
Abstract
Designer receptors exclusively activated by designer drugs (DREADDs) represent a powerful chemogenetic technology for the remote control of neuronal activity and cellular signalling1,2,3,4. The muscarinic receptor-based DREADDs are the most widely used chemogenetic tools in neuroscience research. The Gq-coupled DREADD (hM3Dq) is used to enhance neuronal activity, whereas the Gi/o-coupled DREADD (hM4Di) is utilized to inhibit neuronal activity5. Here we report four DREADD-related cryogenic electron microscopy high-resolution structures: a hM3Dq–miniGq complex and a hM4Di–miniGo complex bound to deschloroclozapine; a hM3Dq–miniGq complex bound to clozapine-N-oxide; and a hM3R–miniGq complex bound to iperoxo. Complemented with mutagenesis, functional and computational simulation data, our structures reveal key details of the recognition of DREADD chemogenetic actuators and the molecular basis for activation. These findings should accelerate the structure-guided discovery of next-generation chemogenetic tools.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

Data availability
The structures of the hM3Dq–miniGq–DCZ, hM4Di–miniGo–DCZ, hM3Dq–miniGq–CNO, hM3R–miniGq–iperoxo and hM3R–iperoxo (local refinement) complexes have been deposited into the PDB (EMDB) databases under accession codes 8E9W (EMD-27966), 8E9X (EMD-27967), 8E9Y (EMD-27968), 8E9Z (EMD-27969) and 8EA0 (EMD-27970), respectively. The cryo-EM micrographs of hM3Dq–miniGq–DCZ, hM4Di–miniGo–DCZ, hM3Dq–miniGq–CNO and hM3R–miniGq–iperoxo have been deposited in the EMPIAR database (https://www.ebi.ac.uk/empiar/) with accession numbers EMPIAR-11185, EMPIAR-11184, EMPIAR-11193 and EMPIAR-11194, respectively. Source data are provided with this paper.
References
Urban, D. J. & Roth, B. L. DREADDs (designer receptors exclusively activated by designer drugs): chemogenetic tools with therapeutic utility. Annu. Rev. Pharmacol. Toxicol. 55, 399–417 (2015).
Roth, B. L. DREADDs for neuroscientists. Neuron 89, 683–694 (2016).
Roth, B. L. How structure informs and transforms chemogenetics. Curr. Opin. Struct. Biol. 57, 9–16 (2019).
Wang, L. et al. Use of DREADD technology to identify novel targets for antidiabetic drugs. Annu. Rev. Pharmacol. Toxicol. 61, 421–440 (2021).
Armbruster, B. N., Li, X., Pausch, M. H., Herlitze, S. & Roth, B. L. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc. Natl Acad. Sci. USA 104, 5163–5168 (2007).
Boyden, E. S., Zhang, F., Bamberg, E., Nagel, G. & Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 8, 1263–1268 (2005).
Isberg, V. et al. Generic GPCR residue numbers—aligning topology maps while minding the gaps. Trends Pharmacol. Sci. 36, 22–31 (2015).
Hu, J. et al. A G protein-biased designer G protein-coupled receptor useful for studying the physiological relevance of Gq/11-dependent signaling pathways. J. Biol. Chem. 291, 7809–7820 (2016).
Nakajima, K. & Wess, J. Design and functional characterization of a novel, arrestin-biased designer G protein-coupled receptor. Mol. Pharmacol. 82, 575–582 (2012).
Guettier, J. M. et al. A chemical-genetic approach to study G protein regulation of β cell function in vivo. Proc. Natl Acad. Sci. USA 106, 19197–19202 (2009).
Inoue, A. et al. Illuminating G-protein-coupling selectivity of GPCRs. Cell 177, 1933–1947.e25 (2019).
Bender, D., Holschbach, M. & Stöcklin, G. Synthesis of n.c.a. carbon-11 labelled clozapine and its major metabolite clozapine-N-oxide and comparison of their biodistribution in mice. Nucl. Med. Biol. 21, 921–925 (1994).
