
Abstract
Although it has long been known that patients with sepsis often have thrombocytopenia and that septic patients with severe thrombocytopenia have a poor prognosis and higher mortality, the role of platelets in the pathogenesis of sepsis is poorly understood. Here we report a protective role of platelets in septic shock. We show that experimental thrombocytopenia induced by intraperitoneal injection of an anti-glycoprotein Ibα monoclonal antibody increases mortality and aggravates organ failure, whereas transfusion of platelets reduces mortality in lipopolysaccharide-induced endotoxemia and a bacterial infusion mouse sepsis model. Plasma concentrations of proinflammatory cytokines TNF-α and IL-6 are elevated by thrombocytopenia and decreased by platelet transfusion in septic mice. Furthermore, we identify that platelets protect from septic shock by inhibiting macrophage-dependent inflammation via the COX1/PGE2/EP4-dependent pathway. Thus, these findings demonstrate a previously unappreciated role for platelets in septic shock and suggest that platelet transfusion may be effective in treating severely septic patients.
Similar content being viewed by others


Introduction
Platelets have a central role in physiological haemostasis by preventing excess bleeding, but are also involved in pathologic arterial thrombosis. Emerging evidence suggests that platelets are also critical components of the immune system1,2. Platelets are activated in patients with systemic inflammation and sepsis, resulting in their sequestration within microcirculation and thrombocytopenia3,4. Severe thrombocytopenia in septic patients is associated with adverse outcome and high mortality5,6,7. Platelets regulate inflammation and sepsis through multiple mechanisms. Platelets express a lipopolysaccharide (LPS) receptor, Toll-like receptor-4, which contributes to thrombocytopenia through a neutrophil-dependent pulmonary sequestration in response to LPS8,9,10. Platelets also interact with other leukocytes, including monocytes11,12. Interaction of activated platelets with monocytes induces nuclear translocation of nuclear factor-κB (NF-κB) and expression of NF-κB-dependent inflammatory genes13,14,15. In addition to direct interactions with leukocytes, platelets contribute to inflammation and immune progression by releasing cytokines and mediators stored in alpha and dense granules upon stimulation16,17.
In the present study, we used LPS-induced endotoxemia model and a bacterial infusion sepsis model through intraperitoneal (i.p.) injection of LPS or an Escherichia coli strain ATCC 25922, respectively, into mice to investigate the effects of experimental thrombocytopenia and platelet transfusion on septic shock. LPS or endotoxin, a component of the outer membrane of Gram-negative bacteria, has an essential role in the pathogenesis of sepsis. LPS administration into mice has become a standard inflammation model and is widely used in sepsis research18. Human sepsis is often caused by a single pathogen. The bacterial infusion model introduces a single pathogen into mice in a controlled manner, allowing reproducible infection, which has also been translated to larger animals for the study of systemic and organ-specific haemodynamics. We demonstrate that experimental thrombocytopenia increases mortality and aggravates organ failure, whereas transfusion of platelets reduces mortality in LPS-induced endotoxemia and E. coli ATCC 25922-induced sepsis. Our data reveal an important new role for platelets in sepsis and define a mechanism by which platelets protect from septic shock.
Results
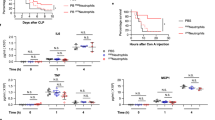
Thrombocytopenia exacerbates septic shock and organ failure
To establish a role of platelets in sepsis-associated inflammation, we induced thrombocytopenia in mice by i.p. injection of a rat antimouse GPIbα monoclonal antibody. Four hours after injection of the antibody, platelet counts were decreased by 90% (Fig. 1a). Platelet counts were not altered by injection of an isotype-matched rat IgG control. We then compared survival rates between the IgG-treated and thrombocytopenic mice after LPS challenge. Unexpectedly, thrombocytopenic mice had a significantly greater mortality rate than the mice administered with control IgG (Fig. 1b). All thrombocytopenic mice died within 36 h after LPS challenge. In contrast, none of the mice treated with control IgG died within 36 h after challenge by LPS. Lethality in sepsis is associated with organ failure. Thus, we examined the effects of thrombocytopenia on liver function in mice challenged with LPS. Plasma concentrations of liver enzymes, alanine aminotransferase and aspartate aminotransferase that are released into the circulation on injury and death of liver cells, respectively, were significantly higher in plasma from antimouse GPIbα monoclonal antibody-treated mice than those that received control IgG (Fig. 1c,d). Lactate dehydrogenase (LDH) is an enzyme found in many tissues, including the liver and heart, and may be released into plasma with hepatic and myocardial damage. Accordingly, plasma LDH concentration was higher in the thrombocytopenic mice than that of control mice (Fig. 1e). Creatine kinase (CK), an enzyme expressed by various tissues and cell types, can be elevated in plasma as a consequence of muscle injury or renal failure due to reduced clearance. Plasma CK concentrations were much higher in thrombocytopenic mice than in IgG-treated mice (Fig. 1f). Together, these results demonstrate that thrombocytopenia exacerbates tissue injury associated with sepsis.

(a) C57BL/6 mice were injected with 4 μg g−1 of body weight of a rat antimouse GPIbα monoclonal antibody (n=7) or control rat IgG (n=8) by i.p. Platelets counts were measured with a HEMAVET HV950FS multispecies haematology analyser before and 4 h after injection of antibody. Values are means±s.d. (b) Mice were then injected with LPS (10 mg kg−1). Survival rate was observed for up to 3 days. P-value was calculated by Wald’s test in a discrete time hazard model using version 9.2 of SAS software. (c–f) Eight hours after LPS treatment, plasma from mice receiving anti-GPIb antibody or IgG control were collected and alanine aminotransferase (ALT; c), aspartate aminotransferase (AST; d), LDH (e) and CK (f) concentrations in plasma were measured. Values are means±s.d. (n=4). Differences between two groups were assessed using unpaired two-tailed Student’s t-test. (g,h) Mice were injected i.p. with a rat antimouse GPIbα monoclonal antibody or rat IgG control. After 4 h, mice were injected i.p. with LPS (10 mg kg−1). Fifteen hours after LPS injection, mice were killed and the lungs were collected (g). Sections of lungs were stained with haematoxylin and eosin and images were captured. Scale bar, 0.5 mm (h).
Thrombocytopenia does not cause inflammatory haemorrhage
Previous studies reported that inflammation caused life-threatening haemorrhage during thrombocytopenia19,20. In those studies, the anti-GPIbα monoclonal antibody was injected intravenously or retro-orbitally, which resulted in a lowering of platelet count to <2.5% of that of control mice. However, no detectable haemorrhage occurred in the lungs (Fig. 1g,h) from mice in which thrombocytopenia was induced by i.p. injection of the anti-GPIbα antibody. In addition, red blood cell counts and haemoglobin concentrations were not significantly different between IgG-treated mice and the anti-GPIbα antibody-treated mice after LPS injection. Thus, inflammatory haemorrhage did not appear to account for the higher mortality rates with LPS in mice rendered thrombocytopenic by i.p. injection of the anti-GPIbα antibody.
