
Abstract
Introduction:
Survivors of esophageal atresia and tracheo–esophageal fistula (EA-TEF) often suffer chronic respiratory tract disease. EA-TEF results from abnormal emergence of the trachea from the foregut. This study in a rat model tests the hypothesis that primary lung maldevelopment might be a downstream consequence of this defect.
Results:
The lung was hypoplastic in rats with EA-TEF although the histological pattern was normal. Maturation and arteriolar wall thickness were unchanged, but mesenchymal control of airway branching was weakened. This branching was deficient from embryonal day (E13) on in adriamycin-treated explants.
Discussion:
In conclusion, the lungs were hypoplastic in rats with experimental EA-TEF due to defective embryonal airway branching. However, arteriolar wall and respiratory epithelial patterns remained normal. These findings suggest that similarly defective lung development might contribute to chronic respiratory disease in EA-TEF patients.
Methods:
Pregnant rats received either 1.75 mg/kg i.p. adriamycin or vehicle on E7, E8, and E9. Lungs were recovered at E15, E18, and E2. Lung weight/body weight ratio, total DNA and protein, radial alveolar count, arteriolar wall thickness, lung maturity, and mesenchymal control of airway branching were assessed. E13 lungs were cultured for 72 h and explant airway branching was measured daily. For comparisons, nonparametric tests (*P < 0.05) were used.
Similar content being viewed by others

Main
In most cases of esophageal atresia (EA), the upper esophagus ends in a blind pouch whereas the distal esophagus is replaced by a tracheo–esophageal fistula (TEF). This malformation (EA-TEF) occurs in 1:2,500 to 3,000 births, and as current survival approaches 90% (1), long-term sequelae have become the focus of attention.
EA-TEF is often associated with other malformations that may cluster in syndromes (Feingold), or VACTERL (Vertebral, Anal, Cardiac, Tracheo-esophageal, Renal, and Limb) or CHARGE (Coloboma, Heart, choanal Atresia, growth Retardation, esophageal atresia, Genitourinary, Ear) associations. Respiratory tract malformations can also occur, and this is not surprising because defective division of the embryonal foregut into esophagus and trachea (2) is the main pathogenic mechanism involved.
Survivors of EA-TEF often experience hoarse cough, repeated pneumonia or atelectasis, bronchiectasis, and chronic bronchitis. Peptic bronchitis (3), gastro-esophageal reflux (4), or the fistula stump account for these sequelae, but malformations of the airway or lung parenchyma (3) or lung hypoplasia might also play a role (5).
Research on the structure of the respiratory tract in human EA-TEF was limited for various reasons, and animal models became necessary for this purpose. Relatively straightforward pharmacologic models were preferred to knockout mice for retinoic acid receptors (6), thyroid transcription factor 1 (TTF-1) (7), shh/Gli2-Gli3 cascade genes (8), fibroblast growth factor-10 (Fgf10) (9), and Foxf1 (10), Nog (11), and Hox genes (12) in which various components of the VACTERL phenotype are also reproduced. Administration of adriamycin (doxorubicin) to pregnant rats (13) and mice (14) accurately reproduced a phenotype of EA-TEF with VACTERL syndrome (13,15,16). Abnormal embryology of the esophagus (2) and of other organs in this model allowed better understanding of some pathogenic mechanisms (17).
We hypothesized that the abnormal mechanisms of tracheo–esophageal separation leading to EA-TEF might extend downstream, compromising airway branching and, ultimately, lung growth and structure. If such is the case, even subtle deficiencies of broncho-alveolar branching, overall growth at birth, or pulmonary maturity could participate both in neonatal or in late respiratory tract disease in these patients. This study tests this hypothesis in rats with experimental EA-TEF.
Results
The numbers of control and adria-exposed broods were 20 and 35, respectively, and the corresponding numbers of embryos and fetuses were 217 and 199, respectively. The proportion of EA-TEF in treated mothers was 79.4%.
