
Abstract
Wnt signaling plays an important role in regulating cell proliferation and differentiation. De-regulation of these signaling pathways has been implicated in many human diseases, ranging from cancers to skeletal disorders. Wnt proteins are a large family of secreted factors that bind to the Frizzled receptors and LRP5/6 co-receptors and initiate complex signaling cascades. Over the past two decades, our understanding of Wnt signaling has been significantly improved due to the identification of many key regulators and mediators of these pathways. Given that Wnt signaling is tightly regulated at multiple cellular levels, these pathways themselves offer ample nodal points for targeted therapeutics. Here, we focus on our current understanding of these pathways, the associations of Wnt signaling with human disorders, and the opportunities to target key components of Wnt signaling for rational drug discovery.
Similar content being viewed by others

Main
Wnt signaling molecules regulate many aspects of animal development and tissue homeostasis. The term ‘Wnt’ was coined from a combination of the Drosophila segment polarity gene Wingless and the mouse proto-oncogene Int-1.1 Many components of Wnt signaling are evolutionarily conserved and there are 19 human Wnt genes.2 Wnt proteins are a large family of secreted cysteine-rich glycoproteins that act as short-range ligands to locally activate receptor-mediated signaling cascades.3 Expression of Wnt genes is developmentally regulated in a coordinated temporal and spatial manner.2
Wnts play important roles in cell fate specification, tissue patterning, and control of asymmetric cell division, whereas Wnt signaling in mammals is more complex and less well-defined.3 Wnts activate at least three signaling pathways. The best understood one is the canonical Wnt pathway that activates transcriptional activity of β-catenin/Tcf.4 By contrast, non-canonical Wnts seem to act through β-catenin-independent pathways that regulate cell movement and organization.5 Abnormal Wnt signaling has been associated with many human diseases, ranging from cancer to degenerative diseases. Thus, studies of Wnt signaling have opened up new avenues in biomedical research, including molecular embryology, stem cell biology, tumorigenesis, regenerative medicine, and rational drug discovery.3, 6, 7
Canonical Wnt signaling pathway and its biological outcomes
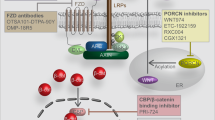
In unstimulated cells, free β-catenin is rapidly degraded through proteosome machinery promoted by APC/Axin/GSK3β complex (Figure 1a). Canonical Wnts bind to Frizzled (Fz) receptor and LRP5/6 co-receptors. Soluble FRPs act as antagonists by squelching Wnts.3 Similarly, Dkk proteins sterically hinder Wnt interaction with LRP5/6. Upon binding of Wnts, Axin translocates to cell membrane and interacts with LRP5/6, and Wnt binding to Fz results in hyperphosphorylation of Disheveled (Dvl) by Casein kinases (CK) Iɛ and II, preventing GSK3β from phosphorylating β-catenin.3 Unphosphorylated β-catenin is stabilized by escaping the recognition of by β-TrCP and translocates to nucleus where it engages LEF/Tcf to activate expression of downstream genes (Figure 1b).3

Schematic representation of the canonical Wnt/β-catenin signaling pathway. (a) In the absence of Wnt signal, β-catenin is recruited into the APC/Axin/GSK3β complex, and phosphorylated by GSK3β at the N-terminal ‘destruction box’. The phosphorylated β-catenin binds to β-Trcp of the proteosome machinery and is targeted for degradation. As the result, no free β-catenin enters nucleus to form transcriptional complex with LEF/Tcf and to regulate downstream gene expression. (b) Wnt binds to its Fz receptor and LRP5/6 co-receptor and activates Dvl, leading to the inhibition of APC/Axin/GSK3β-mediated β-catenin degradation. Stabilized β-catenin forms a transcriptional complex with LEF/Tcf and activates downstream targets such as c-Myc. Negative regulators, such as Dkks, WIF and FRPs, inhibit the interactions between Wnt ligands and their receptors. There is crosstalk between Wnts and other signaling pathways, such as growth factors that activate receptor tyrosine kinases. See text for details.
