sphere initiating cells in serially passaged cultures (Figure 3B) without altering their proliferation in adherent conditions (data not shown). When we injected 3 × 106 786-O cells transfected with sh-HIF2α into NOD/SCID mice, we revealed a marked decrease in tumour volume compared with the masses from 786-O cells transfected with the control sh-Empty vector. These results are shown in a qualitative and quantitative representation in Figure 3C and Table 2, respectively. Moreover, no tumours occurred when 5 × 104 sphere-derived cells transfected with sh-HIF2α were injected into mice (Table 2). Overall, these findings suggest that HIF2α increases cellular plasticity and shifts the dynamic equilibrium in cancer cells with non-stem properties towards stem cell properties. Moreover, the inhibition of HIF2α may have a great impact on cancer therapy.

HIF2 α is important for tumour formation. (A) Western blot showing a nearly complete reduction of HIF2α expression in 786-O cells transfected with sh-HIF2α vector. (B) Qualitative and quantitative representation of the reduction in the number of spheres in 786-O sh-HIF2α-transfected cells compared with controls. *P<0.0026. (C) Image of a xenograft transplant of 3 × 106 cells transfected with 786-O sh-HIF2α and sh-Empty vectors.
HIF2α pathways interact with CXCR4 signalling in human RCC
To further understand the role that HIF2α had in the CSCs phenotype maintenance and its possible interaction with CXCR4 signalling, we examined the effects of the stable knockdown of HIF2α on CXCR4 expression. We found that HIF2α stable knockdown in 786-O cells specifically impaired CXCR4 and VEGF mRNA expression by at least 50% compared with the control cells (Figure 4A). Consistently, the protein level of CXCR4 was strongly reduced during HIF2α inhibition (Figure 4B), suggesting that the HIF2α signalling pathway promotes the expansion of CXCR4-positive RCC cells with CSCs properties. To substantiate these observations in human settings, we investigated HIF2α expression and its correlation with CXCR4 taking advantage of the NextBio bioinformatics tool. We used the ‘Disease Atlas’ section to value the gene expression analysis of human renal tumours. Gene analysis revealed a strong connection of HIF2α expression and RCC subtypes (Figure 4C). An apparent increase in the expression of CXCR4 gene was also observed while only 3 out of 20 of clear cell RCC that express HIF2α, showed a downregulation of CXCR4 (Figure 4D). These observations suggest that HIF2α may be associated with CXCR4 in human clear cell RCC.

The HIF2 α pathways interact with CXCR4 signalling. (A) Real-time PCR of CXCR4, NANOG, and OCT4 genes in 786-O cells transfected with sh-HIF2α vector and with the control vector (sh-Empty). There was a strong reduction in CXCR4 and VEGF expression in 786-O sh-HIF2α cells. *P<0.0001, **P<0.0012. (B) Western blot showing a strong reduction in the CXCR4 protein level in 786-O sh-HIF2α cells. (C) Analysis of HIF2α expression in human clear cell RCC (CC-RCC) tumours vs other tumour subtypes and vs normal kidney tissue (NextBio bioinformatics tool). (D) Correlation of CXCR4-RCC expressing tumours with CC-RCC tumours expressing the HIF2α gene (NextBio bioinformatics tool).
Discussion
Cancer cells with CSCs-like properties are known to drive tumour progression and metastasis. Consistently, the number of marker-positive stem cells identified by immunohistology was found to correlate with a poorer prognosis in cancer patients (Li et al, 2010; Su et al, 2010; Zhang et al, 2012; Nakata et al, 2013). CSCs have also been identified and isolated from RCC patients and cell lines using several putative stem cell markers such as CD105 (endoglin), the intracellular aldehyde dehydrogenase 1 (ALDH1) activity, or the chemokine receptor CXCR4 (Bussolati et al, 2008; Gassenmaier et al, 2013; Ueda et al, 2013). On the basis of these different molecules, which are expressed on the cell surface, CSCs can be involved in specific mechanisms of tumorigenesis. Indeed, CSCs expressing CD105 appear to be important in tumour angiogenesis (Nassiri et al, 2011). Cancer cells with high ALDH1 activity have the great potential to function as CSCs in metastatic RCC cell lines (Ueda et al, 2013). Moreover, RCC cell lines derived from the metastatic site were found to be enriched in CXCR4-positive cells, and these cells showed a higher sphere-forming ability, self-renewal capacity and tumorigenicity (Gassenmaier et al, 2013). Surgical biopsies of RCC patients confirmed that high expression of CXCR4 has a significant prognostic value and therapeutic importance (D'Alterio et al, 2010). However, CXCR4 gene expression did not separate the three most common RCC histologic subtypes: clear cell, chromophobe, and papillary RCC (Cairns, 2010). Therefore, it is important to identify in each RCC subtype the deregulated molecular pathways that are involved in the generation of CXCR4-positive cells.