-
1
Lung measurements: Figure 1 depicts lung weights expressed as percentages of fetal weight. There was significant lung hypoplasia in fetuses treated with adriamycin with EA-TEF (Adria EA) fetuses and a similar trend for fetuses treated with adriamycin without EA-TEF (Adria noEA) ones. There were no differences between the two groups of pups that were exposed to adriamycin. Radial alveolar count showed that airway branching was significantly decreased in both groups of adria-treated rats in comparison with controls. However, these animals had a compensatory larger alveolar diameter. The arterial vessels were apparently identical in all groups, with no differences in pulmonary artery wall thickness. Figure 2 shows that total DNA and protein were diminished at embryonal day 21 (E21) in adria-treated lungs in comparison with controls, revealing decreased cell mass. TTF-1 protein was unchanged in adria-exposed animals. It should be noted that due to the lack of morphometric differences between animals with and without EA-TEF, only those with the malformation were tested.
Figure 1 
Lungs of Adria EA rats were hypoplastic (decreased lung weight/body weight ratio (lung W/body W)) (a) in comparison with controls (n = 21) due to deficient airway branching (decreased RAC) (b). The sacculo-alveolar spaces (c) were wider and the arteriolar wall thickness (d) was unchanged. There was no difference between Adria EA (n = 31) and Adria noEA fetuses (n = 9). Means ± SD. *P < 0.05. Adria EA, treated with adriamycin and EA-TEF; Adria noEA, treated with adriamycin without EA-TEF; EA-TEF, esophageal atresia and tracheo–esophageal fistula; RAC, radial alveolar count.
Figure 2 
Lung cell mass of treated rats was smaller (decreased (a) DNA and (b) protein) than in controls. However, epithelial maturation was unchanged (similar (c) TTF-1 protein). Adria EA and Adria noEA lungs were pooled due to their similar weight and histology. Means ± SD. *P < 0.05 and §NS. Adria EA, treated with adriamycin and EA-TEF; Adria noEA, treated with adriamycin without EA-TEF; EA-TEF, esophageal atresia and tracheo–esophageal fistula; NS, not significant; TTF-1, thyroid transcription factor 1.
-
2
Lung histology: The histologic pattern of the lungs near term was quite similar in the three groups. The organs were at the canalicular-saccular stage of maturation with quite well-developed air spaces separated by thin walls in which the contact with capillaries was apparently normal. Respiratory epithelium was mainly constituted by cuboidal type II pneumocytes that stained normally for TTF-1 ( Figure 3 ).
Figure 3 
Lung histologies in Adria EA and Adria noEA fetuses were similar to controls except for slightly wider air spaces (hematoxylin-eosin(HE), ×100, a–c). Epithelial maturation was similar in the three groups (TTF-1 immunostaining ×200, d–f). Bars = 100 µm. Adria EA, treated with adriamycin and EA-TEF; Adria noEA, treated with adriamycin without EA-TEF; EA-TEF, esophageal atresia and tracheo–esophageal fistula; TTF-1, thyroid transcription factor 1.
-
3
Molecular control of branching: Fgf10 mRNA was similar in the lungs of E15 and E18 animals as compared with controls, but it was significantly decreased near term ( Figure 4 ). Again here, only animals with EA-TEF were assessed.
Figure 4 
Lung branching regulation (Fgf10 mRNA) of Adria EA rats was similar to that of controls on the 15th and 18th gestational days, but it was significantly deficient on the 21st day. Adria EA and Adria noEA lungs were pooled due to their similar weight, RAC, and histology. Means ± SD. *P < 0.05. Adria EA, treated with adriamycin and EA-TEF; Adria noEA, treated with adriamycin without EA-TEF; EA-TEF, esophageal atresia and tracheo–esophageal fistula; RAC, radial alveolar count.
-
4
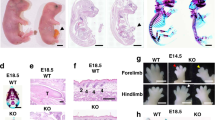
Embryonal tracheo-bronchial branching: As shown in Figure 5 , E13 explants (0 h) were smaller in embryos from both adriamycin-exposed groups, and they developed poorly in the ensuing days in comparison with controls. In Adria EA embryos, the fistula was apparent (arrows) and showed some distal branching after 48 to 72 h in culture. The structure of the large bronchi was abnormal in some Adria noEA embryos with some cystic dilatations (dots). The histograms in Figure 6 show that explant surface and perimeter were significantly smaller in adria-exposed embryos in comparison with controls. The same poor development was seen in terms of terminal buds.
Figure 5 
Airway branching was defective in the explanted lungs (13th gestational day) of both Adria EA and Adria noEA embryos. This deficiency persisted for 72 h. Notice the TEF in explants from embryos with EA (arrows) and the cystic dilatations of the trachea and bronchi in those without it (dots). (Pictures taken at ×40 original augmentation. Bar = 0.5 mm at 0 h. On subsequent days, photographic reduction to fit into the picture.) Adria EA, treated with adriamycin and EA-TEF; Adria noEA, treated with adriamycin without EA-TEF; EA-TEF, esophageal atresia and tracheo–esophageal fistula.
