
Abstract
Ampakines act as positive allosteric modulators of AMPA-type glutamate receptors and facilitate hippocampal long-term potentiation (LTP), a mechanism associated with memory storage and consolidation. The present study investigated the acute effects of farampator, 1-(benzofurazan-5-ylcarbonyl) piperidine, on memory and information processes in healthy elderly volunteers. A double-blind, placebo-controlled, randomized, cross-over study was performed in 16 healthy, elderly volunteers (eight male, eight female; mean age 66.1, SD 4.5 years). All subjects received farampator (500 mg) and placebo. Testing took place 1 h after drug intake, which was around Tmax for farampator. Subjects performed tasks assessing episodic memory (wordlist learning and picture memory), working and short-term memory (N-back, symbol recall) and motor learning (maze task, pursuit rotor). Information processing was assessed with a tangled lines task, the symbol digit substitution test (SDST) and the continuous trail making test (CTMT). Farampator (500 mg) unequivocally improved short-term memory but appeared to impair episodic memory. Furthermore, it tended to decrease the number of switching errors in the CTMT. Drug-induced side effects (SEs) included headache, somnolence and nausea. Subjects with SEs had significantly higher plasma levels of farampator than subjects without SEs. Additional analyses revealed that in the farampator condition the group without SEs showed a significantly superior memory performance relative to the group with SEs. The positive results on short-term memory and the favorable trends in the trail making test (CTMT) are interesting in view of the development of ampakines in the treatment of Alzheimer's disease and schizophrenia.
Similar content being viewed by others
INTRODUCTION
Ampakines act as positive allosteric modulators of AMPA-type glutamate receptors (Arai et al, 1994; Yamada, 1998). In animals the drugs have been shown to facilitate hippocampal long-term potentiation (LTP), a mechanism associated with memory storage and formation (Lynch, 1998; Staubli et al, 1994a, 1994b).
Animal experiments have also demonstrated that ampakines improve performance in a variety of memory tasks like odor discrimination (Larson et al, 1995; Staubli et al, 1994a), spatial mazes (Granger et al, 1993; Staubli et al, 1994a), a spatial delayed-non-match-to-sample task (Hampson et al, 1998), eye-blink conditioning (Shors et al, 1995) and learned fear (Lebrun et al, 2000). Ampakines have also been shown to be effective in reducing age-associated memory deficits in rats (Granger et al, 1996). Furthermore, ampakines appear to be involved in other processes besides memory. Larson et al (1996) showed that the ampakine BPD-29 suppressed methamphetamine-induced stereotyped behavior. Next, Johnson et al (1999) found that the effects of very small doses of antipsychotics were greatly enhanced by the ampakine CX516. These findings led to the development of ampakines for the treatment of both Alzheimer's Disease (AD) and schizophrenia.
The current knowledge about the effects of ampakines on human memory and other cognitive processes is, however, still rather limited. In healthy young adults, 300 mg CX516 improved the picture association, recognition of scent sequence and visuospatial maze performance although it only tended to enhance the recall of playing cards (Ingvar et al, 1997). In another study with young volunteers 600–1200 mg CX516 was found to augment the delayed recall of nonsense syllables relative to baseline, whereas doses of 75–300 mg CX516 and placebo did not enhance performance (Lynch et al, 1996). In healthy elderly subjects (age range 65–75 years) 900 mg CX516 caused a twofold increase in the recall of nonsense syllables (Lynch et al, 1997, 2003). In neither studies did Lynch (2004) find any changes in tests assessing psychomotor performance, heart rate or mood.
To date, only two small-scale studies have been published that investigated ampakines in patients with schizophrenia. One was a double-blind placebo-controlled trial in which CX516 was added to clozapine treatment for 4 weeks ((19 patients in total: 12 were allocated to CX516 treatment and seven to the placebo treatment group) Goff et al, 2001). The ampakine group showed moderate to large improvements compared to the placebo group on attention and memory performance and showed a reduction of negative symptoms. It should be noted, however, that the ampakine group was substantially more impaired at baseline than the placebo group and that the small sample size prohibited any statistical analyses. In the second study, Marenco et al (2002) administered CX516 to four patients with schizophrenia as a stand-alone antipsychotic treatment (in one patient as a single-blind treatment and in three patients as a double-blind treatment) for 2–4 weeks; two of the four patients receiving placebo dropped out. The study failed to yield any noticeable improvements in psychotic symptoms or cognitive processes.
The present study is an exploratory phase I study and looks into the acute effects of farampator (1-(benzofurazan-5-ylcarbonyl) piperidine), a specific positive allosteric modulator of the alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA)-type glutamate receptors. This so-called ampakine is chemically distinct from CX516 and is being developed for the treatment of schizophrenia with emphasis on the improvement of negative symptoms and cognitive dysfunction. The basis for this lies in the glutamate balance theory for schizophrenia in which the potentiation of glutamatergic transmission is seen as having a potentially antipsychotic effect (Johnson et al, 1999). Evidence for glutaminergic involvement in schizophrenia comes from imaging and post-mortem studies that have found abnormalities in brain regions that are connected by glutaminergic circuits. Reduced excitatory (glutamatergic) activity might underlie some of the symptoms. This is substantiated by the finding that drugs like ketamine and phencyclidine, antagonists of NDMA type glutamate receptors exacerbate symptoms in patients and produce psychotic symptoms in healthy individuals. Drugs that enhance glutamatergic transmission on the other hand, might correct the imbalance in this system in the schizophrenic patients.
AMPA receptors are the most abundant glutamate receptors in the brain and mediate fast excitatory neurotransmission as well as activation of co-localized NMDA receptors through partial depolarization of the postsynaptic membrane. Enhancement of AMPA receptor activity is anticipated to upregulate glutamatergic function through both AMPA and NMDA receptors. Using whole-cell patch-clamp recordings in rat cultured hippocampal neurons; farampator has been shown to increase glutamate-induced steady state currents with an EC50 value of 14 μM compared to 95 μM for the ampakine CX516 (unpublished findings). Farampator was also more potent than CX516 in increasing the slope of field excitatory postsynaptic potentials (fEPSPs) in rat hippocampal slices. The effects of farampator were blocked by NBQX. These preliminary results indicate that farampator is an allosteric modulators of AMPA receptor function in hippocampal neurones and that it is as such more potent than CX516.