The most common RCC histological type is clear cell carcinoma, also called conventional RCC, which represents 75%–80% of RCC. LOH of 3p and point mutation of VHL are frequent and early in clear cell RCC (Cairns, 2010). Biallelic VHL inactivation leads to an increased abundance of HIF1α and HIF2α, and compelling evidence suggests that HIF2α, rather than HIF1α, promotes pVHL-deficient RCC tumorigenesis (Baldewijns et al, 2010)
The major goal of our study was to identify RCC cancer cells with different sphere-forming abilities that were preferentially altered in pVHL, HIF1α, or HIF2α. This analysis gave us a unique opportunity to probe the novel molecular pathways involved in the generation of cancer cells with CSCs-like properties. Here, we demonstrated that HIF2α expression was elevated in sphere-derived cells, and we propose that HIF2α-dependent signalling preferentially promoted a CSC-like phenotype involved in tumour initiation and progression of conventional RCCs.
HIF1α and HIF2α have 48% amino acid sequence identity and similar protein structures but are non-redundant in function because they have distinct target genes and mechanisms of regulation (Koh and Powis, 2012). Intriguingly, it appears that in some cell lines, HIF1α is most active during short periods (2–24 h) of intense hypoxia or anoxia (<0.1% O2), whereas HIF2α is active under mild or physiological hypoxia (<5% O2) and continues to remain active even after 48–72 h of hypoxia (Holmquist-Mengelbier et al, 2006). Hence, in certain contexts, HIF1α drives the initial response to hypoxia, but during chronic hypoxic exposure, it is HIF2α that has the major role in driving the hypoxic response (Holmquist-Mengelbier et al, 2006; Koh et al, 2011). This HIF ‘switch’ results in HIF1α and HIF2α playing divergent but complementary roles under both physiological and pathophysiological conditions (Koh and Powis, 2012). Our results strongly indicate that HIF2α, but not HIF1α, generates RCC cells with CSC-like properties.
It is unclear how RCC cells produce HIF2α and acquire a CSC-like phenotype during sphere culture. At the present time, we can only speculate about HIF2α activation in RCC cells and that the activation of HIF2α in CSC-like phenotype is driven by a non-canonical signalling. One possibility is the existence of a link between epidermal growth factor receptor (EGFR) signalling and oxygen-independent HIF2α upregulation in RCC cells. Previous studies have shown that the EGF receptor signalling pathway is usually deregulated in cancer and is suggested to have an important role in RCC (Dias et al, 2013). During sphere culture, EGF stimulates HIF2α protein expression (our unpublished data); this may be achieved by the phosphoinositide 3-kinase/AKT pathway. Obviously, this possibility needs to be examined to better understand the mechanism of the conversion of non-CSCs into CSCs.
To characterise RCC-derived CSCs, we analysed seven putative CSC markers, and CXCR4 was found to be the most expressed. Previous studies in colon cancer cells showed that CXCR4 expression is regulated by hypoxia and HIFs (Romain et al, 2014), but a direct connection among HIF2α, CXCR4 signalling, and RCC-derived CSCs was previously lacking. Significantly, after HIF2α knockdown, CXCR4 expression and enrichment in the CSC population were almost entirely lost. This emphasises that co-activation of both HIF2α and CXCR4 leads to the acquisition of a CSC phenotype. Given that poor clinical prognosis was associated with elevated CXCR4 expression in RCC, it is possible that tumour cells with high levels of CXCR4 expression and activation of HIF2α are more resistant to conventional therapies that lead to more prevalent rates of relapse. Hence, understanding how CSCs are regulated by HIF2α and CXCR4 activation is crucial if they are to be targeted for therapy.