Thrombocytopenia aggravates inflammatory response to LPS
Endotoxemia increases production of endogenous cytokines, including tumor necrosis factor-α (TNF-α) and interleukin (IL)-6, which have important roles in the development of disseminated intravascular coagulation, acute respiratory distress syndrome and septic shock21,22,23,24. Therefore, we investigated whether platelets protect from endotoxemia by altering expression of these cytokines. Indeed, LPS elevated plasma levels of TNF-α and IL-6, and the effect was amplified in thrombocytopenic mice (Fig. 2a,b). These results suggest that platelets normally inhibit endotoxemia-induced cytokine production. Macrophages are a major source of TNF-α in vivo22,25,26. Therefore, we hypothesize that platelets protect from septic shock by attenuating macrophage-dependent inflammatory response, thereby decreasing concentrations of these proinflammatory cytokines in plasma. We further hypothesize that platelet depletion increases TNF-α and IL-6 production by reducing a macrophage inhibitor factor(s). If these hypotheses are correct, then thrombocytopenia should have no effect on LPS-induced increases in plasma TNF-α and IL-6 levels in the mice lacking macrophages. Macrophages were depleted by retro-orbital injection of liposomal clodronate. Macrophage depletion was confirmed by detecting peripheral monocyte concentration by flow cytometry with an Alexa Fluor 488-labelled anti-CD115 monoclonal antibody. Monocyte concentration was reduced by>95% (Fig. 2c,d). As expected, plasma TNF-α concentrations were significantly decreased in macrophage-depleted mice challenged with LPS compared with the mice injected with control liposomes (Fig. 2e). The increase in plasma TNF-α observed in thrombocytopenic mice was absent when macrophages were predepleted.

(a,b) Mice were injected i.p. with a rat antimouse GPIbα monoclonal antibody or rat IgG control. After 4 h, the mice were injected i.p. with LPS (10 mg kg−1). Blood was collected from these mice before or at 1, 2.5, 4 and 8 h after LPS injection. TNF-α (a) and IL-6 (b) concentrations in plasma were measured by an enzyme-linked immunosorbent assay. Values are means±s.d. (n=6). Differences between two groups were assessed using unpaired two-tailed Student’s t-test. (c,d) Mice were injected retro-orbitally with clodronate (40 mg kg−1) or control liposome. Blood was collected from the mice at 24 h after injection of clodronate or liposome. Representative flow cytometry plots of monocyte subsets (c) and quantification of monocyte subsets (d) (n=3) are shown. Error bars indicate s.d. Difference was assessed using unpaired two-tailed Student’s t-test. (e,f) Mice were injected retro-orbitally with clodronate (40 mg kg−1) or control liposomes at 24 and 4 h before LPS injection. The mice were also injected i.p. with a rat antimouse GPIbα monoclonal antibody or rat IgG control at 4 h before LPS injection. Blood was collected before or at 1, 2, 4 and 8 h after LPS injection, and plasma TNF-α (e) and IL-6 (f) concentrations were measured. Values are means±s.d. (n=5). Differences between two groups were assessed using unpaired two-tailed Student’s t-test.
Macrophages do not appear to be a major source of IL-6 production early after LPS challenge, because depletion of macrophages did not alter plasma IL-6 concentrations until more than 4 h after injection of LPS (Fig. 2f). At 8 h after injection of LPS, depletion of macrophages decreased plasma IL-6 concentrations, suggesting that macrophages contribute to plasma IL-6 production at later stages of endotoxemia. Thrombocytopenia increased plasma IL-6 concentrations in control mice administrated liposomes but had no effect in mice predepleted with macrophages (Fig. 2f). These results demonstrate that platelets inhibit macrophage-derived IL-6 but have no effect on IL-6 production derived from other cells.
Thrombocytopenia exacerbates bacterial sepsis
We further investigated the role of platelets in septic shock using a bacterial infusion sepsis model. C57BL/6J mice were injected with the anti-GPIbα monoclonal antibody to deplete platelets. Four hours after injection of the antibody, the mice received i.p. 0.2 ml saline containing 2 × 107 colony-forming unit of E. coli ATCC 25922 per mouse. Thrombocytopenic mice had dramatically higher plasma levels of TNF-α and IL-6 (Fig. 3a,b), and a greater mortality rate compared with the mice administered control IgG (Fig. 3c).

(a,b) Mice were injected i.p. with a rat antimouse GPIbα monoclonal antibody or rat IgG control. After 4 h, the mice were injected i.p. with 0.2 ml saline containing 2 × 107 colony-forming unit (CFU) of E. coli ATCC 25922 per mouse. Blood was collected from these mice before or at 1, 2, 4 and 8 h after injection of bacteria. TNF-α (a) and IL-6 (b) concentrations in plasma were measured by enzyme-linked immunosorbent assays. Values are means±s.d. (n=6). Differences between two groups were assessed using unpaired two-tailed Student’s t-test. (c) C57BL/6 mice were injected i.p. with a rat antimouse GPIbα monoclonal antibody or control rat IgG (4 μg g−1 of body weight). After 4 h, the mice were injected with 0.2 ml saline containing 2 × 107 CFU of E. coli ATCC 25922. Survival rate was observed for up to 3 days. P-value was calculated by Wald’s test in a discrete time hazard model using version 9.2 of SAS software.
Platelet transfusion protects against septic shock
As platelets inhibited macrophage-derived TNF-α and IL-6 production, platelet transfusion may reduce in vivo inflammatory response during sepsis, thereby protecting from septic shock. C57BL/6J mice were injected retro-orbitally with washed platelets from the same background (1 × 109 per mouse) immediately after injection of LPS (i.p). Injection of 1 × 109 platelet per mouse increased platelet counts by ~25% (Fig. 4a). Plasma TNF-α and IL-6 concentrations following LPS were significantly lower in mice injected with platelets versus saline (Fig. 4b,c). Platelet transfusion also attenuated thrombocytopenia during endotoxemia (Fig. 4a). Platelet transfusion increased the survival rate from 16.0% to 41.7% (Fig. 4d) and 6.7% to 40.0% (Fig. 4e), respectively, in LPS-induced endotoxemia model and bacterial infusion sepsis model.