The hallmark of canonical Wnt signaling activation is cytoplasmic accumulation of β-catenin protein. Without Wnts, Tcf/LEF acts as transcriptional repressor by forming a complex with Groucho (Figure 1a). Interaction of β-catenin with Tcf converts it into an activator and induces β-catenin/Tcf target genes, such as c-Myc, cyclin D1, and PPARδ.3, 8, 9, 10 Wnt signaling produces distinct transcriptional outputs, depending on developmental stages and cell types, whereas it remains unclear whether ‘universal target genes’ of Wnt/β-catenin actually exist. Overall outcomes of canonical Wnt signaling include promoting cell proliferation and tissue expansion, and controlling fate determination or terminal differentiation of postmitotic cells.
Non-canonical Wnt signaling pathways and crosstalk with other signaling pathways
Two non-canonical pathways transduce Wnt signals through either JNK/planar cell polarity (PCP) pathway or the calcium-releasing pathway.7 Although possible mechanisms remain to be defined, Dvl may function as a major switchboard between canonical and non-canonical pathways. It is noteworthy that the non-canonical Wnt pathways are poorly understood in mammalian cells.
Wnt/β-catenin signaling may be activated through Wnt-independent mechanisms, including G-protein, ILK, HGF, Gas6, IGF-1, IGF-II, FGF-1, and VEGF. Cross talk occurs between Wnt signaling and other pathways, including MAPK, Ca++/calmodulin kinase II, TGFβ, PKC and retinoid signaling.6
Wnt signaling in stem cell fate determination
Wnt signals not only regulate many developmental processes, but also play an important role in controlling adult tissue self-renewal.3 De-regulation of Wnt pathways tips the homeostatic balance in these tissues.
Intestinal Epithelial Stem Cells
Wnt/β-catenin signaling is a dominant force in controlling cell fate along the crypt-villus axis.3 Mice lacking Tcf4 exhibit the complete absence of the crypt progenitor compartment.11 Conversely, the inhibition of Wnt signaling by the transgenic expression of Dkk1 also results in the complete loss of crypts.3 These findings indicate that Wnt signaling is critical for the proliferation of crypt progenitor cells, and the terminal differentiation of Paneth cells.
Hematopoietic Stem Cells
Wnt signaling may regulate self-renewal of hematopoietic stem cells (HSCs) in an autocrine or paracrine fashion, as Wnts are produced by these progenitors as well as by bone marrow microenvironment.4 Soluble Wnt proteins or overexpression of β-catenin promote proliferation and inhibit differentiation, leading to sustained self-renewal of mouse and human HSCs in vitro.12 Interestingly, mice with conditionally deleted β-catenin revealed no significant hematolymphoid defects.13 Nevertheless, understanding Wnt signaling in HSC differentiation would be facilitated by elucidating the function of specific Wnts produced at the HSC niche.14 Wnt signaling is also implicated in fate determination within the T-lymphoid lineage, as mice lacking Tcf1 or Tcf1/Lef1 exhibited a severe reduction of the earliest thymocyte progenitor compartment.15
Hair Follicle Stem Cells
Wnt signaling is required for the establishment of hair follicle.16 Multipotent bugle stem cells give rise to all hair lineages, the sebocytes, and progenitor cells of the interfollicular epidermis. Mice with mutant Lef1 have few fair follicles.17 Conversely, transgenic overexpression of Lef1 or a constitutively active form of β-catenin results in de novo hair follicle formation in epidermis.16 Multiple hair keratin genes are β-catenin/Tcf targets,18 suggesting that Wnt signaling not only controls hair follicle precursor expansion, but also promotes terminal differentiation of the hair lineage.
Mesenchymal Stem Cells
Mesenchymal stem cells (MSCs) are pluripotent adherent bone marrow stromal cells, which give rise to osteoblastic, chondrogenic, adipogenic and myogenic lineages. Wnt signaling plays an important role in controlling MSC differentiation. Canonical Wnts have been shown to induce osteogenic differentiation.19, 20 Wnt signaling may play dual roles in chondrogenesis.21 Activation of Wnt signaling by Wnt10B inhibits differentiation of preadipocytes and blocks adipose tissue development.22 Wnt signaling also plays a role in regulating myogenesis in mammals.23 When Wnt signaling in preadipocytes is prevented, MSCs differentiate into adipocytes.24 Disruption of Wnt signaling also causes transdifferentiation of myoblasts into adipocytes.24
Aberrant Wnt signaling and human diseases
Given the important and diverse biological functions of Wnt signaling, it is conceivable that defects or deregulations of Wnt signaling pathways may lead to the development of human diseases.3, 6, 7 Table 1 provides a partial list of human diseases that are caused by or associated with currently known Wnt signaling miscues.