Change history
20 October 2015
This paper was modified 12 months after initial publication to switch to Creative Commons licence terms, as noted at publication
References
Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF (2003) Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci USA 100: 3983–3988.
Bussolati B, Bruno S, Grange C, Ferrando U, Camussi G (2008) Identification of a tumor-initiating stem cell population in human renal carcinomas. FASEB J 22: 3696–3705.
Baldewijns MM, van Vlodrop IJ, Vermeulen PB, Soetekouw PM, van Engeland M, de Bruïne AP (2010) VHL and HIF signaling in renal cell carcinogenesis. J Pathol 221: 125–138.
Cairns P (2010) Renal cell carcinoma. Cancer Biomark 9: 461–473.
Clarke MF, Dick JE, Dirks PB, Eaves CJ, Jamieson CH, Jones DL, Visvader J, Weissman IL, Wahl GM (2006) Cancer stem cells—perspectives on current status and future directions: AACR Workshop on cancer stem cells. Cancer Res 66: 9339–9344.
Chiavarina B, Martinez-Outschoorn UE, Whitaker-Menezes D, Howell A, Tanowitz HB, Pestell RG, Sotgia F, Lisanti MP (2012) Metabolic reprogramming and two-compartment tumor metabolism: opposing role(s) of HIF1α and HIF2α in tumor-associated fibroblasts and human breast cancer cells. Cell Cycle 11: 3280–3289.
Covello KL, Kehler J, Yu H, Gordan JD, Arsham AM, Hu CJ, Labosky PA, Simon MC, Keith B (2006) HIF-2alpha regulates Oct-4: effects of hypoxia on stem cell function, embryonic development, and tumor growth. Genes Dev 20: 557–570.
D'Alterio C, Cindolo L, Portella L, Polimeno M, Consales C, Riccio A, Cioffi M, Franco R, Chiodini P, Cartenì G, Mirone V, Longo N, Marra L, Perdonà S, Claudio L, Mascolo M, Staibano S, Falsaperla M, Puglisi M, Martignoni G, Ficarra V, Castello G, Scala S (2010) Differential role of CD133 and CXCR4 in renal cell carcinoma. Cell Cycle 9: 4492–4500.
Dias F, Teixeira AL, Santos JI, Gomes M, Nogueira A, Assis J, Medeiros R (2013) Renal cell carcinoma development and miRNAs: a possible link to the EGFR pathway. Pharmacogenomics 14: 1793–1803.
Gassenmaier M, Chen D, Buchner A, Henkel L, Schiemann M, Mack B, Schendel DJ, Zimmermann W, Pohla H (2013) CXC chemokine receptor 4 is essential for maintenance of renal cell carcinoma-initiating cells and predicts metastasis. Stem Cells 31: 1467–1476.
Gelmini S, Mangoni M, Serio M, Romagnani P, Lazzeri E (2008) The critical role of SDF-1/CXCR4 axis in cancer and cancer stem cells metastasis. J Endocrinol Invest 31: 809–819.
Heddleston JM, Li Z, McLendon RE, Hjelmeland AB, Rich JN (2009) The hypoxic microenvironment maintains glioblastoma stem cells and promotes reprogramming towards a cancer stem cell phenotype. Cell Cycle 8: 1–11.
Holmquist-Mengelbier L, Fredlund E, Löfstedt T, Noguera R, Navarro S, Nilsson H, Pietras A, Vallon-Christersson J, Borg A, Gradin K, Poellinger L, Påhlman S (2006) Recruitment of HIF-1alpha and HIF 2alpha to common target genes is differentially regulated in neuroblastoma:HIF-2alpha promotes an aggressive phenotype. Cancer Cell 10: 413–423.