(a–c) Mice were injected with LPS i.p. (10 mg kg−1) followed by retro-orbital injection of 1 × 109 platelets in 0.2 ml saline or saline only. Blood were collected before or at 1, 2.5, 4 and 8 h after LPS injection. Platelet counts were measured before or at 1 and 8 h after LPS injection and expressed as relative to the counts of saline-treated mice before injection of LPS (a). Plasma TNF-α (b) and IL-6 (c) concentrations were measured. Values are means±s.d. (n=5). Differences between two groups were assessed using unpaired two-tailed Student’s t-test. (d) Mice were injected with LPS (10 mg kg−1), immediately followed by retro-orbital injection of 1 × 109 platelets in 0.2 ml saline or saline only. Survival rate was observed for 3 days. P-value was calculated by Wald’s test in a discrete time hazard model using version 9.2 of SAS software. (e) C57BL/6 mice were injected i.p. with 0.2 ml saline containing 2.5 × 107 colony-forming unit of E. coli ATCC 25922, immediately followed by retro-orbital injection of 1 × 109 platelets in 0.2 ml saline or saline only. Survival rate was observed for up to 3 days. P-value was calculated by Wald’s test in a discrete time hazard model using version 9.2 of SAS software. (f) Mice were injected retro-orbitally with clodronate or control liposomes at 24 and 4 h before LPS injection (5 mg kg−1 of body weight), immediately followed by retro-orbital injection of 1 × 109 platelets in 0.2 ml saline or saline only. Survival rate was observed for up to 3 days. P<0.001 between liposome group and clodronate group; P>0.2 between clodronate group injected with saline and clodronate group injected with platelets. P-value was calculated by Wald’s test in a discrete time hazard model using version 9.2 of SAS software.
Protection against sepsis by platelets depends on macrophages
To determine whether the protective effect of platelet transfusion on sepsis depends on inhibition of macrophage function, macrophages were depleted by retro-orbital injection of liposomal clodronate. As injection of control liposome increased mortality rate in the LPS-challenged mice, a lower dose of LPS was applied. Depletion of macrophage increased survival rate from 8.33% to 50.0% in response to LPS challenge (Fig. 4f). Transfusion of platelets did not significantly increase the survival rate in the macrophage-depleted mice. These results suggest that the protective effect of platelet transfusion on endotoxemia depends on macrophages.
Platelets inhibit macrophage-derived TNF-α and IL-6
Next, in-vitro experiments were performed to address whether platelets directly inhibit macrophage-derived cytokines in response to LPS. Mouse bone marrow-derived macrophages (BMDMs) were stimulated with LPS in the presence or absence of washed mouse platelets. Although addition of platelets increased TNF-α production from macrophages in response to a low-dose LPS (Fig. 5a), TNF-α production elicited by higher concentrations of LPS was dramatically reduced by addition of platelets (Fig. 5a). Similarly, IL-6 production induced by LPS was reduced by addition of platelets (Fig. 5a). As LPS at 100 ng ml−1 induced maximal TNF-α and IL-6 production from macrophages, this concentration of LPS was chosen to investigate the mechanism whereby platelets inhibit macrophage-derived TNF-α in the following experiments.

(a) LPS (1 ng ml−1~10 μg ml−1) was added with buffer or washed platelets (3 × 108 per ml) to BMDMs and incubated for 6 h. TNF-α and IL-6 concentrations in the supernatant were measured. (b) Washed platelets or medium were incubated with 3H-LPS (250 ng ml−1) at 37 °C for 2 h and centrifuged. Disintegrations per minute (DPM) in medium, platelet pellet or supernatant was measured. (c) LPS (100 ng ml−1) was added to BMDMs in the presence or absence of platelets (3 × 108 per ml) and incubated for 6 h. Washed platelets (3 × 108 per ml) were also incubated with LPS (100 ng ml−1) for 30 min or thrombin (Enzyme Research Laboratories, South Bend, IN, 0.1 U ml−1) in aggregometer at 37 °C for 5 min and centrifuged. The resulting supernatants with LPS were added to BMDMs and incubated for 6 h. LPS plus thrombin (Throm) were added to BMDMs and incubated for 6 h as a control. TNF-α and IL-6 concentrations in the supernatant were measured. (d) Washed platelets from Gq knockout mice or wild-type littermates were incubated with thrombin in aggregometer at 37 °C for 5 min and centrifuged. Supernatants plus LPS were added to BMDMs and incubated for 6 h. (e) Platelets (3 × 108 per ml) were pre-incubated with thrombin in aggregometer at 37 °C for 5 min and centrifuged. Serum amyloid A (SAA) (1 μg ml−1) together with buffer or supernatant from thrombin-stimulated platelets were added to BMDMs and incubated for 6 h. Values are means±s.d. (n=3 for all). Differences between two groups were assessed using unpaired two-tailed Student’s t-test.
Platelets express the LPS receptor, Toll-like receptor-4. To exclude the possibility that inhibition of LPS-induced TNF-α production is due to platelet sequestration of LPS, platelets were incubated with H3-labelled LPS for 2 h and the amount of radioactivity associated with platelets was measured. Despite the presence of LPS receptor on platelets, radioactivity remained in the supernatant rather than with platelets (Fig. 5b).
Platelet releasates inhibit TNF-α and IL-6 production
To determine whether platelets inhibit macrophage-derived cytokines through direct interaction between platelets and macrophages or by releasing inhibitory factors, platelets were incubated with LPS for 30 min and then centrifuged. The resulting supernatant containing LPS was added to macrophages. LPS in the presence of platelet releasate elicited less TNF-α and IL-6 production than did LPS alone (Fig. 5c).
The coagulation system is activated during sepsis, resulting in thrombin generation27,28, a physiological platelet agonist. Therefore, during sepsis, platelets are activated not only by LPS but also by physiological agonists such as thrombin. To determine whether platelet activation induced by a physiological agonist has an effect on macrophages, platelets from C57BL/6 mice were stimulated with thrombin and stirred in an aggregometer at 37 °C for 5 min. Supernatant from the thrombin-stimulated platelets markedly inhibited LPS-induced TNF-α and IL-6 production by macrophages (Fig. 5c). Thrombin-induced platelet activation and secretion requires Gq signalling29. As expected, supernatant of thrombin-stimulated wild-type platelets, but not Gq-deficient platelets, inhibited LPS-induced TNF-α production (Fig. 5d). Taken together, these results demonstrate an inhibitory role of platelet activation in LPS-induced TNF-α production by macrophages.
We next examined the effects of platelet releasate on TNF-α production from macrophages induced by serum amyloid A to determine whether platelet activation has a common role in the regulation of macrophage response to inflammatory challenge. Addition of supernatant of thrombin-stimulated platelets reduced serum amyloid A-stimulated TNF-α production by macrophages (Fig. 5e).
The effect of platelet secretion on TNF-α production
Platelets contain two major types of granules: α-granules containing soluble and membrane-associated proteins and dense granules containing small molecules such as nucleotides30. In response to agonist activation, including LPS, platelets secrete granule contents31,32,33. We therefore investigated whether granule releasate contributes to the inhibitory effects of platelets on macrophages. Most components secreted from α-granules are proteins16 that are sensitive to heating or proteinase K treatment. However, neither heating (Supplementary Fig. S1a) nor proteinase K (Supplementary Fig. S1b) treatment blunted the inhibitor effects of activated platelet supernatant on TNF-α production by macrophages.