Figure 6 
In vitro deficient airway branching in terms of airway surface, airway perimeter, and number of terminal buds of lung explants from Adria EA (white bars) and Adria noEA embryos (black bars) in comparison with controls (gray bars). No differences between Adria EA and Adria noEA embryos. Means ± SD. *P < 0.05. Adria EA, treated with adriamycin and EA-TEF; Adria noEA, treated with adriamycin without EA-TEF; EA-TEF, esophageal atresia and tracheo–esophageal fistula.






Discussion
Our results demonstrate that in the most widely used animal model of EA-TEF, the lungs were definitely hypoplastic as a consequence of impaired airway branching. However, as shown by TTF-1 immunostaining and protein expression, neither the maturational status at term nor the arteriolar wall thickness were significantly affected. TTF-1 or Nkx.2.1 is downregulated in lung hypoplasia associated with congenital diaphragmatic hernia (18) in which there is also arteriolar wall thickening and hyper-reactivity, which are absent in EA-TEF. This transcription factor is closely related to tracheo-esophageal cleavage and to lung development because TTF-1−/− mice have rudimentary lungs and abnormal tracheo-esophageal separation (7).
The experiments with explants showed that this pulmonary hypoplasia was determined very early. In these experiments, adriamycin was given between E7 and E9, and tracheo-esophageal cleavage occurred in rats between E11 and E12 (2). The lungs were cultured on E13, and, at that point, adriamycin-exposed embryonal organs were already smaller and less branched than controls. Their development remained impaired until the end of the experiment, and it is significant that Fgf10, a key mesenchymal regulator of the process of endodermal branching (19,20,21,22) whose expression is delayed in mouse embryos exposed to adriamycin (23), was also downregulated in rats with EA-TEF at the end of gestation.
The role of airway tract decompression through the fistula as a possible explanation for this lung hypoplasia in other conditions, like fetal tracheostomy (24), should be ruled out in our experiments, because it occurred both in embryos with and in embryos without fistula. It is of particular interest that there were some abnormal dilatations of the large bronchi in adriamycin-exposed animals and that the distal part of the fistula, isolated from the embryonal environment, was able to branch, probably as a result of local endodermal–mesenchymal molecular interactions. These observations are not the focus of this study and require further investigation.
Respiratory tract defects were observed many years ago in EA-TEF survivors: lung agenesis (25) or hypoplasia (26), tracheal stenosis (27), and pulmonary cysts (28) or sequestrations (29) were occasionally reported. Respiratory malformations were found in 2.8% of patients and in 13.2% of autopsies, suggesting that they might be underdiagnosed (30). This proportion amounted to 13.3% in VACTERL patients (31), and it approached 50% when malformations were specifically examined (32).
Respiratory tract symptoms are constant in EA-TEF patients. Tracheomalacia related to abnormal tracheal rings or tracheal compression is quite common and usually tapers off with more or less energetic treatment weeks or months after EA-TEF repair (33,34). Other symptoms may persist for years. Of 100 EA-TEF survivors, 78 had more than 3 episodes of bronchitis/year that lasted much longer in 48%. Recurrent milk inhalation was considered to be a likely explanation (3). Repeated pneumonia occurs in up to 30% of survivors (35), constrictive bronchitis in 40–70% (36,37), obstructive or restrictive respiratory disease in 18–35% (36,37,38), and bronchial hyper-reactivity in more than 20% of them (36,37). The possible influence of gastro-esophageal reflux on these symptoms was repeatedly evoked, but other causes were also mentioned (4,36). Up to one-third of EA-TEF patients had impaired quality of life (39), and 32% of those aged 5 to 10 y and 8% of those older than 15 y missed school more than 2 wk per year for these reasons (35). A recent follow-up study of a large cohort of adult EA-TEF survivors revealed that 21% had restrictive respiratory disease, 21% had obstructive respiratory disease, and 36% had both ventilatory defects. In addition, 41% had bronchial hyper-reactivity often consistent with asthma, and these patients had overall impaired respiratory quality of life in comparison with a group of age-matched controls (40). Postoperative rib fusions, scoliosis, and gastro-esophageal reflux were found to account in part for these respiratory sequelae, but other explanations are possible.
