
Abstract
Objective: To reassess the validity of the Schofield equations for predicting basal metabolic rate (BMR) in the obese.
Data: The data collection compiled from many studies by Schofield in 1985.
Methods: Nonparametric curve fitting and comparison of groups.
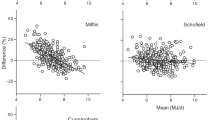
Results: BMR increases more slowly at heavier weights in men (above about 75 kg) and women (above about 65 kg), and to ignore this is to predict overestimates of BMR. Data obtained from studies of different populations show substantial differences, which are influential when data are combined.
Conclusions: The Schofield equations are unsuitable for obese populations. Given that current Western populations exhibit prevalences of obesity many times greater than those in the Schofield database, this indicates the need for further study of suitable predictors for these individuals.
Sponsorship: Ministry of Agriculture, Fisheries and Food, UK.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

References
Chitre, RG, Moseby, AM, Thien, K & Hla Pe, U (1959). Studies on basal metabolism of Burmese people. J. Postgrad. Med., 5, 129–137.
DOH (1999). Health Survey for England, 1999, London: UK Department of Health available at www.doh.gov.uk/public/summary1.htm
FAO/WHO/UNU (1985). Energy and protein requirements, Technical Report Series no. 724 Geneva: WHO
Felloni, G (1936). Contributo alla conoscenza de metabolismo di base degli Italiani. Nota I: il metabolismo di base di sogetti dai 19 ai 25 anni. Quad. Nutr., 3, 419–442.
Forbes, GB (1982). Human Body Composition: Growth, Ageing, Nutrition and Activity, New York: Springer
Forbes, GB (1987). Lean body mass–fat interrelationship in humans. Nutr. Rev., 45, 225–231.
Hastie, TJ & Tibshirani, RJ (1990). Generalized Additive Models, London: Chapman and Hall
Hayter, JE & Henry, CJK (1994). A reexamination of basal metabolic-rate predictive equations—the importance of geographic origin of subjects in sample selection. Eur. J. Clin. Nutr., 48, 702–707.
James, WPT (1992). From S.D.A. to D.I.T. to T.E.F. InEnergy Metabolism: Tissue Determinants and Cellular Corollaries, ed. JM Kinney
Jequier, E (1984). Energy expenditure in obesity. Clin. Endocrinol. Metab., 13, (3) 563–580.
Lenti, C (1937). Contributo alla conoscenza de metabolismo di base degli Italiani. Nota IV: il metabolismo di base di sogetti dai 20 ai 25 anni. Quad. Nutr., 4, 192–225.
McNeill, G, Rivers, JPW, Payne, PR, de Britto, JS & Abel, R (1987). Basal metabolic rate of Indian man. No evidence of metabolic adaptation to a low plane of nutrition. Hum. Nutr. Clin. Nutr., 41c, 473–483.
Miller, CD & Benedict, FG (1937). I: Basal Metabolism of Normal Young Men and Women of Various Races in Hawaii, University of Hawaii Research Publications, no. 15, p 1 Honolulu: University of Hawaii
Miller, AT & Blyth, CS (1953). Lean body mass as a metabolic reference standard. J. Appl. Physiol., 5, 311–316.
Occhuito, A & Pepe, M (1940). Contributo alla conoscenza de metabolismo di base degli Italiani. Nota X: il metabolismo di base di sogetti (maschi) di oltre 20 anni. Quad. Nutr., 7, 142–154.
Pepe, M (1938). Contributo alla conoscenza de metabolismo di base degli Italiani. Nota VII: il metabolismo di base di sogetti dai 18 ai 24 anni. Quad. Nutr., 5, 206–214.
Prentice, AM, Black, AE, Murgatroyd, PR, Goldberg, GR & Coward, WA (1989). Metabolism or appetite: questions of energy balance with particular reference to obesity. J. Hum. Nutr. Diet, 2, 95–104.
Pullicino, E, Copperstone, C, Luzi, L, McNeill, G & Elia, M (1996). Relationship between anthropometric indices of body fat distribution and basal energy metabolism in healthy Maltese women. Acta Diabetol., 33, 198–204.
Rucker, RB (1978). Elevated metabolic rates in obesity. Lancet, 8080, 106–107.
Schofield, W (1985a). Predicting basal metabolic rate, new standards and review of previous work. Clin. Nutr., 39C, (Suppl 1) 5–41.
Schofield, C (1985b). An annotated bibliography of source material for basal metabolic rate data. Clin. Nutr., 39C, (Suppl 1) 42–91.
Webb, P (1981). Energy expenditure and fat free mass in men and women. J. Clin. Nutr., 34, 1816–1826.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Horgan, G., Stubbs, J. Predicting basal metabolic rate in the obese is difficult. Eur J Clin Nutr 57, 335–340 (2003). https://doi.org/10.1038/sj.ejcn.1601542
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.ejcn.1601542
Keywords
This article is cited by
-
Misreporting of dietary energy intake obtained by 24 h recalls in older adults: a comparison of five previous methods using doubly labeled water
European Journal of Clinical Nutrition (2022)
-
Propofol Sedation for Intragastric Balloon Removal: Looking for the Optimal Body Weight Descriptor
Obesity Surgery (2019)
-
Chronic Repression of mTOR Complex 2 Induces Changes in the Gut Microbiota of Diet-induced Obese Mice
Scientific Reports (2016)
-
Plausible self-reported dietary intakes in a residential facility are not necessarily reliable
European Journal of Clinical Nutrition (2016)
-
An approach to quantifying abnormalities in energy expenditure and lean mass in metabolic disease
European Journal of Clinical Nutrition (2014)