ADP secreted from dense granules has an essential role in platelet activation through its receptor P2Y12 and P2Y1 (ref. 34). The addition of the ADP scavenger apyrase did not block the inhibitory effect of platelet releasate on LPS-induced TNF-α production by macrophages (Supplementary Fig. S2a). Furthermore, platelet releasate inhibited LPS-induced TNF-α from macrophages lacking the ADP receptor P2Y12 (Supplementary Fig. S2b), indicating that ADP is not required for the inhibition of macrophages by platelets. Munc13-4 is critical for cargo release from both α- and dense granules of platelets35. Platelet releasates from Unc13dJinx mice that lack Munc13-4 inhibited LPS-induced TNF-α by macrophages (Supplementary Fig. S2c). Taken together, these results suggest that inhibition of macrophage-derived TNF-α by platelets is independent of granule cargo release from platelets.
Platelets inhibit TNF-α through the COX1-dependent pathway
Synthesis of TXA2 from cyclooxygenase 1 (COX1) signalling is an important positive feedback mechanism for platelet activation. To determine whether COX1 is involved in platelet inhibition of macrophages, we examined whether aspirin, a COX1 inhibitor, could reverse the inhibitory effect of platelets on macrophages. Pretreatment of platelets with aspirin markedly reversed inhibition of LPS-induced TNF-α production from macrophages by platelet releasates (Fig. 6a). Accordingly, supernatant from thrombin-activated COX1-deficient platelets had less effect than that of wild-type platelets on inhibiting macrophage-derived TNF-α (Fig. 6b). Considering the importance of TXA2 in regulating platelet activation, we asked whether TXA2 synthesized from COX1 signalling is responsible for platelet inhibition on macrophages. Addition of a stable TXA2 analogue, U46619, failed to inhibit LPS-induced TNF-α production by macrophages (Supplementary Fig. S3a). Supernatant from thrombin-stimulated wild-type platelets inhibited LPS-induced TNF-α production by macrophages lacking the TXA2 receptor, TP (Supplementary Fig. S3b). Thus, platelet inhibition of macrophage function is likely to be independent of TXA2.

(a) Washed platelets were pre-incubated with aspirin (1 mM) or buffer for 5 min and then incubated with thrombin in an aggregometer at 37 °C for 5 min and centrifuged. LPS plus the supernatants of platelets were added to BMDMs and incubated for 6 h. Thrombin or thrombin plus aspirin were added to the wells with LPS in the absence of platelet supernatant as controls. (b) Platelets from COX1 knockout mice (COX1−/−) or wild-type littermates (COX1+/+) were incubated with buffer (resting) or thrombin in an aggregometer at 37 °C for 5 min. LPS plus the supernatants of platelets were added to BMDMs and incubated for 6 h. (c) Washed platelets from C57BL/6J mice were incubated with LPS for 30 min or thrombin in an aggregometer at 37 °C for 5 min. PGE2 in the supernatant were measured by an enzyme-linked immunosorbent assay (Cayman Chemical, Ann Arbor, MI). (d) BMDMs were pre-incubated with buffer, SC-51089 10 μM, AH 6809 10 μM, pertusis toxin (PTX) 0.5 μg ml−1, or GW627368X (GW62) at 37 °C for 15 min. BMDMs were then added with LPS or LPS plus supernatant from thrombin-activated platelets and incubated at 37 °C for 6 h. Values are means±s.d. (n=3 for all). Differences between two groups were assessed using unpaired two-tailed Student’s t-test.
COX1 activation results in the production of many types of prostaglandins. Prostaglandin E2 (PGE2) modulates macrophage function and decreases TNF-α and IL-6 production via its receptors EP1–4 (refs 36, 37). LPS and thrombin stimulated PGE2 production in platelets (Fig. 6c), and PGE2 or heated PGE2 inhibited LPS-induced TNF-α production in macrophages (Supplementary Fig. S3c). To determine whether PGE2 is required for platelet inhibition of LPS-induced TNF-α production by macrophages, macrophages were pre-incubated with antagonists of EP1–4. EP4 antagonist, GW62, but not the antagonists for EP1-3, reversed the inhibitory effect of platelet releasates on LPS-induced TNF-α production from macrophages (Fig. 6d), indicating that platelet inhibition of macrophage function involves the PGE2/EP4 pathway.
In agreement with the in-vitro observations, plasma TNF-α and IL-6 levels were higher in LPS-treated C57BL/6 mice injected with COX1-deficient platelets than with platelets from wild-type littermates (Fig. 7a,b). Injection of COX1-deficient platelets failed to protect from LPS-induced septic shock (Fig. 7c). Next, we investigated the role of aspirin on sepsis. We first determined the effective dose of aspirin in inhibiting platelet COX1 activity in vivo. Thrombin-induced TXA2 production was completely abolished in the platelets isolated from mice administrated with 10 or 25 mg kg−1 of aspirin (Fig. 7d). Aspirin (25 mg kg−1) pretreated mice had significantly higher plasma TNF-α and IL-6 concentrations than control mice (Fig. 7e,f). Aspirin pretreatment also reduced survival rate in the LPS-induced sepsis model (Fig. 7g). Taken together, these results demonstrate that platelet inhibition of inflammatory responses and LPS-induced TNF-α production from macrophages is dependent on platelet COX1 activity.

(a,b) C57BL/6 mice received i.p. injection of LPS (10 mg kg−1), immediately followed by retro-orbital injection of saline or 1 × 109 platelets from COX1-deficient mice (COX1−/−) or wild-type littermates (COX1+/+) in 0.2 ml saline. Blood was collected before or at 1, 2, 4 and 8 h after LPS injection. TNF-α (a) and IL-6 (b) concentrations in plasma were measured. Values are means±s.d. (n=5). Differences between two groups were assessed using unpaired two-tailed Student’s t-test. (c) C57BL/6 mice received i.p. injection of LPS (10 mg kg−1), immediately followed by retro-orbital injection of 1 × 109 platelets from either COX1 knockout mice or wild-type littermates in 0.2 ml saline or saline. Survival rate was observed for 3 days. P<0.001 between control group and the group injected with wild-type platelets; P>0.2 between control group and the group injected with COX1−/− platelets by Wald’s test. (d) C57BL/6 mice received i.p. injection of aspirin (10 or 25 mg kg−1). Blood was collected at 30 min after aspirin injection. Washed platelets were incubated with thrombin (0.1 U ml−1) for 5 min, and TXB2 in the supernatant was measured by enzyme-linked immunosorbent assay kit and expressed as relative to platelets from control mice (without aspirin injection) in the absence of thrombin. Values are means±s.d. (n=3). Differences between two groups were assessed using unpaired two-tailed Student’s t-test. (e,f) C57BL/6 mice received i.p. injection of aspirin (25 mg kg−1) 30 min before i.p. injection of LPS (10 mg kg−1). Blood was collected before or at 1, 2, 4 and 8 h after LPS injection. TNF-α (e) and IL-6 (f) concentrations in plasma were measured. Values are means±s.d. (n=6). Differences between two groups were assessed using unpaired two-tailed Student’s t-test. (g) C57BL/6 mice received i.p. injection of aspirin (25 mg kg−1) at 30 min before i.p. injection of LPS (10 mg kg−1). Survival rate was observed for 3 days. P-value was calculated by Wald’s test in a discrete time hazard model using version 9.2 of SAS software.