Kaelin WG Jr (2008) The von Hippel-Lindau tumour suppressor protein: O2 sensing and cancer. Nat Rev Cancer 8: 865–873.
Kaur S, Singh G, Kaur K (2014) Cancer stem cells: an insight and future perspective. J Cancer Res Ther 10: 846–852.
Keith B, Simon MC (2007) Hypoxia-inducible factors, stem cells, and cancer. Cell 129: 465–472.
Keith B, Johnson RS, Simon MC (2012) HIF1α and HIF2α: sibling rivalry in hypoxic tumour growth and progression. Nat Rev Cancer 12: 9–22.
Khan MI, Czarnecka AM, Duchnowska R, Kukwa W, Szczylik C (2013) Metastasis-initiating cells in renal cancer. Curr Signal Transduct Ther 8: 240–246.
Koh MY, Lemos R Jr, Liu X, Powis G (2011) The hypoxia-associated factor switches cells from HIF-1α to HIF-2α dependent signaling promoting stem cell characteristics, aggressive tumor growth and invasion. Cancer Res 71: 4015–4027.
Koh MY, Powis G (2012) Passing the baton: the HIF switch. Trends Biochem Sci 37: 364–372.
Li T, Su Y, Mei Y, Leng Q, Leng B, Liu Z, Stass SA, Jianq F (2010) ALDH1A1 is a marker for malignant prostate stem cells and predictor of prostate cancer patients’ outcome. Lab Invest 90: 234–244.
Ljungberg B (2004) Prognostic factors in renal cell carcinoma. Urologe 43 (Suppl 3): S119–S120.
McLaughlin JK, Lipworth L (2000) Epidemiologic aspects of renal cell cancer. Semin Oncol 27: 115–123.
Nakata S, Campos B, Bageritz J, Bermejo JL, Becker N, Engel F, Acker T, Momma S, Herold Mende C, Lichter P, Radlwimmer B, Goidts V (2013) LGR5 is a marker of poor prognosis in glioblastoma and is required for survival of brain cancer stem-like cells. Brain Pathol 23: 60–72.
Nassiri F, Cusimano MD, Scheithauer BW, Rotondo F, Fazio A, Yousef GM, Syro LV, Kovacs K, Lloyd RV (2011) Endoglin (CD105): a review of its role in angiogenesis and tumor diagnosis,progression and therapy. Anticancer Res 31: 2283–2290.
Pan J, Zhang Q, Wang Y, You M (2010) 26S proteasome activity is down-regulated in lung cancer stem-like cells propagated in vitro. PLoS One 5: e13298.
Reya T, Morrison SJ, Clarke MF, Weissman IL (2001) Stem cells, cancer, and cancer stem cells. Nature 414: 105–111.
Ricci-Vitiani L, Lombardi DG, Pilozzi E, Biffoni M, Todaro M, Peschle C, De Maria R (2007) Identification and expansion of human colon-cancer-initiating cells. Nature 445: 111–115.
Romain B, Hachet-Haas M, Rohr S, Brigand C, Galzi JL, Gaub MP, Pencreach E, Guenot D (2014) Hypoxia differentially regulated CXCR4 and CXCR7 signaling in colon cancer. Mol Cancer 13: 58.
Shinojima T, Oya M, Takayanagi A, Mizuno R, Shimizu N, Murai M (2007) Renal cancer cells lacking hypoxia inducible factor (HIF)-1alpha expression maintain vascular endothelial growth factor expression through HIF-2alpha. Carcinogenesis 28: 529–536.
Siegel R, Naishadham D, Jemal A (2012) Cancer statistics, 2012. CA Cancer J Clin 62: 10–29.
Singh SK, Clarke ID, Terasaki M, Bonn VE, Hawkins C, Squire J, Dirks PB (2003) Identification of a cancer stem cell in human brain tumors. Cancer Res 63: 5821–5828.
Soeda A, Inagaki A, Oka N, Ikegame Y, Aoki H, Yoshimura S, Nakashima S, Kunisada T, Iwama T (2008) Epidermal growth factor plays a crucial role in mitogenic regulation of human brain tumor stem cells. J Biol Chem 283: 958–966.