Farampator, at micromolar concentrations, potentiates AMPA receptor-mediated responses in a number of in vitro assay systems and enhances hippocampal long-term potentiation (M Shahid et al, unpublished data), suggestive that farampator may, apart from antipsychotic properties, also have cognitive enhancing effects.
As farampator is developed primarily as a putative antipsychotic, and since a large number of cognitive domains can be disturbed in schizophrenia (Green et al, 2004; Heinrichs and Zakzanis, 1998; Nuechterlein et al, 2004), the present study was designed to look into the effects of the drug on tests of memory, as well as information processing (with components of speed of processing, visuospatial processing, and set-shifting) in healthy elderly volunteers.
MATERIALS AND METHODS
Participants
The study was approved by the local medical ethics committee and has been carried out in accordance with the Declaration of Helsinki. Prior to the trial, all participants gave their written informed consent.
Enhancing cognitive performance in young healthy volunteers is rather difficult because they perform close to their optimum (Foster et al, 1998). To circumvent this ceiling effect, we decided to choose elderly subjects, in which an age-related cognitive decline may be expected (Grady and Craik, 2000).
Sixteen healthy elderly volunteers participated in the trial: eight men and eight women with a mean age of 66.1 (SD 4.46; range 60–75 years). Their medical histories indicated they were in good physical and mental health and this was verified by a medical examination, ECG, laboratory examination, and the mini mental state examination (MMSE), the scores of which fell within the normal range (MMSE mean=28.13, SD=1.73, range 25–30). Fifteen volunteers were right-handed and one was ambidextrous. Their educational level varied from primary education to academic schooling (primary n=3, lower n=2, intermediate n=3, higher n=6, academic n=2).
Drug Administration
The study was done according to a double-blind, placebo-controlled, randomized, four-way crossover design with placebo, 500 mg farampator, 2 mg biperiden, and 3 mg rivastigmine as drug conditions. Farampator reaches peak plasma concentrations around an hour after dosing and has a terminal half-life, which is <10 h. A safe and well-tolerated dose of 500 mg farampator was selected on the basis of a double-blind, placebo-controlled phase-I study in healthy young (mean age of 31.3 years) volunteers (n=29) in which a dose range from 15 to 1100 mg was investigated and shown to be devoid of serious or severe adverse events (P Dogterom et al, unpublished data). For biperiden peak plasma concentrations are reached 1–2 h after single dose administration followed by a rapid initial decline of the concentrations to 12% of the peak values after 6 h, and subsequently followed by a slow terminal elimination phase with concentrations close to or below detection limit at 48 h (Hollmann et al, 1987). For rivastigmine the time to reach maximum plasma concentration ranges from 0.8 to 1.2 h. In elderly healthy volunteers, the plasma elimination half-life is 0.9–1.3 h. Renal elimination of the drug's metabolites is rapid and essentially complete after 24 h. Inhibition of ChEi activity (in CSF) is significant by 1.2 h, reaches a peak by 2.4 h and declines slowly until approximately 8.5 h.
Randomization of treatments over the drugs was carried out according to a balanced latin-square design (repeated four times). There was a wash-out of a week and therefore no drug interaction or carryover effects are to be expected, because of the short elimination half times of the drugs and the unlikelihood of lasting effects after a single dose.
Testing took place 1 h after drug-intake, which is around Tmax. The data for biperiden and rivastigmine have been reported elsewhere (Wezenberg et al, 2005).
Tests, Apparatus, and Procedure
The tests employed during the study formed part of a more comprehensive cognitive and psychomotor test battery (Wezenberg et al, 2004). All tests were computerized and were run on a laptop computer for stimulus presentation, while data were recorded using a digitizing tablet (WACOM UD-1218-RE) and a pressure sensitive pen, which could also be used as a cursor. The x and y coordinates of the pen tip on and up to 5 mm above the digitizer were sampled with a frequency of 200 Hz and a spatial accuracy of 0.2 mm (De Jong et al, 1996).
To allow them to familiarize themselves with tests and procedures, the participants were invited to the hospital to perform a practice session during the week preceding the actual trial. For all tests there were five comparable versions for four test days and one practice day. The order of test version administration was counterbalanced across subjects. In an unpublished pilot study, we have assessed possible practice effects, and after an adequate practice these are minimal.
Assessment of Short-Term Memory and Working Memory
Symbol digit recall test
The Symbol digit recall test (SDRT) is based on the extended procedure of the Symbol Digit Substitution Test (SDST)—which will be discussed in the last paragraph of this section—as developed by Kaplan et al (1991) to measure incidental learning and follows directly after the SDST. In the present study, it was used to assess short-term memory. As soon as the participants had completed the SDST they were again presented with the same symbols from the SDST, now shown one at the time, and asked to point out the corresponding numbers (short-term memory or STM). After an interval of 30 min they were again to identify the correct numbers for the symbols (long-term memory or LTM). The outcome measures are the number of correctly identified symbols and the response times.
N-back task
To evaluate working memory an N-back task was chosen, a task widely used in schizophrenia research for the detection of deficits in working memory (Meyer-Lindenberg et al, 2001; Weinberger et al, 1996). In the present version the participants were presented with a sheet of paper depicting a start circle and six potential target circles that surround the start circle. As soon as a stimulus target circle appeared on the computer monitor, the participants needed to draw a line from the start circle towards and ending inside the corresponding target circle. In the 1-back condition, they had to respond to the stimulus that was presented in the previous trial and in the 2-back condition to the stimulus that was presented two trials before. Both conditions were performed twice and lasted a maximum of 2 min after which the number of correct trials was counted. Alternatively, the condition ended as soon as the participant had produced 25 correct trials. The outcome measure is ‘movement time to reach target’, which is faster when working memory is enhanced and slower when it is impaired. The measure ‘distance covered to reach target’ was added as a control variable because movement time is dependent on the distance covered. One participant did not take this test.