Activated Wnt Signaling and Cancers
APC functions as a gatekeeper in colorectal tumorigenesis and both alleles of APC are inactivated in nearly 80% of sporadic colorectal cancers.25 Although the involvement of β-catenin in tumorigenesis was first established in colorectal cancers, the importance of β-catenin in tumorigenesis was highlighted by the discovery of oncogenic β-catenin mutations in colon cancers containing wild-type APC, and many other types of human cancer.26, 27 Mutations of the Axin gene and/or its loss of expression have been found in colorectal cancers, esophageal squamous cell carcinomas, hepatocellular carcinomas, and medulloblastomas.3 Aberrant Wnt signaling may play a role in lymphoid neoplasias, such as pre-B-cell leukemia and myeloid leukemia.28 Interestingly, the overexpression of Dkk1 by myeloma cells is associated with the presence of lytic bone lesions in patients with multiple myeloma.29 Thus, deregulation of Wnt/β-catenin signaling may be involved in the development of many human malignancies. By contrast, non-canonical Wnt5A may function as a tumor suppressor, as Wnt5A is deleted or has reduced expression in hemetopoietic tumors.28 Mice that are hemizygous for Wnt5A develop myeloid leukemia and B-cell lymphomas.28
Wnt Signaling and Developmental Disorders
A nonsense mutation in the Wnt3 gene causes the autosomal recessive disorder Tetra-Amelia (loss of four limbs) in a large consanguineous Turkish family,30 consistent with the important role of Wnts in vertebrate limb development.3 Duplication of Wnt4 gene is linked to an intersex phenotype of an XY patient with ambiguous genitalia.31 Wnt4 and Wnt11 may be involved in kidney development, as revealed by analysis of knockout mice.32 Elevated expression of Wnt4 has been reported in polycystic kidneys in mice, and transgenic overexpression of active forms of β-catenin develops polycystic kidney disease.33 Loss of function mutations of Fz4 and LRP5 are linked to familial exudative vitreoretinopathy (FEVR).34, 35, 36
Wnt Signaling and Skeletal Disorders
Loss-of-function mutations of LRP5 are linked to the low bone mass observed in osteoporosis-pseudoglioma syndrome (OPPG).37 Conversely, gain-of-function LRP5 mutants result in a high bone density and elevated numbers of active osteoblasts in transgenic mice.38, 39 Mice lacking Axin2 exhibit craniosynostosis, resulting from increased proliferation of osteoblast progenitors in the skull sutures.40 Wnt10B mutant animals display a postnatal decrease in bone mass, while overexpression of Wnt4, or Wnt9A/Wnt14 enhances maturation of chondrocytes.20 Naturally occurring missense variants of WNT10B may be linked to human obesity.41 Axin2 mutations have recently been linked to familial tooth agenesis.42 A polymorphic SNP found in FRP3, resulting in a reduced ability to antagonize Wnt signaling, is associated with higher incidence of osteoarthritis in females.43
Wnt Signaling and Neuronal Diseases
Increased expression of Wnt1 may lead to synaptic rearrangement and plasticity in the brains of patients with schizophrenia.44 Several SNPs in Fz3 are associated with susceptibility to schizophrenia,45 and GSK3 activity is reportedly altered in schizophrenia.46 Attenuated β-catenin signaling may be implicated in the development of Alzheimer's disease (AD), the most common neurodegenerative disease characterized with extracellular deposits of the amyloid β-peptide, intracellular deposits of hyperphosphorylated tau, and selective death of the cholinergic neurons.7 β-Catenin and GSK3β are components of presenilin complexes, and loss of Wnt/β-catenin function may underlie the unset and development of AD.47, 48
Wnt Signaling and Skin Disorders
Wnt signaling in the skin regulates stem cell self-renewal and differentiation, hence influencing hair formation.16 Fz6 has been shown to control mammalian hair pattern.49 Wnt/β-catenin promotes the recruitment of fibroblast to wounds of the skin, whereas uncontrolled Wnt signaling may lead to aggressive fibromatosis, resembling a hyperactive wound healing process.50 Similarly, elevated β-catenin signaling is detected in Dupuytren disease, which is a superficial fibromatosis with a profusion of collagen-rich areas of the hand.51
Wnt Signaling and Cardiovascular Diseases
Both canonical Wnt/β-catenin and non-canonical Wnt/calcium pathways are implicated in cardiogenesis and heart diseases.52, 53 For example, mice lacking Dvl1 are more prone to infarct rupture after induction of myocardial infarction.53 Wnt signaling may also be involved in neovascularization in the infarct area. Overexpression of sFRP1 reduces infarct size and improves cardiac function in mice.54 Fz2 is upregulated in cardiac hypertrophy model of rats.53
Therapeutic implications by targeting Wnt signaling pathways