Unfortunately, autopsy lung material from EA-TEF patients is relatively scarce, and, when available, it is rarely suitable for structural and molecular studies. A study of 3.3 million births from California revealed that 53 of 893 infants with EA-TEF also had lung hypoplasia. This diagnosis was firmly established upon autopsy in most cases, although unfortunately, no histological description was made (5). These observations demonstrated that patients with EA-TEF can have abnormal lung structure, particularly when they have associated malformations, and this could account for respiratory morbidity.
The use of a pharmacologic rodent model for these investigations seems largely justified as all features of VACTERL association are strikingly reproduced: laryngo–tracheal ring anomalies leading to stenosis and tracheomalacia are similar in rats and infants with EA-TEF (41,42,43), and the entire spectrum of communicating or noncommunicating foregut malformations seen in infants (sequestrations, cysts, duplications) is reproduced in rats (44,45,46,47). If the embryologic mechanisms of EA-TEF in infants and rats are similar and the malformations observed in the respiratory tract are identical, it is likely that failures of parenchymal development are also of the same nature in both settings. Previous studies on lung parenchyma in the adriamycin rat model found fairly normal canalicular–saccular patterns (48) and hypoplasia with reduced density of type I pneumocytes (49), although, unfortunately, the authors did not attempt to assess branching or lung mass.
Finally, the issue of whether lung growth deficiencies similar to those demonstrated in the rat fit with the pattern of respiratory symptoms observed in EA-TEF survivors should be discussed. In the neonatal period, shortly after repair, there are enough explanations for these symptoms: thoracotomy itself, pneumothorax, peptic bronchitis, the fistula stump, aspiration, and other. Deficient drainage through the sometimes-misplaced upper-right lobe bronchus causes atelectasis (50). Recurrent fistula may produce discrete or severe symptoms (51). The fact that newborns with EA-TEF do not suffer symptoms of lung immaturity or persistent pulmonary hypertension like those observed in congenital diaphragmatic hernia (another condition in which marked lung hypoplasia is present (ref. 52,53)) fits the pattern of less conspicuous lung hypoplasia described in our study. In a parallel investigation of the long-term respiratory morbidity of EA-TEF and congenital diaphragmatic hernia survivors, it was demonstrated that 25% of cases in both groups had forced expiratory volume decreased by −2 z scores and that both had similar degrees of compromise of maximal exercise performance (54). Pulmonary hypoplasia persisting after the neonatal period will be a likely explanation. Mild restrictive lung volumes were measured in EA-TEF survivors beyond the age of 40 (55), and as this restrictive pulmonary disease was similar in patients with and without gastro-esophageal reflux, tracheomalacia, and stenosis, alternative explanations were considered necessary (56).
This study consistently demonstrates that lung development is deficient in rats with EA-TEF in which most features of the human condition are faithfully reproduced. Structural studies of lung development in autopsies of patients are necessary to examine if these lesions are also present.
Methods
Experimental Design
After IdiPAZ Animal Care Committee approval (AP 7795-2010), timed pregnant rats (gestational day 0, E0 = sperm in vaginal smear) were treated with either 1.75 mg/kg i.p. adriamycin (Farmiblastina. Pharmacia, Madrid, Spain) or saline on E7, E8, and E9. Three groups of offspring were compared: Control, Adria noEA, and Adria EA. Lung weight and cell mass, airway branching, parenchymal structure, arteriolar wall thickness, and epithelial maturity (TTF-1) were examined at E21. Mesenchymal control of branching (Fgf10) was assessed at E15, E18, and E21. Embryonal airway branching was observed for 3 d in cultured lungs explanted on E13 aiming at ruling out external influences including those derived from urinary contribution to amniotic fluid (57).