Assessment of Episodic Memory
Verbal memory test
We tested verbal memory with a modified version of the classical Auditory Verbal Learning Test (Vakil and Blachstein, 1993), which originally comprises 15 words that are presented verbally three times in the present study. Under normal circumstances subjects remember more words after each trial. In order to prevent ceiling effects, and to make the test more challenging especially because in elderly people the learning of word lists is compromised, we developed a series of four equivalent lists of 18 words. Immediately after each presentation and after a 5-min and a 30-min delay our participants were asked to recall as many words as they could. After the latter delayed recall trial they were presented with a list of 36 words from which they were asked to identify the 18 words from the original list. The supplementary words were distracters and resembled the correct words in a semantic or phonological manner. The outcome measures are the sum of the number of correct scores for the three immediate recall trials (total direct recall), the two delayed recall trials and the delayed recognition trial.
Picture memory task
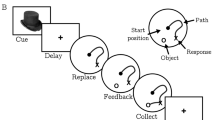
Visual memory or picture memory, has been used widely as a measure of both immediate and delayed recall for the detection of cognitive changes in a wide range of conditions. The picture memory task used in this study (for an example, see Figure 1) was modeled after a task Petrides and Milner (1982) developed for the detection and localization of brain damage. That visual memory tests are also sensitive for drug effects has been shown by numerous studies (Peretti et al, 1997; Rammsayer et al, 2000; Robbins et al, 1997).

Example of the picture memory task. The left panel depicts the sixteen symbols that needed to be memorized; the right panel shows the collection from which the sixteen correct symbols needed to be recognized.
In the first part of our test (immediate visual recognition) the participants were shown a matrix of 16 abstract pictographs or symbols that they were instructed to memorize. Next, they were presented with a matrix of 48 symbols from which they needed to identify the 16 original designs by pointing them out. The second part of the test started with the presentation of another matrix of 48 symbols, containing the earlier correctly remembered symbols. The participants were again asked to commit these designs to memory. They were subsequently asked to pick out the correctly identified symbols, but now only one at the time from several matrixes: after each choice the matrix was scrambled (using the same symbols) after which they had to point out the next correct symbol. The test ended when all symbols had been recognized or when two consecutive mistakes had been made. With this test visual recognition and self-ordered retrieval were evaluated and the outcome measures of both test components were the number of correct responses.
Assessment of Motor Learning
Maze learning task
To study the effect of glutaminergic modulation on motor learning we used the Maze learning task developed by Van Mier et al (1993).
In several studies (among which a PET study) it was shown that the maze task assesses motor learning unaided by sight (Van Mier et al, 1998; Wezenberg et al, 2005). The mazes (for an example, see Figure 2) are cutout structures, creating an enclosed path for the pen to traverse. Each maze design has a 0.5-cm wide and 0.15-cm deep path with a total length of 24 cm that consists of eight segments. To simplify decision-making, the maze features straight stretches with 90° turns. At each intersection subjects need to choose between two opposite directions, one of which leads to a dead end after 0.5 cm. The maze forms a closed loop, that is start and end points are the same. We used four comparable versions and a practice maze in the shape of a square. The participants were instructed to close their eyes and find their way through the maze and make as many loops as possible within 2 min. During the first 30 s they were presented with the practice maze after which they performed three trials with the actual maze followed by a trial with what they had been told was a new maze. This latter maze was effectively a rotated version of the original. The procedure was adopted to control for a general increase in motor-speed that might otherwise have masked signs of improved motor skill. The outcome measure was ‘the number of completed loops’ within 2 min.

Example of the maze test.
Pursuit task
To measure implicit procedural learning a computerized version of the rotor pursuit task was used. This test is based on the classical rotary pursuit task (Ammons, 1951; Siegel et al, 1996). It is a continuous motor task. Subjects had to follow the movement of a large target stimulus on the computer screen with a cursor by moving the pen over the XY-tablet. The speed of the target gradually increases when the cursor is contained within the target but slows down quite quickly when it is not. The target follows a spatially predictable circular path over the screen. The outcome measures for procedural learning are: the ‘total number of rotations’ and ‘Time per loop’ (ie one 360° tracking of the target) for the first five loops.
Assessment of Information Processing
Tangle
The tangle task is a purely visual task requiring high concentration. Subjects are instructed to visually track a particular line that winds through two to four other lines. Our test was modeled after the visualization test from the ‘kit for factor-referenced cognitive tests’ devised by French (1954) and Ekstrom et al (1976). It was selected by the US NAVY to study environmental and other time-course effects and has good task stability and reliability (Bittner et al, 1983, 1986). The tangled lines were presented on the laptop screen. The target line was indicated by a yellow square. On subsequent trials the tangles increased in complexity: they increased in length and included more 90° turns. The paper form depicted a start area and five target areas (numbered 1–5), which reflected the potential maximum target areas on the screen. The outcome measure is the number of correct trials within 2 min.
Symbol digit substitution test
The Symbol digit substitution test (SDST) is a modification of the subtest from the WAIS (Wechsler Adult Intelligence Scale; Wechsler, 1981). Subjects have to substitute symbols for the digits 1–9 on the basis of a given key. The outcome measure is the total number of correct digits completed in 90 s.
According to Hege et al (1997) and Lezak (1995) the test measures a multitude of cognitive components among which visuospatial scanning, intermediate memory, perceptual motor speed and speed of cognitive processing. In an attempt to disentangle these cognitive components we conducted additional analyses of the output data. Based on pen pressure, movement trajectories were defined as either pen-up or pen-down periods to allow the computation of matching times and movement (writing) times. For the motor component, the mean writing times were calculated and for the more cognitive component the mean matching times were derived. For more details of the method of analysis we refer to earlier publications (Jogems-Kosterman et al, 2001; Sabbe et al, 1999; van Hoof et al, 1998; Wezenberg et al, 2005).
Continuous trail making test
The continuous trail making test (CTMT) was develop based on Reitan's (1958) trail making test. Part A, the simplest section of the test, is used to measure visual scanning, motor speed and agility while the more complex part B assesses executive performance, that is set-shifting, working memory and divided attention. Numerous studies have proven the test to be sensitive to various neuropsychiatric conditions, for example schizophrenia (Lezak, 1995; Reitan and Wolfson, 1995).