Gomez, J. L. et al. Chemogenetics revealed: DREADD occupancy and activation via converted clozapine. Science 357, 503–507 (2017).
Jann, M. W., Lam, Y. W. & Chang, W. H. Rapid formation of clozapine in guinea-pigs and man following clozapine-N-oxide administration. Arch. Int. Pharmacodyn. Ther. 328, 243–250 (1994).
Roth, B. L., Sheffler, D. J. & Kroeze, W. K. Magic shotguns versus magic bullets: selectively non-selective drugs for mood disorders and schizophrenia. Nat. Rev. Drug Discov. 3, 353–359 (2004).
Weston, M. et al. Olanzapine: a potent agonist at the hM4D(Gi) DREADD amenable to clinical translation of chemogenetics. Sci. Adv. 5, eaaw1567 (2019).
Nagai, Y. et al. Deschloroclozapine, a potent and selective chemogenetic actuator enables rapid neuronal and behavioral modulations in mice and monkeys. Nat. Neurosci. 23, 1157–1167 (2020).
Thompson, K. J. et al. DREADD agonist 21 is an effective agonist for muscarinic-based DREADDs in vitro and in vivo. ACS Pharmacol. Transl. Sci. 1, 61–72 (2018).
Chen, X. et al. The first structure–activity relationship studies for designer receptors exclusively activated by designer drugs. ACS Chem. Neurosci. 6, 476–484 (2015).
Bonaventura, J. et al. High-potency ligands for DREADD imaging and activation in rodents and monkeys. Nat. Commun. 10, 4627 (2019).
Nehme, R. et al. Mini-G proteins: novel tools for studying GPCRs in their active conformation. PLoS ONE 12, e0175642 (2017).
Kim, K. et al. Structure of a hallucinogen-activated Gq-coupled 5-HT2A serotonin receptor. Cell 182, 1574–1588.e19 (2020).
Garcia-Nafria, J., Nehme, R., Edwards, P. C. & Tate, C. G. Cryo-EM structure of the serotonin 5-HT1B receptor coupled to heterotrimeric Go. Nature 558, 620–623 (2018).
Zhang, S. et al. Inactive and active state structures template selective tools for the human 5-HT5A receptor. Nat. Struct. Mol. Biol. 29, 677–687 (2022).
Duan, J. et al. Cryo-EM structure of an activated VIP1 receptor–G protein complex revealed by a NanoBiT tethering strategy. Nat. Commun. 11, 4121 (2020).
Maeda, S., Qu, Q., Robertson, M. J., Skiniotis, G. & Kobilka, B. K. Structures of the M1 and M2 muscarinic acetylcholine receptor/G-protein complexes. Science 364, 552–557 (2019).
Wang, J. et al. The unconventional activation of the muscarinic acetylcholine receptor M4R by diverse ligands. Nat. Commun. 13, 2855 (2022).
Liu, H. et al. Structure-guided development of selective M3 muscarinic acetylcholine receptor antagonists. Proc. Natl Acad. Sci. USA 115, 12046–12050 (2018).
Thorsen, T. S., Matt, R., Weis, W. I. & Kobilka, B. K. Modified T4 lysozyme fusion proteins facilitate G protein-coupled receptor crystallogenesis. Structure 22, 1657–1664 (2014).
Kruse, A. C. et al. Structure and dynamics of the M3 muscarinic acetylcholine receptor. Nature 482, 552–556 (2012).
Kenakin, T., Watson, C., Muniz-Medina, V., Christopoulos, A. & Novick, S. A simple method for quantifying functional selectivity and agonist bias. ACS Chem. Neurosci. 3, 193–203 (2012).
Wess, J., Maggio, R., Palmer, J. R. & Vogel, Z. Role of conserved threonine and tyrosine residues in acetylcholine binding and muscarinic receptor activation. A study with M3 muscarinic receptor point mutants. J. Biol. Chem. 267, 19313–19319 (1992).