Lung Harvesting and Structural Studies
At the elected end points, animals were killed and the embryos were dissected. The lungs were retrieved, weighed, and processed. DNA was extracted from snap-frozen tissue with the QIAamp DNA Micro kit (Qiagen, Las Rozas, Spain), and total DNA content was determined with a spectrophotometer (Nano Drop; Fisher Scientific, Madrid, Spain) at 260 µm. Total protein was measured using the BCATM protein assay kit (Pierce; Thermo Fisher Scientific, Rockford, IL). Histologic and immunohistochemical studies were carried out in fixed material. After tracheal cannulation, 4% paraformaldehyde was injected at <10 cm/H2O for alveolar distention, and the lungs were then fixed for 24 h. After inclusion in paraffin, 5-µm sections were stained with hematoxylin and eosin, periodic acid, and Masson stainings. Immunohistochemical staining was performed using standard techniques with anti-TTF-1 antibody (Dako Cytomation, Glostrup, Denmark). With the assistance of image analysis software (Image Pro-Plus; Media Cybernetics, Washington, DC), radial alveolar count, according to Emery and Mithal (58), was measured in five areas of three lung sections from five fetuses per group. The largest two diameters of the transected, distended alveoli were measured and averaged. Ten peripheral arterioles with diameters ranging from 50 to 150 µ inform three lung sections from five fetuses per group were selected and the wall thickness in proportion to arterial width ((external diameter − luminal diameter) / external diameter) was measured.
Immunoblotting
TTF-1 protein levels were measured in homogenized lungs in cell disruption buffer (Protein Isolation System Paris, Ambion, TX). The protein content of 50 µg of tissue was measured using a protein assay kit (Pierce; BCA Protein Assay Kit, Rockford, IL). Western blotting was performed with 12% SDS-polyacrylamide gel with anti-TTF-1 antibody (1:6,000; Dako Cytomation, Glostrup, Denmark), and the values were normalized to anti-Cu/Zn superoxide-dismutase (1:1,000, Stressgen, MI).
mRNA Isolation and cDNA Synthesis
Total mRNA was isolated from frozen lungs using the High Pure RNA Tissue Kit (Roche Applied Science, Mannheim, Germany). Concentration and purity of RNAs were determined spectrophotometrically. RNA integrity was analyzed by 1% agarose gel electrophoresis; 1 μg of total RNAs was retrotranscripted to complementary DNAs (cDNAs) by reverse transcription reactions in a 100 μl vol using a High Capacity Reverse Transcription Kit (Applied Biosystems, Carlsbad, CA). All cDNAs were stored at −80 °C until further use.
Reverse-Transcriptase PCR (RT-PCR)
Fgf10 mRNA lung expression was quantified in a Light Cycler 480 with TaqMan probes (Universal Probe Library: no. 111, cat. no. 04693442001; Roche Diagnostics, Barcelona, Spain). Primers used were 5´-TGGAAATGGATACTGACACATTG-3´ and 5´-CTGCTGTTGCTGCTTGT-3´. All RT-PCR reactions were run in duplicate in a total reaction volume of 10 μl using 10 ng of cDNA obtained as described earlier. RT-PCR conditions were 95 °C for 10 min, followed by 45 cycles at 95 °C for 10 s, 60 °C for 30 s, and 72 °C for 1 s. Results were normalized to the expression of the 18S (Assay ID: HS99999901_S1; Applied Biosystems, Carlsbad, CA). The relative mRNA levels were determined by calculating the threshold cycle of each gene using the threshold cycle method.
In Vitro Lung Explant Studies
Embryos were recovered at E13 in sterile conditions and the tracheo–pulmonary bloc was transferred to culture Constar-Transwell cells (Corning, Madrid, Spain) with culture medium (DMEM/F12; Gibco, Barcelona, Spain) with 10% bovine fetal serum, 100 IU/ml penicillin, and 100 μg/ml streptomycin that was changed every 24 h. Incubation was done with 20.9% O2 and 5% CO2 at 37 °C for 72 h. Control (n = 55), Adria EA (n = 57), and Adria noEA (n = 17) lungs were studied. At 0, 24, 48, and 72 h of culture, the explants were examined under inverted microscope (Leica TCS SP5, Barcelona, Spain) at 37 °C and pictures were taken. Airway surface, perimeter, and number of terminal buds (59) were measured.
Variables
Lung weight/body weight (%), total lung DNA and proteins (μg), alveolar radial count (n), alveolar diameter (μm), arteriolar wall thickness (%) were determined as well as alveolar maturity according to the distribution of TTF-1 positive cells, TTF-1 protein, and late gestational control of branching (Fgf10 mRNA). Explant surface in mm2, airway perimeter in mm and number of terminal buds were also determined.