Su Y, Qiu Q, Zhang X, Jiang Z, Leng Q, Liu Z, Stass SA, Jiang F (2010) Aldehyde dehydrogenase 1 A1-positive cell population is enriched in tumor-initiating cells and associated with progression of bladder cancer. Cancer Epidemiol Biomarkers Prev 19: 327–337.
Ueda K, Ogasawara S, Akiba J, Nakayama M, Todoroki K, Ueda K, Sanada S, Suekane S, Noguchi M, Matsuoka K, Yano H (2013) Aldehyde dehydrogenase 1 identifies cells with cancer stem cell-like properties in a human renal cell carcinoma cell line. PLoS One 8: e75463.
Vermeulen L, de Sousa e Melo F, Richel DJ, Medema JP (2012) The developing cancer stem-cell model: clinical challenges and opportunities. Lancet Oncol 13: e83–e89.
Zagzag D, Krishnamachary B, Yee H, Okuyama H, Chiriboga L, Ali MA, Melamed J, Semenza GL (2005) Stromal cell-derived factor-1alpha and CXCR4 expression in hemangioblastoma and clear cell-renal cell carcinoma von Hippel-Lindau loss-of-function induces expression of a ligand and its receptor. Cancer Res 65: 6178–6188.
Zhang SS, Han ZP, Jing YY, Tao SF, Li TJ, Wang H, Wang Y, Li R, Yang Y, Zhao X, Xu XD, Yu ED, Rui YC, Liu HJ, Zhang L, Wei LX (2012) CD133+CXCR4+ colon cancer cells exhibit metastatic potential and predict poor prognosis of patients. BMC Med 10: 85.
Zhong Y, Guan K, Guo S, Zhou C, Wang D, Ma W, Zhang Y, Li C, Zhang S (2010) Sphere derived from the human SK-RC-42 renal cell carcinoma cell line are enriched in cancer stem cells. Cancer Lett 299: 150–160.
Zhou BB, Zhang H, Damelin M, Geles KG, Grindley JC, Dirks PB (2009) Tumour-initiating cells: challenges and opportunities for anticancer drug discovery. Nat Rev Drug Discov 8: 8.
Acknowledgements
We are grateful to Daniele Giannini (Università Politecnica Marche) for his technical support and Most Mauluda Akhtar for her advice on clearly presenting the manuscript. This work was supported by the Italian Ministry of Research (PRIN grant) and the RSA (ex 60%) grant. C Micucci was supported by a fellowship from the University of Marche. We are grateful to the staff of the Experimental Pathology Unit at the Department of Clinical and Molecular Sciences, School of Medicine, University of Marche.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
This work is published under the standard license to publish agreement. After 12 months the work will become freely available and the license terms will switch to a Creative Commons Attribution-NonCommercial-Share Alike 4.0 Unported License.
Supplementary Information accompanies this paper on British Journal of Cancer website
Rights and permissions
From twelve months after its original publication, this work is licensed under the Creative Commons Attribution-NonCommercial-Share Alike 4.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Micucci, C., Matacchione, G., Valli, D. et al. HIF2α is involved in the expansion of CXCR4-positive cancer stem-like cells in renal cell carcinoma. Br J Cancer 113, 1178–1185 (2015). https://doi.org/10.1038/bjc.2015.338
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/bjc.2015.338
Keywords
This article is cited by
-
Hypoxia, oxidative stress, and the interplay of HIFs and NRF2 signaling in cancer
Experimental & Molecular Medicine (2024)
-
ISCA2 inhibition decreases HIF and induces ferroptosis in clear cell renal carcinoma
Oncogene (2022)
-
Hypoxia inducible factor signaling in breast tumors controls spontaneous tumor dissemination in a site-specific manner
Communications Biology (2021)
-
Follistatin-like protein 1 plays a tumor suppressor role in clear-cell renal cell carcinoma
Chinese Journal of Cancer (2018)
-
Co-expression of Cancer Stem Cell Markers OCT4 and NANOG Predicts Poor Prognosis in Renal Cell Carcinomas
Scientific Reports (2018)

{kind=link}
{kind=link}
{kind=link}