Current strategies targeting Wnt signaling have mostly focused on anticancer therapies.6 However, it is conceivable that, with the improvement of our understanding of these pathways, aberrant Wnt signaling may be targeted as potential treatments of other diseases, such as neurodegenerative diseases, degenerative bone diseases, and cardiovascular diseases.6, 7
Targeting Wnt Signaling at Extracellular Level
Wnt ligands themselves can be targeted by antisense molecules, RNAi-based strategy, and neutralizing antibodies.6 It is conceivable that the Wnt signaling antagonists, such as FRPs and Dkks, could be used as natural inhibitors of Wnt signaling in cancer therapy. For example, expression of Dkk-3 was significantly downregulated in primary non-small cell lung carcinomas, and expression of exogenous Dkk-3 gene in non-small cell lung carcinoma cells inhibited cell growth.55 Expression of Dkk-3 and dominant-negative LRP5 mutant in Saos-2 cells significantly reduces invasion capacity and cell motility.56
Targeting Wnt Signaling at Cytoplasmic Level
Wnt/β-catenin signaling is subjected to tight regulations exerted by a network of intracellular protein–protein interactions. Consistent with the central role the APC tumor suppressor in the development of human colorectal cancers,25 the β-catenin-binding domain of APC is sufficient for tumor suppression.57 Introduction of wild-type Axin1-induced apoptosis in hepatocellular and colorectal cancers.58 ICAT was also shown to inhibit the anchorage-independent growth of colorectal tumor cells and tumor growth of colorectal tumor.59
Targeting β-Catenin Expression and Protein Degradation
Direct targeting β-catenin has attracted a broad range of attentions since the commonality of many human cancers with aberrant Wnt signaling results in increased β-catenin levels. Several approaches have been pursued, including anti-sense, RNA interference, and protein knockdown strategies.6 Antisense oligonucleotides directed against β-catenin can decrease β-catenin expression in the tumors and tumor growth, and were able to bring about complete tumor regression, suggesting that these antisense oligonucleotides are effective in reducing tumor growth, and may be of therapeutic value.6 The RNAi approach has also been used to target β-catenin in both cell cultures and in a xenograft tumor model.60 Several recent studies have explored the feasibility of using this degradation process to accelerate the turnover rate of oncogenic β-catenin activity in human tumor cells. It was shown that the stable β-catenin mutant was recruited to the cellular SCF ubiquitination machinery for ubiquitination and degradation.61 Introduction of a chimeric F box protein with multiple copies of the APC 15-amino-acid repeat unit to colon cancer cells induced targeted ubiquitination and proteolytic degradation of free β-catenin while preserving its cellular adhesion counterpart.62 Using a similar strategy, expression of chimeric F-box fusion proteins with the β-catenin-binding domains of Tcf-4 and E-cadherin successfully promotes degradation of β-catenin independent of GSK-3β-mediated phosphorylation.63
Targeting Wnt Signaling at Nuclear Level
Recent structural elucidation of the β-catenin/Tcf complex further highlights the possibility of developing cancer drugs via rational design and high throughput screening.64 Several strategies have been developed by targeting the β-catenin/Tcf transcriptional activity.6 A binding assay for high throughput screening was developed to identify compounds that inhibit association between Tcf4 and β-catenin.65 Using this high-throughput assay, Lepourcelet et al screened nearly 7000 natural compounds and identified several compounds that were shown to disrupt Tcf/β-catenin complexes and to potently antagonize β-catenin-dependent activities.65 Several studies have also explored the use of conditionally replication-competent oncolytic adenoviral vectors that only target tumor cells with aberrantly high β-catenin/Tcf activity.6 It is also conceivable that some of the important downstream mediators of β-catenin/Tcf signaling, such as c-Myc, cyclin D1, PPARδ, COX-2, CD44 and MMP7, can be targeted.6
Targeting Signaling Synergy between Wnt and Other Pathways
Wnt/β-catenin signaling may be regulated through Wnt-independent mechanisms.6 For example, growth factors that activate receptor tyrosine kinases may regulate β-catenin activity.6, 66 It has been shown that tyrosine kinase inhibitors, such as STI-571/Gleevec, downregulate β-catenin signaling.66 An in vivo study on combinatorial chemoprevention of intestinal neoplasia was reported by targeting both Wnt and EGF pathways.67 Retinoic acids have been shown to inhibit β-catenin/Tcf activity.6 These findings are encouraging and open the door for drug therapies by targeting both Wnt and other signaling pathways.