Heitz, F. et al. Site-directed mutagenesis of the putative human muscarinic M2 receptor binding site. Eur. J. Pharmacol. 380, 183–195 (1999).
Nawaratne, V. et al. New insights into the function of M4 muscarinic acetylcholine receptors gained using a novel allosteric modulator and a DREADD (designer receptor exclusively activated by a designer drug). Mol. Pharmacol. 74, 1119–1131 (2008).
Abdul-Ridha, A., Lane, J. R., Sexton, P. M., Canals, M. & Christopoulos, A. Allosteric modulation of a chemogenetically modified G protein-coupled receptor. Mol. Pharmacol. 83, 521–530 (2013).
Haga, K. et al. Structure of the human M2 muscarinic acetylcholine receptor bound to an antagonist. Nature 482, 547–551 (2012).
Jumper, J. et al. Highly accurate protein structure prediction with AlphaFold. Nature 596, 583–589 (2021).
McCorvy, J. D. et al. Structural determinants of 5-HT2B receptor activation and biased agonism. Nat. Struct. Mol. Biol. 25, 787–796 (2018).
Wacker, D., Stevens, R. C. & Roth, B. L. How ligands illuminate GPCR molecular pharmacology. Cell 170, 414–427 (2017).
Suno, R. et al. Structural insights into the subtype-selective antagonist binding to the M2 muscarinic receptor. Nat. Chem. Biol. 14, 1150–1158 (2018).
Flock, T. et al. Universal allosteric mechanism for Gα activation by GPCRs. Nature 524, 173–179 (2015).
Xia, R. et al. Cryo-EM structure of the human histamine H1 receptor/Gq complex. Nat. Commun. 12, 2086 (2021).
Cao, C. et al. Structure, function and pharmacology of human itch GPCRs. Nature 600, 170–175 (2021).
Mobbs, J. I. et al. Structures of the human cholecystokinin 1 (CCK1) receptor bound to Gs and Gq mimetic proteins provide insight into mechanisms of G protein selectivity. PLoS Biol. 19, e3001295 (2021).
Yin, Y. L. et al. Molecular basis for kinin selectivity and activation of the human bradykinin receptors. Nat. Struct. Mol. Biol. 28, 755–761 (2021).
Blin, N., Yun, J. & Wess, J. Mapping of single amino acid residues required for selective activation of Gq/11 by the m3 muscarinic acetylcholine receptor. J. Biol. Chem. 270, 17741–17748 (1995).
Xu, P. et al. Structural insights into the lipid and ligand regulation of serotonin receptors. Nature 592, 469–473 (2021).
Kooistra, A. J. et al. GPCRdb in 2021: integrating GPCR sequence, structure and function. Nucleic Acids Res. 49, D335–D343 (2021).
Wang, Y. et al. Molecular recognition of an acyl-peptide hormone and activation of ghrelin receptor. Nat. Commun. 12, 5064 (2021).
Zhang, X. et al. Structures of the human cholecystokinin receptors bound to agonists and antagonists. Nat. Chem. Biol. 17, 1230–1237 (2021).
Peck, J. V., Fay, J. F. & Strauss, J. D. High-speed high-resolution data collection on a 200 keV cryo-TEM. IUCrJ 9, 243–252 (2022).
Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. J. Struct. Biol. 152, 36–51 (2005).
Bepler, T., Kelley, K., Noble, A. J. & Berger, B. Topaz-Denoise: general deep denoising models for cryoEM and cryoET. Nat. Commun. 11, 5208 (2020).
Punjani, A., Rubinstein, J. L., Fleet, D. J. & Brubaker, M. A. cryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nat. Methods 14, 290–296 (2017).
Punjani, A., Zhang, H. & Fleet, D. J. Non-uniform refinement: adaptive regularization improves single-particle cryo-EM reconstruction. Nat. Methods 17, 1214–1221 (2020).