In part A, subjects have to connect as many letters in alphabetical order as they can in 120 s. In part B, they have to connect as many numbers to their corresponding letters in the alphabet by alternating between the two sequences, for example 1A2B etc, in 120 s. The outcome measures are ‘total number connected’, ‘total number of errors’, and ‘type of errors’. In part B, one can erroneously connect numbers to numbers or letters to letters (NNLL) instead of numbers to letters. One can also commit so-called switching errors by connecting the numbers to the wrong letters or letters to the wrong numbers by connecting numbers to letters or vice versa in the wrong order, for example 1B2C instead of 1A2B (NLLN).
Control Measures
Serum concentrations
For each volunteer blood samples were taken at 55, 210, and 345 min after drug intake. In one female participant the last blood sampling did not succeed.
The farampator concentrations in the serum samples were determined by means of a liquid chromatographic assay with mass spectrometric detection (LC-MS) after solid phase extraction. Bioanalyses were performed at the Department of Clinical Pharmacology and Kinetics of NV Organon (Oss, the Netherlands).
Side effects
All participants were monitored for SEs by the research nurse and physician. The reported SEs were coded using MedDRA (6.1).
Alertness measures
To control for possible alterations in alertness as a confounder for memory improvements, subjects performed several tests measuring various aspects of alertness before dosing and 60 min after drug administration.
Measurements of saccadic eye movements (SEM) were chosen as the (neuro)physiological measure for alertness. Subjects need to visually follow a target that changes position suddenly and at random intervals. The target consists of an array of light-emitting diodes on a bar fixed at 50 cm in front of the head support. The size of the target shifts was fixed at 30° (15 from middle to left–right). The outcome measure is the overall mean peak velocity.
A simple (SRT) and a choice reaction time test (CRT) were used to assess behavioral alertness. In the SRT, a yellow square was presented on the monitor at random intervals (0.5–1.5 s). Subjects were instructed to press the response button on the response module as fast as possible after detection of the target. The trial consisted of 20 targets with mean reaction time as the outcome measure. In the CRT the words ‘left’ and ‘right’ were presented randomly on the monitor and subjects had to press the corresponding buttons on the response module as fast as possible. Again, per trial 20 targets were presented with the outcome measure mean reaction time.
For the assessment of subjective alertness the participants completed a mood rating scale (MRS; Bond and Lader, 1974). The three factors subjective alertness, calmness and contentment were derived from the self-report questionnaire comprising 16 visual analogue scales.
Statistical Analyses
Statistical evaluation (using SPSS 11 for Windows) was performed with GLM Repeated Measures Analysis of Variance (ANOVA) with ‘drugs’ as within-subject factor for all tests. For the verbal memory test (VMT), maze learning task and SDRT, also the results for the within-subject factor ‘trial’ and the interaction between ‘trial’ and ‘drugs’ were analyzed. Subsequently, univariate ANOVAs were performed with the grouping factor side effects (SEs: yes/no) for the farampator condition and the placebo condition separately.
RESULTS
Plasma Levels
Figure 3a presents the mean plasma levels of farampator. The highest concentrations were found in the first blood sample taken at 55 min after drug intake although it should be noted that the variation in plasma levels was very high at this time point. In the subsequent serum samples, farampator concentrations steadily declined and showed less variation. As only three blood samples were taken, it cannot be ruled out that the true peak concentrations occurred at another time (slightly earlier or later) than at the first measurement.

The left graph (a) depicts the mean plasma levels and standard error bars of farampator and the right graph (b) the level of farampator for participants with and without side effects.
Memory and Information Processing
Both the descriptive statistics and F-statistics of the test performance for the farampator and placebo conditions are presented in Table 1.
Short-term and working memory
As can be seen in Figure 4, farampator significantly improved the performance on the STM trial of the SDRT by enhancing the number of correctly remembered symbols. The improvement in the LTM trial did not reach significance.

The effect of farampator on the performance measures of the SDRT in terms of the number of correctly identified symbol-digit pairs during the short-term memory trials (STM) and the long-term memory trials (LTM).
Neither the 1-back nor the 2-back trials of the N-back task generated differences between placebo and farampator for the parameters movement time (MT) and distance covered.
Episodic memory
In the three immediate recall trials of the VMT all participants showed equal increases in the number of words they managed to recall in both conditions. Surprisingly, farampator tended to impair the performance in the delayed recall trials and significantly impaired the recognition performance relative to placebo.
In the picture memory test no significant effects were found for simple drug-placebo comparisons although farampator tended to impair performance in the second part of the test.
Motor learning
The number of completed loops increased non-significantly from trial 1 to 3 in the maze learning task. However, no significant differences were found in the performance between placebo and farampator. From trial 3 (learned maze) to trial 4 (the new/rotated maze) the number of completed loops decreased significantly, which suggests that the performance measures indeed reflect motor learning rather than motor speed. Again, the outcomes were comparable for the two conditions.
Similarly, no differences were found between placebo and farampator in the pursuit task for the drug-placebo comparisons in the parameters ‘number of rotations’ and ‘mean loop time’.
Information processing speed
Neither the SDST nor the tangle task revealed any drug-placebo differences. Also in the CTMT no significant drug-placebo effects were found for the number of correctly connected symbols or the total number of errors, although the participants did tend to make fewer switching errors (NLLN) in the farampator condition.
Control Measures
Side effects
Table 2 lists the reported SEs. Only the SEs that were reported more than once are listed. In the farampator condition more participants experienced headache and nausea than in the placebo condition. A high degree of somnolence was reported by almost one third of the participants, but to the same degree in both the drug and the placebo condition. All SEs were transient and self-limiting.
Figure 3b shows the differences in plasma levels of farampator relative to the presence and absence of SEs. A one-way ANOVA revealed significantly higher concentrations of farampator in the first serum sample (after 55 min) in participants that reported to experience SEs than in participants that did not (F(1,14)=6.59, p=0.022). This difference disappeared in the subsequent samples (after 210 and 345 min). The group differences in plasma levels for the specific SEs did not reach significance.