Stem Cell-Based Therapy and Regenerative Medicine
Wnt signaling plays an important role in development and in regulating stem cell proliferation and self-renewal.4 These functional features render Wnts and their signaling mediators appealing therapeutic agents for regenerative medicine. For example, some Wnts may be used as therapeutic agents for osteoporosis by promoting osteoblast differentiation and bone formation, while some Wnts may be used as a treatment of neurodegenerative diseases. Wnts may also be used through a stem cell-based therapy approach to facilitate organ and/or tissue regeneration, or to enhance injury healing. However, these applications would require better understanding of the biological functions of Wnts in stem cells in order to avoid possible detrimental adverse effects associated uncontrolled stem cell proliferation.
Unanswered questions and future directions
A quarter-century-long study of Wnt signaling has significantly advanced our understanding about many aspects of Wnt pathways.3, 6, 7 However, the tasks to thoroughly understand these pathways remain daunting. Technical challenges to obtain highly purified and biologically active recombinant Wnt proteins not only have slowed down the structural and biological studies, but also hamper their possible use as therapeutic agents. As Wnt signaling in mammals is more complex than that in model organisms, many questions need to be addressed. For most of the ∼20 Wnts in mammals, we do not know what their biological functions are and how they initiate their signaling cascades. How do different Wnt proteins utilize Fz receptors and LRP5/6 co-receptors? Recent studies reported that Wnts may also relay their signals through atypical receptors.3 Do these atypical receptors play a universal role in Wnt signaling or a limited role in certain specific tissue/cell types? What are the role(s), if any, of Wnts in neuronal development? How does Wnt signaling crosstalk with other important signaling pathways, especially receptor tyrosine kinases? What are the molecular determinants/switches for canonical vs non-canonical Wnt signaling pathways? What causes the accumulation of cytoplasmic/nuclear β-catenin protein in many human tumors without any detectable genetic defects in the canonical Wnt pathway? Addressing questions would not only allow us to better understand possible pathogenic roles of aberrant Wnt signaling in human diseases, but also provide opportunities to target the pathways by rational drug discovery.
References
Nusse R, Varmus HE . Many tumors induced by the mouse mammary tumor virus contain a provirus integrated in the same region of the host genome. Cell 1982;31:99–109.
Wodarz A, Nusse R . Mechanisms of Wnt signaling in development. Annu Rev Cell Dev Biol 1998;14:59–88.
Clevers H . Wnt/beta-catenin signaling in development and disease. Cell 2006;127:469–480.
Reya T, Clevers H . Wnt signalling in stem cells and cancer. Nature 2005;434:843–850.
Kohn AD, Moon RT . Wnt and calcium signaling: beta-catenin-independent pathways. Cell Calcium 2005;38:439–446.
Luu HH, Zhang R, Haydon RC, et al. Wnt/beta-catenin signaling pathway as a novel cancer drug target. Curr Cancer Drug Targets 2004;4:653–671.
Moon RT, Kohn AD, De Ferrari GV, et al. WNT and beta-catenin signalling: diseases and therapies. Nat Rev Genet 2004;5:691–701.
