
An intricate vocal anatomy underlies the subtleties of this animal's melodic mating calls.
Abstract
Animals' sound-producing organs often act as an integrated whole — particular vocal structure are not directly associated with the creation of discrete syllables1,2,3,4. But here we show that the ‘chuck’ of the ‘whine-chuck’ mating call of the túngara frog, Physalaemus pustulosus, is caused by a fibrous mass attached to the vocal folds; the chuck is eliminated by removal of this structure, although the frog still tries to produce the sound. Sexual selection affects the acoustic complexity of the frog's call5,6,7, so evolution may have shaped this unusual vocalization, which is akin to the two-voiced song of songbirds6,8.
Similar content being viewed by others


Main
Calls of túngara frogs consist of a whine followed by up to seven chucks. The whine is a long-frequency sweep, whereas the chuck is a shorter sound whose fundamental frequency is half that of the whine (for details, see supplementary information). Females prefer a whine–chuck call to a whine alone (probability of 0.856; 3,135 versus 527; exact binomial, P < 0.000001).
In P. pustulosus and some close relatives, the vocal folds support a fibrous mass that extends into the bronchus (see supplementary information). The size of this mass correlates with the presence or absence of chuck-like call components among species within the genus1, within the P. pustulosus species group9 and between populations of the túngara frog's sister species, P. petersi10.
To investigate whether the fibrous mass could be responsible for production of the chuck sound, we recorded calls of male túngara frogs in the field (‘pre-treatment’ condition). We then anaesthetized the frogs, surgically removed the posterior elongation of the fibrous mass, and recorded their calls after they had recovered (‘post-treatment’ condition). A sham control replicated all aspects of the surgery apart from ablation of the mass. An additional control omitted surgical manipulation but replicated the feeding, housing and handling used in the other treatments.
As the fundamental frequency of the chuck is half that of the whine, its odd harmonics do not coincide with the whine's harmonics, although its even harmonics do (see supplementary information). We predicted that ablation of the fibrous mass would reduce the amount of energy in odd relative to even harmonics of the chuck, but would not influence the spectral features of the whine or the increased amplitude at the call's end.
The relative energy in odd versus even harmonics of the chuck was significantly reduced after surgical treatment (F2,17 = 21.77, P < 0.001; Fig. 1a, b). The harmonic-energy ratio was significantly lower than in either of the two controls (Tukey post-hoc test; all comparisons, P < 0.001); the two controls did not differ from one another (Tukey test, P = 0.538). There was no treatment effect in the same analysis of the whine (F2,17 = 0.891, P = 0.429).

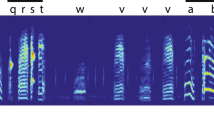
a, Relative amplitude of odd (red bars) to even (blue) harmonics in each surgical-treatment group for ‘chuck’ (left panel) and ‘whine’ (right panel) call components; means and standard errors are shown. Surgery, n = 8; sham, n = 6; none, n = 6. b–d, Representative calls (top, relative amplitude of waveforms; bottom, spectrogram) by an individual male in pre- (left panels) and post-treatment (right panels) conditions after b, ablation of the ‘chuck’-associated fibrous mass; c, sham surgery; and d, no surgery.
Although the chuck is diagnosed by its fundamental frequency relative to that of the whine, it also increases the amplitude of the end of the call. An increase in amplitude is therefore evidence that the male is attempting to produce a chuck. As predicted, there was no treatment effect on the relative amplitude at the end of the call (F2,21 = 0.287, P = 0.754; Fig. 1c, d). Males without the posterior elongation of the fibrous mass therefore still increase call amplitude at the end of the whine, as they would when producing a chuck, but the spectral structure of the chuck is missing. Simply put, males without an elongated fibrous mass on their vocal folds try to produce chuck sounds but are unable to do so.
References
Drewry, G., Heyer, W. R. & Rand, A. S. Copeia 1982, 636–645 (1982).
Suthers, R. A. Nature 347, 473–477 (1990).
Fee, M. S., Shraiman, B., Pesaran, B. & Mitra, P. P. Nature 395, 67–71 (1998).
Fitch, W. T. Anim. Behav. 63, 407–428 (2002).
Ryan, M. J. Science 209, 523–525 (1980).
Ryan, M. J. The Túngara Frog: A Study in Sexual Selection and Communication (Univ. Chicago Press, Chicago, 1985).
Ryan, M. J., Fox, J. H., Wilczynski, W. & Rand, A. S. Nature 343, 66–67 (1990).
Greenwalt, C. H. Bird Song: Acoustics and Physiology (Smithsonian Institution Press, Washington DC, 1968).
Ryan, M. J. & Drewes, R. C. Biol. J. Linn. Soc. 40, 37–52 (1990).
Boul, K. E. & Ryan, M. J. Copeia 2004, 624–631 (2004).
Author information
Authors and Affiliations
Corresponding author
Supplementary information
Supplementary information
(DOC 107 kb)
Supplementary audio 1
Advertisement call of a túngara frog recorded in the wild before surgical removal of the laryngeal fibrous masses. The second component of the call is half the fundamental frequency of the first component. (WAV 51 kb)
Supplementary audio 2
Advertisement call of the same túngara frog after surgical removal of the laryngeal fibrous masses. The odd harmonics of the second component of the call are missing. (WAV 51 kb)
Rights and permissions
About this article
Cite this article
Gridi-Papp, M., Rand, A. & Ryan, M. Complex call production in the túngara frog. Nature 441, 38 (2006). https://doi.org/10.1038/441038a
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/441038a
This article is cited by
-
Are frog calls relatively difficult to locate by mammalian predators?
Journal of Comparative Physiology A (2023)
-
Conspecific disturbance odors act as alarm cues to affect female mate choice in a treefrog
Behavioral Ecology and Sociobiology (2022)
-
Epigenomic changes in the túngara frog (Physalaemus pustulosus): possible effects of introduced fungal pathogen and urbanization
Evolutionary Ecology (2019)
-
Adaptive changes in sexual signalling in response to urbanization
Nature Ecology & Evolution (2018)
-
Environmental conditions limit attractiveness of a complex sexual signal in the túngara frog
Nature Communications (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