Alertness measures
Table 3 lists the descriptive statistics and F-statistics for the alertness measures.
No significant effects were found although the drug tended to interact (nonsignificantly) with time on the SEM: in the placebo condition peak velocity marginally decreased, whereas after farampator peak velocity appeared to increase over baseline.
Additional Analyses: The Relationship between Drug Effects on Memory, Information Processing and SEs
In view of the differences in plasma levels of farampator for the participants with and without SEs additional analyses were performed. Univariate ANOVAs were performed to reanalyze test performance with the grouping factor SE (yes/no), separately for placebo and for farampator SEs on placebo or farampator performance. No drug-placebo comparisons were made for the SE and SE-free groups, since the subjects who had SEs under the two conditions were not the same and because of sample-size considerations (N is too small). Table 4 presents the effect of the SEs on test performance.
Short-term and working memory
Whereas placebo-related SEs did not affect SDRT performance, the presence of drug-induced SEs did so significantly: although the performance of both groups was superior throughout the farampator condition, the participants without SEs showed significantly more improvement than the group with SEs.
Neither the presence of placebo-related nor drug-induced SEs affected performance in the N-back task.
Episodic memory
In the VMT, the placebo-related side effects had no effect on performance. The farampator-induced SEs on the other hand did affect performance: participants experiencing SEs performed worse than participants who did not. This applied to the immediate recall trials and both delayed recall trials (STM and LTM), but not to the recognition trial. The result for the delayed recall trials is depicted in Figure 5.

The effect (number of words recalled) of SEs on the VMT performance in the two delayed recall conditions (DR) for the placebo and farampator conditions; LMT=Long-Term Memory Trial approximately 30 min after learning the words.
In the picture memory task no significant effects were found for the placebo-related SEs. For the farampator-induced SEs a trend was found in the first trial: participants with SEs tended to perform worse than participants without SEs. No effects were found in the second trial.
Motor learning
No effects were found for the SEs reported in the placebo condition on the maze-learning task. There was an interaction between trial (4 and 3) and Group for the farampator-induced SEs: in the group of participants that did not report any SEs performance steadily improved from trial 1 to trail 3 and then dropped sharply in the new (rotated) maze in trial 4. In the group that did report SEs the performance enhancement was halted after the second trial, making the drop in trial 4 less severe.
The performance on the pursuit task was not affected by the presence or absence of either placebo-related or drug-induced SEs.
Information processing
No effects were found for the placebo-related SEs in the SDST, tangle task or CTMT and in the tangle task the drug-induced SEs also did not affect performance.
Farampator-induced SEs did affect SDST and CTMT performance to some extent. In both tasks the group without SEs showed a clearly superior performance than the group with SEs, but this was only marginally significant in the matching times in the SDST and the number of correctly connected pairs in the CTMT.
DISCUSSION
This is the first study to show effects of the ampakine farampator on memory and information processing. The results revealed that 500 mg farampator caused a clear-cut improvement in the short-term memory of healthy elderly volunteers, but appeared to impair episodic memory. In addition, it was found that the plasma levels of participants that reported SEs was higher than the levels found for the participants that did not experience any SEs. Post hoc analyses revealed that in contrast to placebo, farampator had selectively improved the memory performance in the group without SEs and not in the group with SEs.
Modulation of Cognitive Processes
Memory
Short-term memory was clearly enhanced after farampator: in the SDRT the drug always led to a superior performance, irrespective of the presence or absence of SEs, although the participants without SEs performed significantly better than the group with SEs.
Initially, the analyses of the episodic memory data generated negative effects of farampator both in the delayed recall conditions and in the recognition of verbal material. Supplementary analyses, however, revealed that the impairments predominantly occurred in the delayed free recall of the verbal material in the subjects that had reported drug-induced SEs; the effect was not found in the word recognition condition.
Analysis of the motor learning data initially yielded comparable results in that no effects were found when farampator was compared to placebo. Post hoc analyses demonstrated, however, that the presence of SEs had negatively affected maze learning in the farampator condition. The group without SEs showed a gradual increase in the number of loops they produced in the course of the test, indicating normal learning, whereas the group that had reported SEs did not show such progress in their performance.
The positive effects of farampator on memory are in line with the results for CX516 in human volunteers reported in the literature (Lynch, 2004). More important, although perhaps not surprising, was our finding that performance improvements tended to be dependent on the absence or presence of (subjective) side effects. Earlier studies had demonstrated that distractions (eg feeling unwell) could negatively affect performance because they tend to decrease concentration or divide attention (Carver, 1994; Lezak, 1995; Meyer et al, 2000; Schmidt and Lee, 1999). The adverse drug reactions that were reported in a study using high doses (900–1200 mg) of CX516 included mild headache, salivation and/or swallowing reactions and spontaneous leg movements (Lynch et al, 1996). Hence, barring headache, the adverse reactions our participants reported after farampator appear new, although it needs to be noted that to date reports on adverse reactions to ampakines are scarce. Our experimental setting, that is the long test days, the number of tests, the double-blind treatment and strange hospital environment, was likely to induce some tiredness and anxiety and perhaps even somnolence, headache, or nausea. To exclude the study environment as a cause for the SEs, the effects of the SEs reported for placebo were analyzed carefully. Apart from a few minor nonsignificant effects, they proved not to have affected performance. Although we did not explicitly check for the nature of placebo-related SEs, nor the intensity, time of onset and duration on performance, it is likely that they persisted less long and were less intense to have caused any effect on performance.
The overall lack of a positive effect of farampator on the episodic memory measures used in this study was unexpected. It was especially surprising for the VMT as in other studies using the ampakine CX516, it had been a nonsense syllable test that had generated the strongest effects (Lynch et al, 1996, 1997). It should be noted, however, that only those participants that had experienced SEs had shown a decrement in their performance. Moreover, this group proved to have higher plasma levels, which may imply that the beneficial effects on memory of ampakines are nonlinear, or that they are counteracted by the appearance of SE's at higher plasma levels. This may mean that improvements in performance are only induced by an optimal plasma level of farampator and that the dose of farampator may have been too high for some of the elderly subjects in this study.