He TC, Sparks AB, Rago C, et al. Identification of c-MYC as a target of the APC pathway [see comments]. Science 1998;281:1509–1512.
Tetsu O, McCormick F . Beta-catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature 1999;398:422–426.
He TC, Chan TA, Vogelstein B, et al. PPARdelta is an APC-regulated target of nonsteroidal anti-inflammatory drugs. Cell 1999;99:335–345.
Korinek V, Barker N, Moerer P, et al. Depletion of epithelial stem-cell compartments in the small intestine of mice lacking Tcf-4. Nat Genet 1998;19:379–383.
Van Den Berg DJ, Sharma AK, Bruno E, et al. Role of members of the Wnt gene family in human hematopoiesis. Blood 1998;92:3189–3202.
Cobas M, Wilson A, Ernst B, et al. Beta-catenin is dispensable for hematopoiesis and lymphopoiesis. J Exp Med 2004;199:221–229.
Zhang J, Niu C, Ye L, et al. Identification of the haematopoietic stem cell niche and control of the niche size. Nature 2003;425:836–841.
Okamura RM, Sigvardsson M, Galceran J, et al. Redundant regulation of T cell differentiation and TCRalpha gene expression by the transcription factors LEF-1 and TCF-1. Immunity 1998;8:11–20.
Fuchs E, Tumbar T, Guasch G . Socializing with the neighbors: stem cells and their niche. Cell 2004;116:769–778.
van Genderen C, Okamura RM, Farinas I, et al. Development of several organs that require inductive epithelial-mesenchymal interactions is impaired in LEF-1-deficient mice. Genes Dev 1994;8:2691–2703.
Zhou P, Byrne C, Jacobs J, et al. Lymphoid enhancer factor 1 directs hair follicle patterning and epithelial cell fate. Genes Dev 1995;9:700–713.
Si W, Kang Q, Luu HH, et al. CCN1/Cyr61 is regulated by the canonical Wnt signal and plays an important role in Wnt3A-induced osteoblast differentiation of mesenchymal stem cells. Mol Cell Biol 2006;26:2955–2964.
Hartmann C . A Wnt canon orchestrating osteoblastogenesis. Trends Cell Biol 2006;16:151–158.
Hartmann C, Tabin CJ . Dual roles of Wnt signaling during chondrogenesis in the chicken limb. Development 2000;127:3141–3159.
Bennett CN, Longo KA, Wright WS, et al. Regulation of osteoblastogenesis and bone mass by Wnt10b. Proc Natl Acad Sci USA 2005;102:3324–3329.
Cossu G, Borello U . Wnt signaling and the activation of myogenesis in mammals. EMBO J 1999;18:6867–6872.
Ross SE, Hemati N, Longo KA, et al. Inhibition of adipogenesis by Wnt signaling. Science 2000;289:950–953.
Kinzler KW, Vogelstein B . Lessons from hereditary colorectal cancer. Cell 1996;87:159–170.
Morin PJ . beta-catenin signaling and cancer. Bioessays 1999;21:1021–1030.
Polakis P . The oncogenic activation of beta-catenin. Curr Opin Genet Dev 1999;9:15–21.
Liang H, Chen Q, Coles AH, et al. Wnt5a inhibits B cell proliferation and functions as a tumor suppressor in hematopoietic tissue. Cancer Cell 2003;4:349–360.
Tian E, Zhan F, Walker R, et al. The role of the Wnt-signaling antagonist DKK1 in the development of osteolytic lesions in multiple myeloma. N Engl J Med 2003;349:2483–2494.
Niemann S, Zhao C, Pascu F, et al. Homozygous WNT3 mutation causes tetra-amelia in a large consanguineous family. Am J Hum Genet 2004;74:558–563.
Jordan BK, Shen JH, Olaso R, et al. Wnt4 overexpression disrupts normal testicular vasculature and inhibits testosterone synthesis by repressing steroidogenic factor 1/beta-catenin synergy. Proc Natl Acad Sci USA 2003;100:10866–10871.
Perantoni AO . Renal development: perspectives on a Wnt-dependent process. Semin Cell Dev Biol 2003;14:201–208.