Discussion
Growing evidence indicates that platelets are key effectors in many inflammatory diseases. In sepsis, thrombocytopenia is a frequent complication and closely associates with increased mortality6,7. Whether thrombocytopenia is a marker for more disseminated disease or a contributor to poor outcomes is not known. In the present study, we demonstrate that antibody-induced thrombocytopenia in mice increases mortality in a LPS-induced endotoxemia and in a bacterial infusion sepsis model. Depletion of platelets worsened organ injury during septic shock, whereas transfusion of platelets significantly reduced mortality. These findings imply that platelets have a beneficial role in sepsis and protect against septic shock.
TNF-α has a key role in sepsis pathogenesis21,22,25. Platelets potentiate macrophage-derived TNF-α production in response to low-dose LPS (≤100 pg ml−1) under conditions where TNF-α increased only several fold over the baseline level38,39. Platelets also potentiate TNF-α production and inflammation in the mice injected with low-dose LPS40,41. Although we observed that platelets enhanced low-dose LPS (1 ng ml−1)-elicited TNF-α production, our findings demonstrate that platelets dramatically inhibit high-dose LPS (⩾10 ng ml−1)-induced TNF-α and IL-6 production by macrophages. Platelets also inhibited human macrophage-derived TNF-α production (Supplementary Fig. S4). Therefore, platelets appear to serve as a switch for the inflammatory response during sepsis in that they may promote inflammation at early stage of infection, but inhibit macrophage-dependent inflammation when plasma concentrations of LPS are high. Plasma LPS concentration, which can reach several ng ml−1 to over 100 μg ml−1, in severe sepsis carries prognostic significance. High LPS concentrations are associated with increased adverse outcome and higher mortality rate42,43.
Plasma IL-6 also predicts mortality in sepsis24,44. Although antibiotics decrease the overall mortality of septic mice, all animals with IL-6 levels in plasma >14.0 ng ml−1 die, regardless of whether they receive antibiotics24, suggesting that uncontrolled inflammation is an important cause of mortality. Interestingly, in the bacterial infusion sepsis model, we observed 100% mortality in control mice (injected with IgG) that had plasma IL-6 concentrations >14.0 ng ml−1, whereas all of the mice with plasma IL-6 concentrations <14.0 ng ml−1 survived. Plasma IL-6 levels in thrombocytopenic mice increased 10- and 29-fold over control animals (>63 and 176 ng ml−1, respectively) at 4 and 8 h after injection of bacteria. TNF-α concentrations in thrombocytopenic mice increased 77-fold over controls at 2 h after injection of bacteria. These data indicate that platelets have an essential role in modulating inflammation during sepsis, and that thrombocytopenia, which often occurs in severe septic patients, may be an important cause for the uncontrolled inflammation and death.
A previous study reported lethal lung haemorrhage in thrombocytopenic mice challenged with LPS19. In that study, the anti-GPIbα monoclonal antibody was injected intravenously, which resulted in a lowering of platelet count to <2.5% of that of control mice. They then performed platelet transfusion studies to determine the minimal concentration of platelets that was required for preventing inflammatory bleeding. Platelets from IL4Rα/GPIbα transgenic mice, which were resistant to the anti-GPIbα antibody, were injected into the inflamed mice. It was found that 10–15% of normal platelet counts was sufficient to prevent inflammatory bleeding19. In our study, thrombocytopenia was elicited by i.p. injection of an anti-GPIbα monoclonal antibody, which lowered platelet counts to ~10% of control. Consistent with previous studies, which reported that platelet counts of 10–15% normal were sufficient to prevent inflammatory bleeding19,45, no significant haemorrhage was observed in the lungs and brains from the thrombocytopenic mice. Therefore, in the present study, the increased mortality from sepsis in thrombocytopenic mice does not appear to be due to inflammatory haemorrhage, but rather to an exacerbated inflammatory response. To reconcile previous findings19,20 with our data, we induced thrombocytopenia in mice by retro-orbital injection of the anti-GPIbα antibody. In agreement with the previous studies, retro-orbital injection of the anti-GPIbα antibody induced much severe thrombocytopenia (platelet counts were reduced to <5% of that of control mice) and injection of LPS caused severe lung haemorrhage (Supplementary Fig. S5).
Our in-vitro and in vivo data suggest that the beneficial effect of platelets is likely to be due to COX1-dependent generation of PGE2 that acts on macrophage EP4 receptors. PGE2 is a key macrophage modulator36,46,47,48. LPS stimulates COX2 expression and PGE2 production in many cells by upregulating COX2 transcription through NF-κB-dependent pathways37,49,50. However, COX1 is the dominant isoform in platelets. Previous studies reported that pretreatment with non-steroidal anti-inflammatory drugs enhances endotoxin-induced production of TNF-α, IL-6 and IL-8 concentrations in the blood of human volunteers51,52. Consistent with these findings, aspirin pretreated mice had significantly higher plasma TNF-α and IL-6 concentrations and reduced survival rate than that of control mice in the LPS-induced sepsis model. COX metabolizes arachidonic acid to many biologically active eicosanoids such as prostaglandins, prostacyclins and thromboxane, and both COX1 and COX2 are activated during sepsis. However, they appear to have distinct roles in septic shock. Although aspirin reduced the survival rate in LPS-challenged mice, a COX2-specific inhibitor NS-398 increased the survival rate in LPS-induced endotoxemia53, suggesting that in contrast to COX1, COX2-derived products aggravate sepsis. Platelet-specific deletion of COX1 or the PGE2 synthase may be helpful in further evaluating the protective role of platelets in septic shock. Our data do suggest that other pathways may also be involved in platelet inhibition of macrophage function, because aspirin or COX1 deficiency only partially reversed platelet inhibition of LPS-induced TNF-α production by macrophages (Fig. 6a,b). In fact, following injection of COX1-deficient platelets, plasma TNF-α concentrations were lower, although to a lesser degree than that observed after injection of wild-type platelets (Fig. 7a). In this regard, a previous study showed that platelet-derived microparticles reduced the release of TNF-α by macrophages activated with LPS54.