Another tentative explanation for the negative results is that our memory tests may have been too difficult for this age-group. We opted for 18-item word lists and a set of 16 pictographs to prevent ceiling effects and to create a state of pseudo-dementia. However, lists comprising a maximum of ten items are more common in neuropsychological research in elderly subjects (Bouma et al, 1998; Lezak, 1995). It should be noted, though, that all the participants did show the expected learning curves in both the placebo and the drug conditions of the memory tasks.
Information processing
The effects of farampator on information processing are somewhat more subtle than its effects on memory. No significant effects were found in the first drug-placebo comparisons, although in the CTMT there was a trend indicating a decrease in the number of switching errors. This effect will be discussed in more detail in the section on ampakines for the treatment of schizophrenia.
The post hoc analyses also did not generate significant effects for the SDST, although it did reveal a trend: the participants without SEs were faster in matching the correct numbers to the symbols in the farampator condition than those with SEs.
For the tangle task, measuring more visuospatial processes, our extra analyses failed to detect a difference between the participants with or without SEs. Also in the CTMT, which requires matching as well as set-shifting skills, no significant effects were found although in the group without SEs the number of correctly connected letters and numbers proved to have increased noticeably in the farampator condition.
As ours are the first data reflecting the effects of an ampakine on information processing, it is not possible to compare the present results with other data from literature. To date, attention has mainly been focused on memory tests to examine cognitive effects of ampakines. One study did assess psychomotor performance with a finger tapping test and a digit cancellation test (Ingvar et al, 1997) but could not report any effects. The tests applied in the present study contained more prominent cognitive components than motor-speed components.
Ampakines and Schizophrenia
The positive results on short-term memory and the favorable trends in the trail making test (CTMT) are of interest in view of the development of farampator for schizophrenia. Memory is one of the cognitive domains that are found to be frequently impaired in schizophrenia (Nuechterlein et al, 2004). Although the CTMT version we used is considered as a test for processing speed (Nuechterlein et al, 2004), it also measures another process due to the necessity to shift continuously between numbers and letters. It is well known that set-shifting is also often disturbed in schizophrenia (Donohoe and Robertson, 2003; Pantelis et al, 1999) and it has even been identified as a predictor of relapse in first-episode schizophrenia (Chen et al, 2005).
Interestingly, in animal research it has been established that AMPA and NMDA receptors are essential in facilitating working memory and set-shifting. Schmitt et al (2005) demonstrated that gene-targeted mice lacking the AMPA receptor subunit GluR-A have deficits in hippocampal long-term potentiation (LTP) and spatial working memory deficits. In their study Stefani et al (2003) showed that blockage of AMPA receptors in the medial prefrontal cortex impaired set shifting in a maze-based task where rats were required to switch between brightness and texture discrimination strategies.
Suggestions for Future Research
The present study compared farampator with placebo at a single-dose level. It would be interesting to see whether future research will replicate these findings at other doses, with special emphasis on the lower doses, as the present findings indicate that impairments are more likely to occur at the higher plasma levels of farampator, which also induce side effects. It would also be worthwhile to study the effects of ampakines in other age-groups than the healthy elderly subjects of the current study. It is known that aging causes neurobiological changes, including metabolic and neurochemical changes, as well as changes in functional circuitry (Li and Dinse, 2002). Possibly, the interaction of SEs with drug-condition is partly caused for a large part by the slower metabolic processes in older age.
As farampator was developed to ameliorate the cognitive deficits seen in schizophrenia, the experimental results of the present study using healthy volunteers need to be compared with the findings from randomized controlled studies with patients.
CONCLUSION
Farampator has a significant, positive effect on short-term memory functioning in healthy, elderly volunteers, regardless of plasma level or SEs. In addition, a modest reduction of errors was found in a set-switching task. In the participants that did experience side effects, episodic memory was impaired, but in subjects that did not experience side effects memory processes tended to be enhanced, though this did not reach the significance level.
References
Ammons RB (1951). Effect of distribution of practice on rotary pursuit hits. J Exp Psychol 41: 17–22.
Arai A, Kessler M, Xiao P, AmbrosIngerson J, Rogers G, Lynch G (1994). A centrally active-drug that modulates Ampa receptor gated currents. Brain Res 638: 343–346.
Bittner AC, Carter RC, Kennedy RS, Harbeson MM, Krause M (1986). Performance evaluation tests for environmental-research (Peter)—evaluation of 114 measures. Percept Mot Skills 63: 683–708.
Bittner AC, Carter RC, Krause M, Kennedy RS, Harbeson MM (1983). Performance evaluation tests for environmental-research (Peter)—moran and computer batteries. Aviat Space Environ Med 54: 923–928.
Bond A, Lader M (1974). Use of analog scales in rating subjective feelings. Br J Med Psychol 47: 211–218.
Bouma A, Mulder J, Lindeboom J (1998). Neuropsychologische diagnostiek, handboek. Zwets & Zeitlinger: Lisse.
Carver CS (1994). Cognitive-processes and self-regulation—determinants of concentration and distraction. Appl Psychol Int Rev 43: 387–391.
Chen EYH, Hui CLM, Dunn ELW, Miao MYK, Yeung WS, Wong CK et al (2005). A prospective 3-year longitudinal study of cognitive predictors of relapse in first-episode schizophrenic patients. Schizophr Res 77: 99–104.
de Jong WP, Hulstijn W, Jogems-Kosterman BJM, Smits-Engelsman BCM (1996). OASIS software and its application in experimental handwriting research. In: Simmers ML, Leedham CG, Thomassen AJWM (eds). Handwriting and Drawing Research: Basic and Applied Issues. IOS: Amsterdam.
Donohoe G, Robertson IH (2003). Can specific deficits in executive functionina explain the negative symptoms of schizophrenia? A review. Neurocase 9: 97–108.
Ekstrom RB, French JW, Harmon H, Dermen D (1976). Manual for the Kit of Factor Referenced Cognitive Tests. Educational Testing Service: Princeton, New York.