Rosenthal, P. B. & Henderson, R. Optimal determination of particle orientation, absolute hand, and contrast loss in single-particle electron cryomicroscopy. J. Mol. Biol. 333, 721–745 (2003).
Heymann, J. B. & Belnap, D. M. Bsoft: image processing and molecular modeling for electron microscopy. J. Struct. Biol. 157, 3–18 (2007).
Sanchez-Garcia, R. et al. DeepEMhancer: a deep learning solution for cryo-EM volume post-processing. Commun. Biol. 4, 874 (2021).
Pettersen, E. F. et al. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D 60, 2126–2132 (2004).
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D 66, 213–221 (2010).
Chen, V. B. et al. MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr. D 66, 12–21 (2010).
Olsen, R. H. J. et al. TRUPATH, an open-source biosensor platform for interrogating the GPCR transducerome. Nat. Chem. Biol. 16, 841–849 (2020).
Lomize, M. A., Pogozheva, I. D., Joo, H., Mosberg, H. I. & Lomize, A. L. OPM database and PPM web server: resources for positioning of proteins in membranes. Nucleic Acids Res. 40, D370–D376 (2012).
Brooks, B. R. et al. CHARMM: the biomolecular simulation program. J. Comput. Chem. 30, 1545–1614 (2009).
Jo, S., Kim, T., Iyer, V. G. & Im, W. CHARMM-GUI: a web-based graphical user interface for CHARMM. J. Comput. Chem. 29, 1859–1865 (2008).
Lee, J. et al. CHARMM-GUI input generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM simulations using the CHARMM36 additive force field. J. Chem. Theory Comput. 12, 405–413 (2016).
Wu, E. L. et al. CHARMM-GUI membrane builder toward realistic biological membrane simulations. J. Comput. Chem. 35, 1997–2004 (2014).
Huang, J. et al. CHARMM36m: an improved force field for folded and intrinsically disordered proteins. Nat. Methods 14, 71–73 (2017).
Klauda, J. B. et al. Update of the CHARMM all-atom additive force field for lipids: validation on six lipid types. J. Phys. Chem. B 114, 7830–7843 (2010).
Case, D. A. et al. AMBER v.2020 (Univ. of California, San Francisco, 2020).
Roe, D. R. & Cheatham, T. E. PTRAJ and CPPTRAJ: software for processing and analysis of molecular dynamics trajectory data. J. Chem. Theory Comput. 9, 3084–3095 (2013).
Humphrey, W., Dalke, A. & Schulten, K. VMD: visual molecular dynamics. J. Mol. Graphics 14, 33–38 (1996).
Pettersen, E. F. et al. UCSF ChimeraX: structure visualization for researchers, educators, and developers. Protein Sci. 30, 70–82 (2021).
Acknowledgements
This work was supported by NIH grant U24DK116194, the NIMH Psychoactive Drug Screening Program and the Michael Hooker Distinguished Professorship to B.L.R. We thank J. Peck and J. Strauss of the UNC CryoEM Core Facility for their technical assistance in this project. The Titan X Pascal used for this research was donated to J.F.F. by the NVIDIA Corporation.
Author information
Authors and Affiliations
Contributions
S.Z. designed the experiments, performed the cloning, expression and purification of transducer-coupled DREADD or WT complexes, built and refined the models, performed mutagenesis and functional studies, performed the binding and ELISA assays, and prepared the figures, tables and manuscript. R.H.G. performed and analysed the docking and MD simulations. X.-P.H. performed cAMP Glo-Sensor and calcium flux experiments. Y.L. assisted in the functional experiments. B.E.K. and C.C. assisted in the protein expression experiments. J.F.F. made the grids and collected and processed the cryo-EM data. B.L.R supervised the entire project and prepared the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Beili Wu and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Schematic diagrams of the constructs used in this study.