Statistical Methods
Mean values from the different groups were compared with nonparametric methods (Mann-Whitney for two groups and Kruskall–Wallis for more than two groups). When appropriate, parametric methods (ANOVA or Student’s t test) were preferred. The null hypothesis was rejected if the differences were >5% (P < 0.05).
Statement of Financial Support
This study was supported in part by IdiPAZ grants and by the Spanish Health Institute Carlos III (Grant N. RD08/0072: Maternal, Child Health and Development Network).
References
Spitz L . Oesophageal atresia. Orphanet J Rare Dis 2007;2:24.
Possögel AK, Diez-Pardo JA, Morales C, Navarro C, Tovar JA . Embryology of esophageal atresia in the adriamycin rat model. J Pediatr Surg 1998;33:606–12.
Dudley NE, Phelan PD . Respiratory complications in long-term survivors of oesophageal atresia. Arch Dis Child 1976;51:279–82.
Delius RE, Wheatley MJ, Coran AG . Etiology and management of respiratory complications after repair of esophageal atresia with tracheoesophageal fistula. Surgery 1992;112:527–32.
van Dooren M, Tibboel D, Torfs C . The co-occurrence of congenital diaphragmatic hernia, esophageal atresia/tracheoesophageal fistula, and lung hypoplasia. Birth Defects Res Part A Clin Mol Teratol 2005;73:53–7.
Mendelsohn C, Lohnes D, Décimo D, et al. Function of the retinoic acid receptors (RARs) during development (II). Multiple abnormalities at various stages of organogenesis in RAR double mutants. Development 1994;120:2749–71.
Minoo P, Su G, Drum H, Bringas P, Kimura S . Defects in tracheoesophageal and lung morphogenesis in Nkx2.1(-/-) mouse embryos. Dev Biol 1999;209:60–71.
Kim PC, Mo R, Hui Cc C . Murine models of VACTERL syndrome: Role of sonic hedgehog signaling pathway. J Pediatr Surg 2001;36:381–4.
Warburton D, Lee MK . Current concepts on lung development. Curr Opin Pediatr 1999;11:188–92.
Mahlapuu M, Enerbäck S, Carlsson P . Haploinsufficiency of the forkhead gene Foxf1, a target for sonic hedgehog signaling, causes lung and foregut malformations. Development 2001;128:2397–406.
Que J, Choi M, Ziel JW, Klingensmith J, Hogan BL . Morphogenesis of the trachea and esophagus: current players and new roles for noggin and Bmps. Differentiation 2006;74:422–37.
Boulet AM, Capecchi MR . Targeted disruption of hoxc-4 causes esophageal defects and vertebral transformations. Dev Biol 1996;177:232–49.
Diez-Pardo JA, Baoquan Q, Navarro C, Tovar JA . A new rodent experimental model of esophageal atresia and tracheoesophageal fistula: preliminary report. J Pediatr Surg 1996;31:498–502.
Ioannides AS, Chaudhry B, Henderson DJ, Spitz L, Copp AJ . Dorsoventral patterning in oesophageal atresia with tracheo-oesophageal fistula: Evidence from a new mouse model. J Pediatr Surg 2002;37:185–91.
Xia H, Migliazza L, Montedonico S, Rodriguez JI, Diez-Pardo JA, Tovar JA . Skeletal malformations associated with esophageal atresia: clinical and experimental studies. J Pediatr Surg 1999;34:1385–92.
Otten C, Migliazza L, Xia H, Rodriguez JI, Diez-Pardo JA, Tovar JA . Neural crest-derived defects in experimental esophageal atresia. Pediatr Res 2000;47:178–83.
Tovar JA . Stephen L. Gans Distinguished Overseas Lecture. The neural crest in pediatric surgery. J Pediatr Surg 2007;42:915–26.
Losada A, Tovar JA, Xia HM, Diez-Pardo JA, Santisteban P . Down-regulation of thyroid transcription factor-1 gene expression in fetal lung hypoplasia is restored by glucocorticoids. Endocrinology 2000;141:2166–73.
Bellusci S, Grindley J, Emoto H, Itoh N, Hogan BL . Fibroblast growth factor 10 (FGF10) and branching morphogenesis in the embryonic mouse lung. Development 1997;124:4867–78.
Ramasamy SK, Mailleux AA, Gupte VV, et al. Fgf10 dosage is critical for the amplification of epithelial cell progenitors and for the formation of multiple mesenchymal lineages during lung development. Dev Biol 2007;307:237–47.