French JW (1954). Manual for Kit of Selected Tests for Reference Aptitude and Achievement Factors. Educational Testing Service: Princeton, New York.
Foster JK, Lidder PG, Sunram SI (1998). Glucose and memory: fractionation of enhancement effects? Psychopharmacology (Berlin) 137: 227–259.
Goff DC, Leahy L, Berman I, Posever T, Herz L, Leon AC et al (2001). A placebo-controlled pilot study of the ampakine CX516 added to clozapine in schizophrenia. J Clin Psychopharmacol 21: 484–487.
Grady CL, Craik FI (2000). Changes in memory processing with age. Curr Opin Neurobiol 10: 224–231.
Granger R, Deadwyler S, Davis M, Moskovitz B, Kessler M, Rogers G et al (1996). Facilitation of glutamate receptors reverses an age-associated memory impairment in rats. Synapse 22: 332–337.
Granger R, Staubli U, Davis M, Perez Y, Nilsson L, Rogers GA et al (1993). A drug that facilitates glutamatergic transmission reduces exploratory activity and improves performance in a learning-dependent task. Synapse 15: 326–329.
Green MF, Nuechterlein KH, Gold JM, Barch DM, Cohen J, Essock S et al (2004). Approaching a consensus cognitive battery for clinical trials in schizophrenia: the NIMH-MATRICS conference to select cognitive domains and test criteria. Biol Psychiatry 56: 301–307.
Hampson RE, Rogers G, Lynch G, Deadwyler SA (1998). Facilitative effects of the ampakine CX516 on short-term memory in rats: Enhancement of delayed-nonmatch-to-sample performance. J Neurosci 18: 2740–2747.
Hege SG, Ellinwood Jr EH, Wilson WH, Helligers CA, Graham SM (1997). Psychomotor effects of the anxiolytic abecarnil: a comparison with lorazepam. Psychopharmacology (Berlin) 131: 101–107.
Heinrichs RW, Zakzanis KK (1998). Neurocognitive deficit in schizophrenia: a quantitative review of the evidence. Neuropsychology 12: 426–445.
Hollmann M, Muller-Peltzer H, Greger G, Brode E, Perucca E, Grimaldi R (1987). Pharmacokinetic-dynamic study on different oral biperiden formulations in volunteers. Pharmacopsychiatry 20: 72–77.
Ingvar M, AmbrosIngerson J, Davis M, Granger R, Kessler M, Rogers GA et al (1997). Enhancement by an ampakine of memory encoding in humans. Exp Neurol 146: 553–559.
Jogems-Kosterman BJM, Zitman FG, van Hoof JJM, Hulstijn W (2001). Psychomotor slowing and planning deficits in schizophrenia. Schizophr Res 48: 317–333.
Johnson SA, Luu NT, Herbst TA, Knapp R, Lutz D, Arai A et al (1999). Synergistic interactions between ampakines and antipsychotic drugs. J Pharmacol Exp Ther 289: 392–397.
Kaplan E, Fein D, Morris R, Delis D (1991). Wais-R as a neuropsychological instrument. The Psychological Corporation: San Antonio, TX.
Larson J, Lieu T, Petchpradub V, Leduc B, Ngo H, Rogers GA et al (1995). Facilitation of olfactory learning by a modulator of AMPA receptors. J Neurosci 15: 8023–8030.
Larson J, Quach CN, Leduc BQ, Nguyen A, Rogers GA, Lynch G (1996). Effects of an AMPA receptor modulator on methamphetamine-induced hyperactivity in rats. Brain Res 738: 353–356.
Lebrun C, Pilliere E, Lestage P (2000). Effects of S 18986-1, a novel cognitive enhancer, on memory performances in an object recognition task in rats. Eur J Pharmacol 401: 205–212.
Lezak MD (1995). Neuropsychological Assessment, 3rd edn. Oxford University Press: New York.
Li SC, Dinse HR (2002). Aging of the brain, sensorimotor, and cognitive processes. Neurosci Biobehav Rev 26: 729–732.
Lynch G (2004). AMPA receptor modulators as cognitive enhancers. Curr Opin Pharmacol 4: 4–11.
Lynch G (1998). Memory and the brain: unexpected chemistries and a new pharmacology. Neurobiol Learn Memory 70: 82–100.
Lynch G, Granger R, Ambros-Ingerson J, Davis CM, Kessler M, Schehr R (2003). Evidence that a positive modulator of AMPA-type glutamate receptors improves delayed recall in aged humans. Exp Neurol 145: 89–92.
Lynch G, Granger R, AmbrosIngerson J, Davis CM, Kessler M, Schehr R (1997). Evidence that a positive modulator of AMPA-type glutamate receptors improves delayed recall in aged humans. Exp Neurol 145: 89–92.
Lynch G, Kessler M, Rogers G, AmbrosIngerson J, Granger R, Schehr RS (1996). Psychological effects of a drug that facilitates brain AMPA receptors. Int Clin Psychopharmacol 11: 13–19.
Marenco S, Egan MF, Goldberg TE, Knable MB, McClure RK, Winterer G et al (2002). Preliminary experience with an ampakine (CX516) as a single agent for the treatment of schizophrenia: a case series. Schizophr Res 57: 221–226.
Meyer JS, Thornby J, Crawford K, Rauch GM (2000). Reversible cognitive decline accompanies migraine and cluster headaches. Headache 40: 638–646.
Meyer-Lindenberg A, Poline JB, Kohn PD, Holt JL, Egan MF, Weinberger DR et al (2001). Evidence for abnormal cortical functional connectivity during working memory in schizophrenia. Am J Psychiatry 158: 1809–1817.
Nuechterlein KH, Barch DM, Gold JM, Goldberg TE, Green MF, Heaton RK (2004). Identification of seperable cognitive factors in schizophrenia. Schizophr Res 72: 29–39.
Pantelis C, Barber FZ, Barnes TRE, Nelson HE, Owen AM, Robbins TW (1999). Comparison of set-shifting ability in patients with chronic schizophrenia and frontal lobe damage. Schizophr Res 37: 251–270.