Rodova M, Islam MR, Maser RL, et al. The polycystic kidney disease-1 promoter is a target of the beta-catenin/T-cell factor pathway. J Biol Chem 2002;277:29577–29583.
Robitaille J, MacDonald ML, Kaykas A, et al. Mutant frizzled-4 disrupts retinal angiogenesis in familial exudative vitreoretinopathy. Nat Genet 2002;32:326–330.
Toomes C, Bottomley HM, Jackson RM, et al. Mutations in LRP5 or FZD4 underlie the common familial exudative vitreoretinopathy locus on chromosome 11q. Am J Hum Genet 2004;74:721–730.
Kondo H, Hayashi H, Oshima K, et al. Frizzled 4 gene (FZD4) mutations in patients with familial exudative vitreoretinopathy with variable expressivity. Br J Ophthalmol 2003;87:1291–1295.
Gong Y, Slee RB, Fukai N, et al. LDL receptor-related protein 5 (LRP5) affects bone accrual and eye development. Cell 2001;107:513–523.
Little RD, Carulli JP, Del Mastro RG, et al. A mutation in the LDL receptor-related protein 5 gene results in the autosomal dominant high-bone-mass trait. Am J Hum Genet 2002;70:11–19.
Boyden LM, Mao J, Belsky J, et al. High bone density due to a mutation in LDL-receptor-related protein 5. N Engl J Med 2002;346:1513–1521.
Yu HM, Jerchow B, Sheu TJ, et al. The role of Axin2 in calvarial morphogenesis and craniosynostosis. Development 2005;132:1995–2005.
Christodoulides C, Scarda A, Granzotto M, et al. WNT10B mutations in human obesity. Diabetologia 2006;49:678–684.
Lammi L, Arte S, Somer M, et al. Mutations in AXIN2 cause familial tooth agenesis and predispose to colorectal cancer. Am J Hum Genet 2004;74:1043–1050.
Loughlin J, Dowling B, Chapman K, et al. Functional variants within the secreted frizzled-related protein 3 gene are associated with hip osteoarthritis in females. Proc Natl Acad Sci USA 2004;101:9757–9762.
Miyaoka T, Seno H, Ishino H . Increased expression of Wnt-1 in schizophrenic brains. Schizophr Res 1999;38:1–6.
Katsu T, Ujike H, Nakano T, et al. The human frizzled-3 (FZD3) gene on chromosome 8p21, a receptor gene for Wnt ligands, is associated with the susceptibility to schizophrenia. Neurosci Lett 2003;353:53–56.
Kozlovsky N, Belmaker RH, Agam G . GSK-3 and the neurodevelopmental hypothesis of schizophrenia. Eur Neuropsychopharmacol 2002;12:13–25.
Mudher A, Lovestone S . Alzheimer's disease-do tauists and baptists finally shake hands? Trends Neurosci 2002;25:22–26.
Caricasole A, Copani A, Caruso A, et al. The Wnt pathway, cell-cycle activation and beta-amyloid: novel therapeutic strategies in Alzheimer's disease? Trends Pharmacol Sci 2003;24:233–238.
Guo N, Hawkins C, Nathans J . Frizzled6 controls hair patterning in mice. Proc Natl Acad Sci USA 2004;101:9277–9281.
Cheon SS, Cheah AY, Turley S, et al. Beta-catenin stabilization dysregulates mesenchymal cell proliferation, motility, and invasiveness and causes aggressive fibromatosis and hyperplastic cutaneous wounds. Proc Natl Acad Sci USA 2002;99:6973–6978.
Varallo VM, Gan BS, Seney S, et al. Beta-catenin expression in Dupuytren's disease: potential role for cell-matrix interactions in modulating beta-catenin levels in vivo and in vitro. Oncogene 2003;22:3680–3684.
Olson EN, Schneider MD . Sizing up the heart: development redux in disease. Genes Dev 2003;17:1937–1956.
van Gijn ME, Daemen MJ, Smits JF, et al. The wnt-frizzled cascade in cardiovascular disease. Cardiovasc Res 2002;55:16–24.
Barandon L, Couffinhal T, Ezan J, et al. Reduction of infarct size and prevention of cardiac rupture in transgenic mice overexpressing FrzA. Circulation 2003;108:2282–2289.