a-b, Modified snake diagrams of hM3Dq and hM4Di from the GPCRdb including the N-terminal and ICL3 truncation sites. The LgBit was fused at the C terminus of the hM3Dq receptor. DREADD mutation sites are indicated by red circles. Residues in yellow circles were removed from the expression constructs. c, Schematic diagram of the 3-in-1 vector (pFastDual plasmid-based) for the expression of miniGq or miniGo. The HiBit was fused at the C-terminus of the Gβ subunit. d-e, The activities of intact and ICL3 deletions of hM3Dq (d) or hM4Di (e) under the stimulation by DCZ, respectively. See Supplementary Table 4 for fitted parameter values that represent the mean ± SEM of n = 4 biologically independent experiments. f-g, Surface expression levels of intact and ICL3 deletions of hM3Dq and hM4Di. The WT hM3R and hM4R are also included as controls. Data represent the mean ± SEM of n = 3 biologically independent experiments. See Supplementary Table 11 for the details.
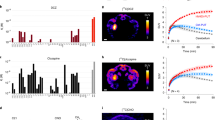
Extended Data Fig. 2 CryoEM analysis for the complexes of DCZ-bound hM3Dq-miniGq, DCZ-bound hM4Di-miniGo, CNO-bound hM3Dq-miniGq, and iperoxo-bound hM3R-miniGq.
For each of the respective complexes are shown: a, Selected frame aligned micrograph. The experiment was repeated three times with similar results. b, Histograms of defocus values for micrographs used in the single-particle analysis (see Extended Data Table 1 for more details). c, Orientational distribution heat map. d, 2D plots of the gold-standard Fourier shell correlation (GSFSC) between half maps (black) and FSC between model and the b-factor sharpened map for respective refined model (red) as calculated by phenix.mtirage. Local refinement GSFSC is shown in gray and the locally refined map to the model is shown in plumb. e, Local resolution heat-map calculated using the local windowed FSC method.
Extended Data Fig. 3 Overall structure and CHS binding site of the iperoxo bound hM3R-miniGq complex.
a, Superposition of the overall structures of the iperoxo-bound hM3R-miniGq and DCZ-bound hM3Dq-miniGq. The density map of the iperoxo is shown as gray meshes. b-c, The extracellular (b) and intracellular (c) views of structural comparison of the receptors between hM1R-G11 (PDB: 6OIJ) and hM3Dq-miniGq complexes, respectively. d-e, The extracellular (d) and intracellular (e) views of structural comparison of the receptors between hM3Dq-miniGq and hM4Di-miniGo complexes, respectively. f, Comparison of the interfaces between TM5 of hM1R or hM3Dq and α5 helix of G11 or miniGq subunit in the structures of hM1R-G11 or hM3Dq-miniGq complex. g, Three views of the superposition of hM1R (6OIJ) and hM3R structures. The CHS molecules in the hM1R or hM3R structure are shown in gray or yellow spheres, respectively.
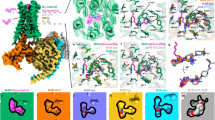
Extended Data Fig. 4 Ligand binding pockets in DREADD and WT receptors.
a, Side view of the binding pocket of the hM4Di-DCZ structure. Both DCZ and surrounding residues interacting with DCZ are shown in stick models. b, Interactions of the 8-chloro group of CNO with surrounding residues in the CNO-hM3Dq structure. The compound CNO is shown in both stick and sphere models and DCZ and surrounding residues are shown in the ball-and-stick model. c, Superposition of the binding pockets of hM3R-iperoxo, hM1R-iperoxo (6OIJ), and hM2R-iperoxo (6OIK) structures. The inset shows the structural comparison of the iperoxo molecules in the binding pockets of these three structures. Hydrogen bonds are indicated as dashed lines. d, Side view of the binding pockets of superposed hM3Dq-DCZ and hM3R-iperoxo structures. The deeper subpocket occupied by the heterocyclic group of iperoxo is formed by N3x37, V3x40, F5x47, W6x48, and N6x52. e-f, Structural comparisons of the hM3Dq-DCZ and rM3R-tiotropium (TIO, 4U15) structures (e), and the hM4Di-DCZ and hM2R-QNB (PDB: 3UON) structures (f). Major conformational differences of the TM4, TM5, and TM6 on the extracellular side and ligands are indicated by red and blue arrows, respectively. Ligands are shown in stick models. g, Interactions between DCZ or TIO and Y/C3x33, Y6x51, and N6x52 in hM3Dq or rM3R structure. The Y3x33 is shown in spheres. The conformational changes of the ligands and residues are indicated by gray and red arrows, respectively. h, Top view of the binding pocket comparison of predicted hM3R structure from the alphafold2 (AF2) server, modeled hM3Dq structure by alphafold2 server, and hM3R in the structure of the iperoxo-bound hM3R-miniGq complex.