Abler LL, Mansour SL, Sun X . Conditional gene inactivation reveals roles for Fgf10 and Fgfr2 in establishing a normal pattern of epithelial branching in the mouse lung. Dev Dyn 2009;238:1999–2013.
Hirashima T, Iwasa Y, Morishita Y . Mechanisms for split localization of Fgf10 expression in early lung development. Dev Dyn 2009;238:2813–22.
Hajduk P, Murphy P, Puri P . Fgf10 gene expression is delayed in the embryonic lung mesenchyme in the adriamycin mouse model. Pediatr Surg Int 2010;26:23–7.
Fewell JE, Hislop AA, Kitterman JA, Johnson P . Effect of tracheostomy on lung development in fetal lambs. J Appl Physiol 1983;55:1103–8.
Ein SH, Shandling B, Wesson D, Filler RM . Esophageal atresia with distal tracheoesophageal fistula: associated anomalies and prognosis in the 1980s. J Pediatr Surg 1989;24:1055–9.
Stark Z, Patel N, Clarnette T, Moody A . Triad of tracheoesophageal fistula-esophageal atresia, pulmonary hypoplasia, and duodenal atresia. J Pediatr Surg 2007;42:1146–8.
Toyama WM . Esophageal atresia and tracheoesophageal fistula in association with bronchial and pulmonary anomalies. J Pediatr Surg 1972;7:302–7.
Hemalatha V, Batcup G, Brereton RJ, Spitz L . Intrathoracic foregut cyst (foregut duplication) associated with esophageal atresia. J Pediatr Surg 1980;15:178–80.
Schwartz DL, So HB, Anban T, Creedon JJ . Total lung sequestration in association with oesophageal atresia and multiple tracheo-oesophageal fistulae. Z Kinderchir 1983;38:410–11.
Montedonico S, Díez-Pardo JA, Lassaletta L, Tovar JA . [Respiratory malformations associated with esophageal atresia]. Cir Pediatr 1999;12:61–4.
de Jong EM, Felix JF, Deurloo JA, et al. Non-VACTERL-type anomalies are frequent in patients with esophageal atresia/tracheo-esophageal fistula and full or partial VACTERL association. Birth Defects Res Part A Clin Mol Teratol 2008;82:92–7.
Usui N, Kamata S, Ishikawa S, et al. Anomalies of the tracheobronchial tree in patients with esophageal atresia. J Pediatr Surg 1996;31:258–62.
Filler RM, Messineo A, Vinograd I . Severe tracheomalacia associated with esophageal atresia: results of surgical treatment. J Pediatr Surg 1992;27:1136–40.
Corbally MT, Spitz L, Kiely E, Brereton RJ, Drake DP . Aortopexy for tracheomalacia in oesophageal anomalies. Eur J Pediatr Surg 1993;3:264–6.
Chetcuti P, Phelan PD . Respiratory morbidity after repair of oesophageal atresia and tracheo-oesophageal fistula. Arch Dis Child 1993;68:167–70.
Chetcuti P, Phelan PD, Greenwood R . Lung function abnormalities in repaired oesophageal atresia and tracheo-oesophageal fistula. Thorax 1992;47:1030–4.
Malmström K, Lohi J, Lindahl H, et al. Longitudinal follow-up of bronchial inflammation, respiratory symptoms, and pulmonary function in adolescents after repair of esophageal atresia with tracheoesophageal fistula. J Pediatr 2008;153:396–401.
Couriel JM, Hibbert M, Olinsky A, Phelan PD . Long term pulmonary consequences of oesophageal atresia with tracheo-oesophageal fistula. Acta Paediatr Scand 1982;71:973–8.
Somppi E, Tammela O, Ruuska T, et al. Outcome of patients operated on for esophageal atresia: 30 years’ experience. J Pediatr Surg 1998;33:1341–6.
Sistonen S, Malmberg P, Malmström K, et al. Repaired oesophageal atresia: respiratory morbidity and pulmonary function in adults. Eur Respir J 2010;36:1106–12.
Qi BQ, Merei J, Farmer P, et al. Tracheomalacia with esophageal atresia and tracheoesophageal fistula in fetal rats. J Pediatr Surg 1997;32:1575–9.
Xia H, Otten C, Migliazza L, Diez-Pardo JA, Tovar JA . Tracheobronchial malformations in experimental esophageal atresia. J Pediatr Surg 1999;34:536–9.