Platelet transfusion has been recommended in septic patients with severe thrombocytopenia, in particular in patients with bleeding or in patients at risk for bleeding55. Historically, the purpose of platelet transfusion in sepsis is to halt or prevent bleeding. Our results suggest a potential beneficial effect of platelet transfusion in modulating immune response in sepsis, especially in the severe septic patients. Further studies of platelet transfusion in high-risk sepsis patients may be warranted.
Methods
Mice
Mice deficient in Gαq (ref. 29), P2Y12 (ref. 56), TP57, Munc 13-4 (Unc13dJinx mice)35, COX1 (ref. 58) and wild-type littermates from heterozygous breeding were used in this study. C57BL/6J mice were purchased from Jackson Laboratories. Mice were bred and maintained in the University of Kentucky Animal Care Facility, following institutional and National Institutes of Health guidelines after approval by the Institutional Animal Care and Use Committee. Eight- to 12-week-old males with a variety of genetic manipulations were used in most experiments.
Preparation of mouse platelets
Blood was collected from the abdominal aortas of isoflurane-anaesthetized mice (8–10 weeks) using 1/7 volume of ACD (85 mM trisodium citrate, 83 mM dextrose and 21 mM citric acid) as the anticoagulant59. Platelets were then washed once with CGS (0.12 M sodium chloride, 0.0129 M trisodium citrate and 0.03 MD-glucose, pH 6.5), resuspended in saline at 5 × 109 per ml for in vivo experiments or in RPMI 1640 medium containing 10 mM HEPES at 3 × 108 per ml for in-vitro experiments and incubated for 1 h at 22 °C before use.
Survival studies
LPS endotoxemia model: male mice (8–10 weeks old) were injected i.p. with 10 mg kg−1 LPS (0111:B4, purchased from Sigma) in a volume of 200 μl that was diluted in saline. Survival after LPS challenge was assessed every 12 h for 3 days. All survived mice were killed at the end of the third day.
Bacterial infusion sepsis model: E. coli ATCC 25922 was grown in Luria broth medium overnight. After washing once with sterile saline, the bacteria were resuspended and diluted in sterile saline. Male mice (8–10 weeks old) were injected i.p. with bacteria in a volume of 200 μl saline. Survival after injection of bacteria was assessed every 4 h for 3 days. All survived mice were killed at the end of the third day.
Platelet depletion and transfusion
Mice were injected i.p. with a commercially available rat antimouse GPIbα monoclonal antibody (Emfret Analytics, Wurzburg, Germany, 4 μg g−1 body weight) for platelet depletion. Mice were injected with same amount of isotype-matched rat IgG as controls. For platelet counts, mice were bled from the retro-orbital plexus under isoflurane anaesthesia (IsoFlo; Abbott Laboratories, Abbott Park, IL). Blood was collected into a heparin-containing tube and added with 10 mM (final concentration) EDTA as an anticoagulant. Platelet counts in whole blood were analysed with a HEMAVET HV950FS multispecies haematology analyser. For platelet transfusion, 1 × 109 washed platelets resuspended in 0.2 ml saline were injected into the retro-orbital plexus for each recipient mouse.
Macrophage depletion
Mice were injected retro-orbitally with 40 mg kg−1 of liposomal clodronate (L-α-Phosphatidyl-choline/cholesterol clodronate; Encapsula NanoSciences, Nashville, TN) at 24 and 4 h, respectively, before being challenged with LPS. The same amount of plain liposomes were injected as control. This method is specific for depletion of monocytes and macrophages, which undergo apoptosis following phagocytosis of the liposomal clodronate. Depletion of monocytes/macrophages was confirmed by flow cytometry analysis. EDTA anticoagulated blood was subjected to red blood cell lysis. White blood cells were resuspended in flow buffer (Hank’s balanced salt solution with 0.1% BSA and 5 mM EDTA) and incubated with Alexa Fluor 488-labelled anti-CD115 and APC-Cy7-labelled anti-CD45 antibodies.
Isolation of macrophages
Macrophages were isolated from mouse bone marrow and plated in a plastic flask60. Bone marrow macrophages were cultured for 7 days in the presence of 15% conditional medium from L929 cells.
Blood chemistry and cytokine measurements
Aspartate aminotransferase, alanine aminotransferase, CK and LDH were measured by Comparative Clinical Pathology Services (Columbia, MO). TNF-α and IL-6 were measured by ELISA assay (eBioscience, San Diego, CA). To observe the effects of platelets on TNF-α and IL-6 production in macrophages, washed platelets or medium were incubated with LPS (100 ng ml−1) for 30 min and added to 2 × 105 macrophages precoated in 96-well plates. To observe the effects of platelet releasates on TNF-α and IL-6 production in macrophages, washed platelets were incubated with LPS (100 ng ml−1) at room temperature for 30 min or thrombin (0.1 U ml−1) in an aggregometer with stirring at 1,000 r.p.m. at 37 °C for 5 min and then centrifuged at 2,500 r.p.m. at 22 °C for 3 min. Supernatants from LPS- or thrombin-stimulated platelets were added with LPS to 96-well plates precoated with 2 × 105 macrophages per well and incubated at 37 °C for 6 h.
Histologic examination
Mice were injected i.p. with an antimouse GPIbα monoclonal antibody or control IgG. After 4 h, LPS (10 mg kg−1) was injected i.p. Mice were anaesthetized and the lungs and brains were isolated at 15 h after injection of LPS. The lungs and brains were fixed with 4% paraformaldehyde in 0.1 m phosphate buffer (pH 7.4) for 2 days at 4 °C and then processed to paraffin embedding. Sections were cut at 5 μm and stained by haematoxylin and eosin. Images were captured with a Leica DM IRB microscope using × 20 objective (Leica Microsystems, Wetzlar, Germany).
Statistical analyses
The survival assay was analysed by Wald’s test in a discrete time hazard model using version 9.2 of SAS software. Significance in experiments comparing two groups was examined by unpaired two-tailed Student’s t-test. Values are reported as the mean±s.d. A value of P<0.05 was considered significant.
Additional information
How to cite this article: Xiang, B. et al. Platelets protect from septic shock by inhibiting macrophage-dependent inflammation via the cyclooxygenase 1 signalling pathway. Nat. Commun. 4:2657 doi: 10.1038/ncomms3657 (2013).
References
Semple, J. W., Italiano, J. E. Jr. & Freedman, J. Platelets and the immune continuum. Nat. Rev. Immunol. 11, 264–274 (2011).
Li, Z., Yang, F., Dunn, S., Gross, A. K. & Smyth, S. S. Platelets as immune mediators: their role in host defense responses and sepsis. Thromb. Res. 127, 184–188 (2011).
Russwurm, S. et al. Platelet and leukocyte activation correlate with the severity of septic organ dysfunction. Shock 17, 263–268 (2002).