Extended Data Fig. 5 Functional validations of iperoxo and QNB towards the WT or DREADD receptors.
a—d, Binding affinities of iperoxo towards the WT and DREADD receptors. See Supplementary Table 5 for fitted parameter values that represent the mean ± SEM of n = 3 biologically independent experiments. e-f, Iperoxo, ACh, and DCZ agonist activities toward DREADD receptors by BRET2 assay. See Supplementary Table 6 for fitted parameter values that represent the mean ± SEM of n = 4 biologically independent experiments. g-h, Iperoxo, ACh, and DCZ agonist activities toward DREADD receptors by calcium flux assay (g) and cAMP Glo-sensor assay (h). See Supplementary Table 7 for fitted parameter values that represent the mean ± SEM of n = 3–5 biologically independent experiments. i-l, Gq activities of hM3R (i) and hM3Dq (j) under the stimulation of compounds ACh, DCZ, QNB, TIO, and 384 by FLIPR calcium assay and Gi/o activities of hM4R (k) and hM4Di (l) under the stimulation of compounds ACh, DCZ, QNB, tiotropium (TIO), AD-DX 384 (384) by split-luciferase-based cAMP Glo-sensor assay. See Supplementary Table 8 for fitted parameter values that represent mean ± SEM of n = 3 biologically independent experiments. m-p, Mutagenesis analysis of three arginines (R177, R184, and R552) in the ACh-induced activation for hM3R (m-n) and DCZ-induced activation for hM3Dq (o-p) by the BRET2 assay, respectively. See Supplementary Table 9 for fitted parameter values that represent mean ± SEM of n = 4 biologically independent experiments.
Extended Data Fig. 6 MD simulations of iperoxo and DCZ in WT and DREADD receptors.
a, Activation of hM4R and hM4R (A203G) under the stimulation of iperoxo by the BRET assay. See Supplementary Table 10 for fitted parameter values that represent mean ± SEM of n = 4 biologically independent experiments. b-c, Basal activities of hM3Dq (b) and hM4Di (c) by BRET2 assay. Data represent the mean ± SEM. ***p = 0.0004 (hM3Dq, n = 6), ****p < 0.0001 (hM4Di, n = 6), unpaired two-tailed student t-test. d-e, Interaction between W4x57 and N3x37 at the start and end snapshots of the simulation of hM3Dq-iperoxo (d) and hM3R-iperoxo (e). f-g, Representative poses of iperoxo (f) and DCZ (g) in the hM3R (A5x461G) and hM3Dq (G5x461A) during the MD simulation, respectively. h—m, Statistics analysis of the iperoxo-N153 distance (isoxazole oxygen of iperoxo to ND2 of N153) and W200-N153 distance (indole N of W200 to OD1 of N153) in the simulation of hM3R-iperoxo (h and i), hM3Dq-iperoxo (j and k), and hM3R (A5x461G)-iperoxo (l and m), respectively. n-o, Statistics analysis of the DCZ-D148 distance [nitrogen on the piperazine ring (N02) of DCZ to OD2 of D148] in the simulation of hM3Dq-DCZ (n) and hM3Dq (G5x461A)-DCZ (o), respectively. The x-axis represents the binned distance and the y-axis is the count of frames from the simulations. Each color represents separate runs from the respective simulations.