Tsuji T, Nozaki I, Miyazaki M, et al. Antiproliferative activity of REIC/Dkk-3 and its significant down-regulation in non-small-cell lung carcinomas. Biochem Biophys Res Commun 2001;289:257–263.
Hoang BH, Kubo T, Healey JH, et al. Dickkopf 3 inhibits invasion and motility of Saos-2 osteosarcoma cells by modulating the Wnt-beta-catenin pathway. Cancer Res 2004;64:2734–2739.
Shih IM, Yu J, He TC, et al. The beta-catenin binding domain of adenomatous polyposis coli is sufficient for tumor suppression [in process Citation]. Cancer Res 2000;60:1671–1676.
Satoh S, Daigo Y, Furukawa Y, et al. AXIN1 mutations in hepatocellular carcinomas, and growth suppression in cancer cells by virus-mediated transfer of AXIN1. Nat Genet 2000;24:245–250.
Sekiya T, Nakamura T, Kazuki Y, et al. Overexpression of Icat induces G(2) arrest and cell death in tumor cell mutants for adenomatous polyposis coli, beta-catenin, or Axin. Cancer Res 2002;62:3322–3326.
Verma UN, Surabhi RM, Schmaltieg A, et al. Small interfering RNAs directed against beta-catenin inhibit the in vitro and in vivo growth of colon cancer cells. Clin Cancer Res 2003;9:1291–1300.
Cong F, Zhang J, Pao W, et al. A protein knockdown strategy to study the function of beta-catenin in tumorigenesis. BMC Mol Biol 2003;4:10.
Su Y, Ishikawa S, Kojima M, et al. Eradication of pathogenic beta-catenin by Skp1/Cullin/F box ubiquitination machinery. Proc Natl Acad Sci USA 2003;100:12729–12734.
Liu J, Stevens J, Matsunami N, et al. Targeted degradation of beta-catenin by chimeric F-box fusion proteins. Biochem Biophys Res Commun 2004;313:1023–1029.
Daniels DL, Eklof Spink K, Weis WI . beta-catenin: molecular plasticity and drug design. Trends Biochem Sci 2001;26:672–678.
Lepourcelet M, Chen YN, France DS, et al. Small-molecule antagonists of the oncogenic Tcf/beta-catenin protein complex. Cancer Cell 2004;5:91–102.
Zhou L, An N, Haydon RC, et al. Tyrosine kinase inhibitor STI-571/Gleevec down-regulates the beta-catenin signaling activity. Cancer Lett 2003;193:161–170.
Torrance CJ, Jackson PE, Montgomery E, et al. Combinatorial chemoprevention of intestinal neoplasia. Nat Med 2000;6:1024–1028.
Acknowledgements
We apologize to those authors whose original studies were not cited due to space constraints. The reported work was supported, in part, by research grants from The Brinson Foundation (TCH), American Cancer Society (TCH, HHL), and the National Institutes of Health (TCH, RCH). TCH was a recipient of the Outstanding Overseas Young Investigator Collaboration Award from the Natural Science Foundation of China (NSFC #30228026) and a recipient of the Bayu Scholar of Chongqing Municipality, Chongqing, China.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interest statement
The authors declare no conflict of interest.
Rights and permissions
About this article
Cite this article
Luo, J., Chen, J., Deng, ZL. et al. Wnt signaling and human diseases: what are the therapeutic implications?. Lab Invest 87, 97–103 (2007). https://doi.org/10.1038/labinvest.3700509
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/labinvest.3700509
Keywords
This article is cited by
-
Head and neck squamous cell carcinoma: a potential therapeutic target for the Wnt signaling pathway
European Journal of Plastic Surgery (2022)
-
TGF-β and WNT signaling pathways in cardiac fibrosis: non-coding RNAs come into focus
Cell Communication and Signaling (2020)
-
Ubiquitylation and degradation of adenomatous polyposis coli by MKRN1 enhances Wnt/β-catenin signaling
Oncogene (2018)
-
Phytocannabinoids modulate emotional memory processing through interactions with the ventral hippocampus and mesolimbic dopamine system: implications for neuropsychiatric pathology
Psychopharmacology (2018)
-
The hypothesis that Helicobacter pylori predisposes to Alzheimer’s disease is biologically plausible
Scientific Reports (2017)
