
Abstract
Birds can perceive the reflectance of ultraviolet light by biological structures1,2,3,4,5. Here we show that the skin of the mouth and body of starling nestlings substantially reflects light in the ultraviolet range and that young in which this reflectance is reduced will gain less mass than controls, despite low background levels of ultraviolet and visible light in the nest. We suggest that this ultraviolet reflectance from starling nestlings and its contrast with surrounding surfaces are important for parental decisions about food allocation.
Similar content being viewed by others


Main
Reflectance of ultraviolet light in the 300–400-nanometre range from feathers, fruits, insects and vole scent marks is used by birds to make decisions about mate choice and food selection1,2,3,4,5. The function of ultraviolet reflectance by the brightly coloured skin of some birds is unknown6,7, but it may play a role in parent–offspring communication6.
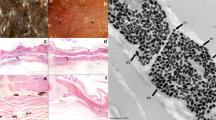
The flanges (skin surrounding the mouth) in nestlings of both starlings (Sturnus vulgaris) and great tits (Parus major) is strongly reflective (Fig. 1a,c, respectively; for methods, see supplementary information). In addition, the body skin of starling nestlings from two separate populations (in Switzerland and Britain; see supplementary information) reflects in the ultraviolet (Fig. 1b,e), although the body skin of great-tit (Fig. 1d) and blackbird (Turdus merula) nestlings does not6.

a–d, Reflectance spectra from nestling skin. Starlings: mouth flanges (a), n = 10; body skin (b), n = 10. Great tits: mouth flanges (c), n = 25; body skin (d), n = 22. Blue, untreated skin; red, skin treated with petroleum jelly containing ultraviolet-light blocker; green, skin treated with petroleum jelly without ultraviolet blocker; curves show medians ± s.d. e, Nest irradiance (black curve; n = 5) and reflectance spectra from mouth flanges (orange; n = 10) and body skin (purple; n = 6) of starling nestlings; median curves are shown.
We investigated the function of this skin reflectance in starlings by applying ultraviolet-light blockers8 on the nestlings' bodies and flanges and measuring their body-mass gain after two hours9. Nestlings with ultraviolet-reflecting skin (‘+UV’) gained relatively more mass than non-reflecting (‘−UV’) nestlings (difference in mass gain ± s.e.: +UV, 0.23 ± 0.14 g and −UV, −0.24 ± 0.12 g; F1,27 = 4.84, P = 0.036; when included as a random factor in the model (nestrandom), the nest effect was not significant, F11,27 = 0.30). In most broods, the mean mass gained by +UV nestlings was greater than that gained by −UV nestlings (sign test, P = 0.038). By contrast, we found that modifying ultraviolet reflectance on only the mouth flanges in both starling and great tit nestlings did not lead to significant differences in mass gain (starlings: F1,101 = 0.38, P = 0.54, nestrandom was not significant; great tits: F2,330 = 0.142, P = 0.87, nestrandom was not significant).
Our results indicate that artificial reduction of the ultraviolet reflectance of body skin and flanges affects mass gain by nestlings. The brightness of skin reflectance in ultraviolet and visible light by starling nestlings did not correlate with either their body mass or tarsus length, but showed a positive relation with their cell-mediated immune responses (see supplementary information; F1,13 = 7.99, P = 0.014; mean brood values: F1,6 = 20.96, P = 0.006). The brightness of nestling skin reflectance in these birds might therefore signal their phenotypic quality and/or condition6,10.
An alternative explanation could be that nestlings that do not reflect in the ultraviolet might be less easily detected by their parents in dark nests (Fig. 1e) than those that do9. We found that ultraviolet-blocking material on the body skin spread after two hours to other young in the brood, reducing the overall body-skin ultraviolet reflectance of the brood. This reduction in ultraviolet contrast between the chicks and their nest, in association with differences in flange ultraviolet reflectance, could lead to less efficient chick detection by the parents, who rely on achromatic or chromatic mechanisms to detect them. This might explain the observed body-mass differences.
The phylogenetic patterns in the evolution of nanostructures7 and the pigments responsible for ultraviolet reflectance by skin still need to be determined. Given the likely function of skin reflectance in parent–offspring communication, the ecological factors that affect skin reflectance and the associated parental responses should be investigated.
References
Cuthill, I. C. et al. Adv. Study Behav. 29, 159–214 (2000).
Bennett, A. T. D., Cuthill, I. C., Partridge, J. C. & Maier, E. J. Nature 380, 433–435 (1996).
Bennett, A. T. D., Cuthill, I. C. & Norris, K. J. Am. Nat. 144, 848–860 (1994).
Sheldon, B. C., Andersson, S., Griffiths, S. C., Örnborg, J. & Sendecka, J. Nature 402, 874–877 (1999).
Viitala, J., Korpimäki, E., Palokangas, P. & Koivula, M. Nature 373, 425–427 (1995).
Hunt, S., Kilner, R. M., Langmore, N. E. & Bennett, A. T. D. Proc. R. Soc. Lond. 270, (suppl.) 25–28 (2003).
Prum, R. O. & Torres, R. J. Exp. Biol. 206, 2409–2429 (2003).
Andersson, S. & Amundsen, T. Proc. R. Soc. Lond. B 264, 1587–1591 (1997).
Heeb, P., Schwander, T. & Faoro, S. Anim. Behav. 66, 637–642 (2003).
Kilner, R. M. Proc. R. Soc. Lond. B 264, 963–968 (1997).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
About this article
Cite this article
Jourdie, V., Moureau, B., Bennett, A. et al. Ultraviolet reflectance by the skin of nestlings. Nature 431, 262 (2004). https://doi.org/10.1038/431262a
Published:
Issue Date:
DOI: https://doi.org/10.1038/431262a
This article is cited by
-
Experimental reduction of a nest ectoparasite affects mouth coloration of nestling Cliff Swallows Petrochelidon pyrrhonota
Journal of Ornithology (2022)
-
The early maternal environment shapes the parental response to offspring UV ornamentation
Scientific Reports (2021)
-
Exposure to UV radiance predicts repeated evolution of concealed black skin in birds
Nature Communications (2020)
-
Skin and flange colour, but not ectoparasites, predict condition and survival in starling nestlings
Behavioral Ecology and Sociobiology (2017)
-
Blocking of ultraviolet reflectance on bird eggs reduces nest predation by aerial predators
Journal of Ornithology (2016)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