Extended Data Fig. 7 Docking poses of clozapine in inactive rM3R and olanzapine in hM3Dq structures.
a, Chemical structures of JHU 37152 (J52), JHU 37160 (J60), compound 6 from the previous SAR study, and AF-DX 384 (384). b-c, Two views of the superposition of the preferred docking pose of clozapine (CLZ) and binding pose of tiotropium (TIO) in the inactive state rM3R structure (4DAJ). d, Superposition of the preferred docking pose of the olanzapine (OLP) and binding pose of DCZ in the hM3Dq structure. The conformational differences from DCZ to OLP are indicated by red arrows.
Extended Data Fig. 8 Receptor-Gq protein interfaces in different complexes.
a—h Interfaces mediated by the feature residue EH5.22 (a—d) and NH5.24 (e—h) in the Gq protein engaged H1R (a, e), CCK1R (b, f), B1R (c, g), and MRGPRX4R (d, h) complexes, respectively.
Extended Data Fig. 9 Two distinct Gq coupling modes.
a-b, Comparison of the receptor-G protein interfaces mediated by the EH5.22 (a) and NH5.24 (b) in the 5-HT2A-miniGq and hM3Dq-miniGq structures, respectively. c, Conformational changes of the TM6 of the receptor and α5 helix of the Gq subunit among the 5-HT2AR-miniGq (6WHA), hM1R-G11 (6OIJ), hM3Dq-miniGq, and H1R-miniGq (7DFL) structures. d, Structural comparison of the Gq proteins in the complexes of receptors 5-HT2AR (6WHA), B1R (7EIB), B2R (7F2O), ghrelin receptor (7F9Y), CCK1R (7MBY), CCK2R (7F8W), MRGPRX2 (7S8L), MRGPRX4 (7S8P), hM1R (6OIJ), hM3Dq, and H1R (7DFL). e, Sequence alignment of α5 helices of all Gα subunits. Residues involved in the interactions with TM5 of hM1R are indicated by the blue arrows. And two feature residues in Gq/11 subfamily are indicated by the red arrows.
Supplementary information
Supplementary Information
This file contains Supplementary Figs. 1–8, Supplementary Tables 1–11 and legends for Supplementary Videos 1–6.
Video 1
MD simulation of hM3R–iperoxo. The iperoxo and surrounding residues are coloured cyan and violet, respectively.
Video 2
MD simulation of hM3Dq–iperoxo. The iperoxo and surrounding residues are coloured cyan and violet, respectively.
Video 3
ECL2 dynamics in the MD simulation of hM3R–Apo. The top view of the hM3R simulations and Cα of T215 and F222 (used in the distance measurement) are shown in spheres.
Video 4
ECL2 dynamics in the MD simulation of hM3Dq–iperoxo. The top view of the hM3Dq–iperoxo simulations and Cα of T215 and F222 (used in the distance measurement) are shown in spheres.
Video 5
MD simulation of hM3R(A239G)–iperoxo. The iperoxo and surrounding residues are coloured cyan and violet, respectively.
Video 6
MD simulation of hM3Dq(G239A)–DCZ. The DCZ and surrounding residues are coloured cyan and violet, respectively.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Zhang, S., Gumpper, R.H., Huang, XP. et al. Molecular basis for selective activation of DREADD-based chemogenetics. Nature 612, 354–362 (2022). https://doi.org/10.1038/s41586-022-05489-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-022-05489-0
This article is cited by
-
Psychedelics: preclinical insights provide directions for future research
Neuropsychopharmacology (2024)
-
Programmable synthetic receptors: the next-generation of cell and gene therapies
Signal Transduction and Targeted Therapy (2024)
-
Molecular mechanism of muscarinic acetylcholine receptor M3 interaction with Gq
Communications Biology (2024)
-
Structure, function and drug discovery of GPCR signaling
Molecular Biomedicine (2023)
-
Xanomeline displays concomitant orthosteric and allosteric binding modes at the M4 mAChR
Nature Communications (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