Dawrant MJ, Giles S, Bannigan J, Puri P . Adriamycin produces a reproducible teratogenic model of vertebral, anal, cardiovascular, tracheal, esophageal, renal, and limb anomalies in the mouse. J Pediatr Surg 2007;42:1652–8.
Qi BQ, Beasley SW . Communicating bronchopulmonary foregut malformations in the adriamycin-induced rat model of oesophageal atresia. Aust N Z J Surg 1999;69:56–9.
Qi BQ, Beasley SW . Pathohistological study of adriamycin-induced tracheal agenesis in the fetal rat. Pediatr Surg Int 1999;15:17–20.
Beasley SW, Diez Pardo J, Qi BQ, Tovar JA, Xia HM . The contribution of the adriamycin-induced rat model of the VATER association to our understanding of congenital abnormalities and their embryogenesis. Pediatr Surg Int 2000;16:465–72.
Qi BQ, Beasley SW, Williams AK . Evidence of a common pathogenesis for foregut duplications and esophageal atresia with tracheo-esophageal fistula. Anat Rec 2001;264:93–100.
Otcu S, Kaya M, Ozturk H, et al. Esophageal, tracheal and pulmonary parenchymal alterations in experimental esophageal atresia and tracheoesophageal fistula. A histological and morphometric study. Eur Surg Res 2002;34:405–10.
Karabulut B, Bostanci I, Tokat AO, et al. Pulmonary parenchymal alveolar histological study in experimental tracheo-oesophageal malformations. West Indian Med J 2008;57:20–3.
Hattori H, Okazaki S, Higuchi Y, Yoshibayashi M, Yamamoto T . Ectopic bronchus: an insufficiently recognized malformation causing respiratory morbidity in VATER association. Am J Med Genet 1999;82:140–2.
Bruch SW, Hirschl RB, Coran AG . The diagnosis and management of recurrent tracheoesophageal fistulas. J Pediatr Surg 2010;45:337–40.
Suen HC, Catlin EA, Ryan DP, Wain JC, Donahoe PK . Biochemical immaturity of lungs in congenital diaphragmatic hernia. J Pediatr Surg 1993;28:471–5.
Alfonso LF, Arnaiz A, Alvarez FJ, et al. Lung hypoplasia and surfactant system immaturity induced in the fetal rat by prenatal exposure to nitrofen. Biol Neonate 1996;69:94–100.
Gischler SJ, van der Cammen-van Zijp MH, Mazer P, et al. A prospective comparative evaluation of persistent respiratory morbidity in esophageal atresia and congenital diaphragmatic hernia survivors. J Pediatr Surg 2009;44:1683–90.
Biller JA, Allen JL, Schuster SR, Treves ST, Winter HS . Long-term evaluation of esophageal and pulmonary function in patients with repaired esophageal atresia and tracheoesophageal fistula. Dig Dis Sci 1987;32:985–90.
Robertson DF, Mobaireek K, Davis GM, Coates AL . Late pulmonary function following repair of tracheoesophageal fistula or esophageal atresia. Pediatr Pulmonol 1995;20:21–6.
Smith NP, Losty PD, Connell MG, Mayer U, Jesudason EC . Abnormal lung development precedes oligohydramnios in a transgenic murine model of renal dysgenesis. J Urol 2006;175:783–6.
Emery JL, Mithal A . The number of alveoli in the terminal respiratory unit of man during late intrauterine life and childhood. Arch Dis Child 1960;35:544–7.
Islam S, Narra V, Coté GM, Manganaro TF, Donahoe PK, Schnitzer JJ . Prenatal vitamin E treatment improves lung growth in fetal rats with congenital diaphragmatic hernia. J Pediatr Surg 1999;34:172–6.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Xiaomei, L., Aras-Lopez, R., Martinez, L. et al. Lung hypoplasia in rats with esophageal atresia and tracheo–esophageal fistula. Pediatr Res 71, 235–240 (2012). https://doi.org/10.1038/pr.2011.41
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/pr.2011.41
This article is cited by
-
Oesophageal atresia
Nature Reviews Disease Primers (2019)
-
Abnormal Sonic hedgehog signaling in the lung of rats with esophageal atresia induced by adriamycin
Pediatric Research (2014)
-
Abnormal control of lung branching in experimental esophageal atresia
Pediatric Surgery International (2013)