Levi, M. Platelets in sepsis. Hematology 10, (Suppl 1): 129–131 (2005).
Akca, S. et al. Time course of platelet counts in critically ill patients. Crit. Care Med. 30, 753–756 (2002).
Sharma, B. et al. Thrombocytopenia in septic shock patients–a prospective observational study of incidence, risk factors and correlation with clinical outcome. Anaesth. Intens. Care 35, 874–880 (2007).
Moreau, D. et al. Platelet count decline: an early prognostic marker in critically ill patients with prolonged ICU stays. Chest 131, 1735–1741 (2007).
Andonegui, G. et al. Platelets express functional Toll-like receptor-4. Blood 106, 2417–2423 (2005).
Clark, S. R. et al. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat. Med. 13, 463–469 (2007).
Ma, A. C. & Kubes, P. Platelets, neutrophils, and neutrophil extracellular traps (NETs) in sepsis. J. Thromb. Haemost. 6, 415–420 (2008).
Evangelista, V. et al. Src family kinases mediate neutrophil adhesion to adherent platelets. Blood 109, 2461–2469 (2007).
Vieira-de-Abreu, A., Campbell, R. A., Weyrich, A. S. & Zimmerman, G. A. Platelets: versatile effector cells in hemostasis, inflammation, and the immune continuum. Semin. Immunopathol. 34, 5–30 (2012).
Weyrich, A. S., McIntyre, T. M., McEver, R. P., Prescott, S. M. & Zimmerman, G. A. Monocyte tethering by P-selectin regulates monocyte chemotactic protein-1 and tumor necrosis factor-alpha secretion. Signal integration and NF-kappa B translocation. J. Clin. Invest. 95, 2297–2303 (1995).
Weyrich, A. S. et al. Activated platelets signal chemokine synthesis by human monocytes. J. Clin. Invest. 97, 1525–1534 (1996).
Dixon, D. A. et al. Expression of COX-2 in platelet-monocyte interactions occurs via combinatorial regulation involving adhesion and cytokine signaling. J. Clin. Invest. 116, 2727–2738 (2006).
Blair, P. & Flaumenhaft, R. Platelet alpha-granules: basic biology and clinical correlates. Blood Rev. 23, 177–189 (2009).
Cloutier, N. et al. Platelets can enhance vascular permeability. Blood 120, 1334–1343 (2012).
Doi, K., Leelahavanichkul, A., Yuen, P. S. & Star, R. A. Animal models of sepsis and sepsis-induced kidney injury. J. Clin. Invest. 119, 2868–2878 (2009).
Goerge, T. et al. Inflammation induces hemorrhage in thrombocytopenia. Blood 111, 4958–4964 (2008).
Boulaftali, Y. et al. Platelet ITAM signaling is critical for vascular integrity in inflammation. J. Clin. Invest. 123, 908–916 (2013).
Tracey, K. J. & Cerami, A. Tumor necrosis factor: a pleiotropic cytokine and therapeutic target. Annu. Rev. Med. 45, 491–503 (1994).
Parameswaran, N. & Patial, S. Tumor necrosis factor-alpha signaling in macrophages. Crit. Rev. Eukaryot. Gene Expr. 20, 87–103 (2010).
Kruttgen, A. & Rose-John, S. Interleukin-6 in sepsis and capillary leakage syndrome. J. Interferon Cytokine Res. 32, 60–65 (2012).
Turnbull, I. R. et al. Antibiotics improve survival in sepsis independent of injury severity but do not change mortality in mice with markedly elevated interleukin 6 levels. Shock 21, 121–125 (2004).
Pfeffer, K. et al. Mice deficient for the 55 kd tumor necrosis factor receptor are resistant to endotoxic shock, yet succumb to L. monocytogenes infection. Cell 73, 457–467 (1993).
Flynn, J. L. et al. Tumor necrosis factor-alpha is required in the protective immune response against Mycobacterium tuberculosis in mice. Immunity 2, 561–572 (1995).
Esmon, C. T. et al. Inflammation, sepsis, and coagulation. Haematologica 84, 254–259 (1999).
Levi, M., van der Poll, T. & Schultz, M. Systemic versus localized coagulation activation contributing to organ failure in critically ill patients. Semin. Immunopathol. 34, 167–179 (2012).
Offermanns, S., Toombs, C. F., Hu, Y. H. & Simon, M. I. Defective platelet activation in G alpha(q)-deficient mice. Nature 389, 183–186 (1997).
Ren, Q., Ye, S. & Whiteheart, S. W. The platelet release reaction: just when you thought platelet secretion was simple. Curr. Opin. Hematol. 15, 537–541 (2008).
Shashkin, P. N., Brown, G. T., Ghosh, A., Marathe, G. K. & McIntyre, T. M. Lipopolysaccharide is a direct agonist for platelet RNA splicing. J. Immunol. 181, 3495–3502 (2008).
Cognasse, F. et al. Toll-like receptor 4 ligand can differentially modulate the release of cytokines by human platelets. Br. J. Haematol. 141, 84–91 (2008).
Zhang, G. et al. Lipopolysaccharide stimulates platelet secretion and potentiates platelet aggregation via TLR4/MyD88 and the cGMP-dependent protein kinase pathway. J. Immunol. 182, 7997–8004 (2009).
Kahner, B. N., Shankar, H., Murugappan, S., Prasad, G. L. & Kunapuli, S. P. Nucleotide receptor signaling in platelets. J. Thromb. Haemost. 4, 2317–2326 (2006).
Ren, Q. et al. Munc13-4 is a limiting factor in the pathway required for platelet granule release and hemostasis. Blood 116, 869–877 (2010).
Strassmann, G., Patil-Koota, V., Finkelman, F., Fong, M. & Kambayashi, T. Evidence for the involvement of interleukin 10 in the differential deactivation of murine peritoneal macrophages by prostaglandin E2. J. Exp. Med. 180, 2365–2370 (1994).
Wu, D. & Meydani, S. N. Mechanism of age-associated up-regulation in macrophage PGE2 synthesis. Brain. Behav. Immun. 18, 487–494 (2004).
Aiura, K. et al. Interaction with autologous platelets multiplies interleukin-1 and tumor necrosis factor production in mononuclear cells. J. Infect. Dis. 175, 123–129 (1997).
Scull, C. M., Hays, W. D. & Fischer, T. H. Macrophage pro-inflammatory cytokine secretion is enhanced following interaction with autologous platelets. J. Inflamm. (Lond.) 7, 53 (2010).
Aslam, R. et al. Platelet Toll-like receptor expression modulates lipopolysaccharide-induced thrombocytopenia and tumor necrosis factor-alpha production in vivo. Blood 107, 637–641 (2006).
Looney, M. R. et al. Platelet depletion and aspirin treatment protect mice in a two-event model of transfusion-related acute lung injury. J. Clin. Invest. 119, 3450–3461 (2009).