Peretti CS, Danion JM, Kauffmannmuller F, Grange D, Patat A, Rosenzweig P (1997). Effects of haloperidol and amisulpride on motor and cognitive skill learning in healthy volunteers. Psychopharmacology (Berlin) 131: 329–338.
Petrides M, Milner B (1982). Deficits on subject-ordered tasks after frontal-lobe and temporal-lobe lesions in man. Neuropsychologia 20: 249.
Rammsayer TH, Rodewald S, Groh D (2000). Dopamine-antagonistic, anticholinergic, and GABAergic effects on declarative and procedural memory functions. Brain Res Cogn Brain Res 9: 61–71.
Reitan RM (1958). Qualitative versus quantitative mental changes following brain-damage. J Psychol 46: 339–346.
Reitan RM, Wolfson D (1995). Category test and trail making test as measures of frontal-lobe functions. Clin Neuropsychol 9: 50–56.
Robbins TW, Semple J, Kumar R, Truman MI, Shorter J, Ferraro A et al (1997). Effects of scopolamine on delayed-matching-to-sample and paired associates tests of visual memory and learning in human subjects: comparison with diazepam and implications for dementia. Psychopharmacology (Berlin) 134: 95–106.
Sabbe B, Hulstijn W, van Hoof J, Tuynman-Qua HG, Zitman F (1999). Retardation in depression: assessment by means of simple motor tasks. J Affect Disord 55: 39–44.
Schmidt RA, Lee TD (1999). Motor Control and Learning: A Behavioral Emphasis, 3rd edn. Human Kinetics Publishers: USA.
Schmitt WB, Sprengel R, Mack V, Draft RW, Seeburg PH, Deacon RMJ et al (2005). Restoration of spatial working memory by genetic rescue of GluR-A-deficient mice. Nat Neurosci 8: 270–272.
Shors TJ, Servatius RJ, Thompson RF, Rogers G, Lynch G (1995). Enhanced glutamatergic neurotransmission facilitates classical-conditioning in the freely moving rat. Neurosci Lett 186: 153–156.
Siegel BV, Trestman RL, O’Flaithbheartaigh S, Mitropoulou V, Amin F, Kirrane R et al (1996). D-amphetamine challenge effects on wisconsin card sort test. Performance in schizotypal personality disorder. Schizophr Res 20: 29–32.
Staubli U, Perez Y, Xu FB, Rogers G, Ingvar M, Stoneelander S et al (1994a). Centrally active modulators of glutamate receptors facilitate the induction of long-term potentiation in-vivo. Proc Nat Acad Sci USA 91: 11158–11162.
Staubli U, Rogers G, Lynch G (1994b). Facilitation of glutamate receptors enhances memory. Proc Nat Acad Sci USA 91: 777–781.
Stefani MR, Groth K, Moghaddam B (2003). Glutamate receptors in the rat medial prefrontal cortex regulate set-shifting ability. Behav Neurosci 117: 728–737.
Vakil E, Blachstein H (1993). Rey auditory-verbal learning test—structure-analysis. J Clin Psychol 49: 883–890.
van Hoof JJM, Jogems-Kosterman BJM, Sabbe BGC, Zitman FG, Hulstijn W (1998). Differentiation of cognitive and motor slowing in the Digit Symbol Test (DST): differences between depression and schizophrenia. J Psychiatr Res 32: 99–103.
Van Mier H, Hulstijn W, Petersen SE (1993). Changes in motor planning during the acquisition of movement patterns in a continuous task. Acta Psychol 82: 291–312.
Van Mier H, Tempel LW, Perlmutter JS, Raichle ME, Petersen SE (1998). Changes in brain activity during motor learning measured with PET: effects of hand of performance and practice. J Neurophysiol 80: 2177–2199.
Wechsler D (1981). The psychometric tradition—developing the wechsler adult intelligence scale. Contemp Educ Psychol 6: 82–85.
Weinberger DR, Mattay V, Callicott J, Kotrla K, Santha A, vanGelderen P et al (1996). fMRI applications in schizophrenia research. Neuroimage 4: S118–S126.
Wezenberg E, Hulstijn W, Sabbe BGC, Ruigt GSF, Verkes RJ (2004). Psychomotor and cognitive effects of lorazepam and d-amphetamine in healthy subjects. Development of sensitive screening tests of psychomotor- and cognitive functions. Eur Neuropsychopharmacol 14: S362.
Wezenberg E, Verkes RJ, Sabbe BGC, Ruigt GSF, Hulstijn W (2005). Modulation of memory and visuospatial processes by biperiden and rivastigmine in elderly healthy subjects. Psychopharmacology (Berlin) 181: 582–594.
Yamada KA (1998). Modulating excitatory synaptic neurotransmission: Potential treatment for neurological disease? Neurobiol Dis 5: 67–80.
Acknowledgements
We thank them for their financial and technical support. We also thank Jan Leijtens and Sevla Cinar (MD) for their valuable assistance in the collection and analysis of the data.
Author information
Authors and Affiliations
Corresponding author
Additional information
Conflict of interest
Funding for the study was provided by NV Organon.
Rights and permissions
About this article
Cite this article
Wezenberg, E., Jan Verkes, R., Ruigt, G. et al. Acute Effects of the Ampakine Farampator on Memory and Information Processing in Healthy Elderly Volunteers. Neuropsychopharmacol 32, 1272–1283 (2007). https://doi.org/10.1038/sj.npp.1301257
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.npp.1301257
Keywords
This article is cited by
-
Pharmacological targeting of cognitive impairment in depression: recent developments and challenges in human clinical research
Translational Psychiatry (2022)
-
Distinct contributions of GluA1-containing AMPA receptors of different hippocampal subfields to salience processing, memory and impulse control
Translational Psychiatry (2022)
-
Augmenting cognitive work: a review of cognitive enhancement methods and applications for operational domains
Cognition, Technology & Work (2022)
-
Cellular and Molecular Changes in Hippocampal Glutamate Signaling and Alterations in Learning, Attention, and Impulsivity Following Prenatal Nicotine Exposure
Molecular Neurobiology (2020)
-
Postmortem Brain: An Underutilized Substrate for Studying Severe Mental Illness
Neuropsychopharmacology (2014)