Opal, S. M. et al. Relationship between plasma levels of lipopolysaccharide (LPS) and LPS-binding protein in patients with severe sepsis and septic shock. J. Infect. Dis. 180, 1584–1589 (1999).
Behre, G. et al. Endotoxin concentration in neutropenic patients with suspected gram-negative sepsis: correlation with clinical outcome and determination of anti-endotoxin core antibodies during therapy with polyclonal immunoglobulin M-enriched immunoglobulins. Antimicrob. Agents Chemother. 36, 2139–2146 (1992).
Remick, D. G., Bolgos, G. R., Siddiqui, J., Shin, J. & Nemzek, J. A. Six at six: interleukin-6 measured 6 h after the initiation of sepsis predicts mortality over 3 days. Shock 17, 463–467 (2002).
Loria, G. D., Romagnoli, P. A., Moseley, N. B., Rucavado, A. & Altman, J. D. Platelets support a protective immune response to LCMV by preventing splenic necrosis. Blood 121, 940–950 (2013).
Xu, X. J., Reichner, J. S., Mastrofrancesco, B., Henry, W. L. Jr. & Albina, J. E. Prostaglandin E2 suppresses lipopolysaccharide-stimulated IFN-beta production. J. Immunol. 180, 2125–2131 (2008).
Sakamoto, A., Matsumura, J., Mii, S., Gotoh, Y. & Ogawa, R. A prostaglandin E2 receptor subtype EP4 agonist attenuates cardiovascular depression in endotoxin shock by inhibiting inflammatory cytokines and nitric oxide production. Shock 22, 76–81 (2004).
Nemeth, K. et al. Bone marrow stromal cells attenuate sepsis via prostaglandin E(2)-dependent reprogramming of host macrophages to increase their interleukin-10 production. Nat. Med. 15, 42–49 (2009).
D’Acquisto, F. et al. Involvement of NF-kappaB in the regulation of cyclooxygenase-2 protein expression in LPS-stimulated J774 macrophages. FEBS Lett. 418, 175–178 (1997).
Abate, A., Oberle, S. & Schroder, H. Lipopolysaccharide-induced expression of cyclooxygenase-2 in mouse macrophages is inhibited by chloromethylketones and a direct inhibitor of NF-kappa B translocation. Prostaglandins Other Lipid Mediat. 56, 277–290 (1998).
Martich, G. D., Danner, R. L., Ceska, M. & Suffredini, A. F. Detection of interleukin 8 and tumor necrosis factor in normal humans after intravenous endotoxin: the effect of antiinflammatory agents. J. Exp. Med. 173, 1021–1024 (1991).
Spinas, G. A., Bloesch, D., Keller, U., Zimmerli, W. & Cammisuli, S. Pretreatment with ibuprofen augments circulating tumor necrosis factor-alpha, interleukin-6, and elastase during acute endotoxinemia. J. Infect. Dis. 163, 89–95 (1991).
Reddy, R. C. et al. Selective inhibition of COX-2 improves early survival in murine endotoxemia but not in bacterial peritonitis. Am. J. Physiol. Lung Cell Mol. Physiol. 281, L537–L543 (2001).
Sadallah, S., Eken, C., Martin, P. J. & Schifferli, J. A. Microparticles (ectosomes) shed by stored human platelets downregulate macrophages and modify the development of dendritic cells. J. Immunol. 186, 6543–6552 (2011).
Zimmerman, J. L. Use of blood products in sepsis: an evidence-based review. Crit. Care Med. 32, S542–S547 (2004).
Foster, C. J. et al. Molecular identification and characterization of the platelet ADP receptor targeted by thienopyridine antithrombotic drugs. J. Clin. Invest. 107, 1591–1598 (2001).
Thomas, D. W. et al. Coagulation defects and altered hemodynamic responses in mice lacking receptors for thromboxane A2. J. Clin. Invest. 102, 1994–2001 (1998).
Langenbach, R. et al. Prostaglandin synthase 1 gene disruption in mice reduces arachidonic acid-induced inflammation and indomethacin-induced gastric ulceration. Cell 83, 483–492 (1995).
Zhang, G. et al. Distinct roles for Rap1b protein in platelet secretion and integrin alphaIIbbeta3 outside-in signaling. J. Biol. Chem. 286, 39466–39477 (2011).
Davies, J. Q. & Gordon, S. Isolation and culture of murine macrophages. Methods Mol. Biol. 290, 91–103 (2005).
Acknowledgements
This work was supported by the American Heart Association Midwest Affiliate Grant-in-Aid 0855698G (to Z.L.) and in part supported by the American Society of Hematology (ASH) bridge Grant Award and the NIH/National Center for Research Resources Centers of Biomedical Research Excellence in Obesity and Cardiovascular Disease Grant P20 RR021954. B.X. is a recipient of the American Heart Association Postdoctoral Fellowship Award. This work was support in part by resources provided by the Lexington VA Medical Center. We thank Dr. Richard Charnigo for his assistance with statistical analysis and Dr. Wendy S. Katz for her technical support for the histologic analysis. We thank Dr. Prabhakara R. Nagareddy and Judy F. Glass for their assistance with detection of monocyte depletion by flow cytometry.
Author information
Authors and Affiliations
Contributions
B.X. and Z.L. designed the research; Z.L. supervised the study; B.X. and G.Z. carried out most of the experiments; L.G. and X.-A.L. carried out some experiments of in vivo sepsis model; S.W.W. provided Jinx mice and helped with the analysis of secretion data; A.J.M. helped with PGE2 measurement; A.J.M., A.D. and S.S.S. provided extra technical assistance; S.S.S. provided valuable suggestions; B.X., A.D., S.S.S. and Z.L. wrote the manuscript. All authors commented on the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Information
Supplementary Figures S1-S5 (PDF 1378 kb)
Rights and permissions
About this article
Cite this article
Xiang, B., Zhang, G., Guo, L. et al. Platelets protect from septic shock by inhibiting macrophage-dependent inflammation via the cyclooxygenase 1 signalling pathway. Nat Commun 4, 2657 (2013). https://doi.org/10.1038/ncomms3657
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/ncomms3657
This article is cited by
-
EGFR of platelet regulates macrophage activation and bacterial phagocytosis function
Journal of Inflammation (2024)
-
TLR4 inhibitor alleviates sepsis-induced organ failure by inhibiting platelet mtROS production, autophagy, and GPIIb/IIIa expression
Journal of Bioenergetics and Biomembranes (2022)
-
Association of thrombocytopenia and infection in patients with ST-elevation myocardial infarction undergoing percutaneous coronary intervention
BMC Cardiovascular Disorders (2021)
-
Kupffer cell receptor CLEC4F is important for the destruction of desialylated platelets in mice
Cell Death & Differentiation (2021)
-
Role of platelet biomarkers in inflammatory response
Biomarker Research (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
