
Abstract
Bursting bubbles at the ocean-surface produce airborne salt-water spray-droplets, in turn, forming climate-cooling marine haze and cloud layers. The reflectance and ultimate cooling effect of these layers is determined by the spray’s water-uptake properties that are modified through entrainment of ocean-surface organic matter (OM) into the airborne droplets. We present new results illustrating a clear dependence of OM mass-fraction enrichment in sea spray (OMss) on both phytoplankton-biomass, determined from Chlorophyll-a (Chl-a) and Net Primary Productivity (NPP). The correlation coefficient for OMss as a function of Chl-a increased form 0.67 on a daily timescale to 0.85 on a monthly timescale. An even stronger correlation was found as a function of NPP, increasing to 0.93 on a monthly timescale. We suggest the observed dependence is through the demise of the bloom, driven by nanoscale biological processes (such as viral infections), releasing large quantities of transferable OM comprising cell debris, exudates and other colloidal materials. This OM, through aggregation processes, leads to enrichment in sea-spray, thus demonstrating an important coupling between biologically-driven plankton bloom termination, marine productivity and sea-spray modification with potentially significant climate impacts.
Similar content being viewed by others

Introduction
The marine aerosol produces haze and cloud layers overlying an immense ocean covering >70% of the Earth’s surface. Small changes even in low-albedo layers superimposing this relatively dark surface can have profound effects on the global radiation budget and climate change. Organic matter mass-fraction enrichment in sea spray aerosol (OMss, defined here as the percentage OM mass in sea spray relative to the total OM plus sea salt mass) influences the global albedo through altering the reflectance of marine haze1 and cloud layers2. Recent results3 assert that a relatively constant sea surface carbon pool controls OMss rather than biomass abundance or biological productivity and that the constant pool leads to a constant sub-micron sea spray aerosol OMss of the order of 10%. OM in marine aerosol significantly alters the albedo of these layers due to its influence on hygroscopic properties4. Recent studies lead to contrasting perspectives on the relative contribution of water-soluble OM (WSOM) and water-insoluble OM (WIOM) to OMss in primary-produced sea spray aerosol5 from bubble-bursting process and its ultimate cloud droplet nucleating properties6,7.
Bubble-mediated spray production studies in a ship borne laboratory cruising the northeast Atlantic during a phytoplankton bloom, revealed WIOM enrichment up to 80% in submicron sea spray aerosol mass8 while other studies using laboratory-cultures of microalgae7 report enrichments of less than 30%. Analysis of enrichment fractions as a function of biological activity, using changes in chlorophyll-a (Chl-a) concentration as a surrogate of surface photosynthetic primary production, also lead to conflicting results. Satellite measurements of Chl-a9,10 yielded a significant correlation between enrichment and Chl-a and hence biological activity, leading to enrichment parameterisations being developed for global models; however, other studies3 relating instantaneous Chl-a to OMss found no relationship, reporting that the organic carbon content of freshly emitted sea spray aerosol was effectively invariant at 5%, despite significant variability in seawater Chl-a levels. The former studies were conducted over the northeast Atlantic while the latter were conducted in the northwest Atlantic seas to the southwest of Massachusetts and the Pacific California. In the latter study, they3 found that the chemical composition of sea spray organics remained invariant from low-to-high chlorophyll conditions. Based on the above results, it was concluded that oceanic primary organic aerosol production is regulated by the static Dissolved Organic Carbon (DOC) reservoir and consequently, OMss and cloud nucleation activity of nascent sea spray aerosol is relatively constant over the global ocean.
Results
We conducted two types of experiments: one producing a multi-year dataset spanning the period from 1st January 2009 till 30th September 2011 and covering three phytoplankton bloom periods while operating a continuous measurement programme of OM, sea salt and OMss in conjunction with a daily-timescale reanalysis dataset of satellite-derived biological proxy outputs (i.e. Chl-a and Net Primary Productivity - NPP, – see methods section); and a second type comprising discrete bubble-bursting experiments in laboratory tanks using water cultured in the E. huxleyi phytoplankton species, one of the most abundant species in the NE Atlantic. Both experiments where undertaken at the Mace Head atmospheric research station11, a northeast Atlantic World Meteorological Organisation Global Atmospheric Watch station which allowed the unique opportunity to gather the single-most extensive dataset on OMss enrichment in sea spray aerosol, carried in air advecting from highly productive oceanic waters, using state-of-the-art aerosol mass spectrometry. OM and sea salt concentrations were derived from High Resolution Time of Flight Aerosol Mass Spectrometry (AMS) measurements. Standard measurement and calibration procedures12,13 were applied for the OM measurements, with the modified analysis to derive sea salt concentration14. Sea spray enrichment factor was calculated as a ratio of the organic matter and the total sea spray. For the bubble-mediated mesocosm experiments, phytoplankton species were cultured in low-bacteria natural sea water utilising bubble-mediated spray production tanks similar to those used in previous ship-borne experiments amidst plankton blooms8.
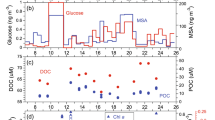
In both ambient air experiments and bubble-tank experiments, we observed the occurrence of striking OM ‘enrichment bursts’ (Fig. 1). During these bursts, the absolute ambient submicron OM mass concentration increased 10-fold and exceeded ~4 μg m−3 while OMss exceeded 95%. Similarly, in the bubble-tank culture experiments, during the OM bursts, OMss exceeded 95% and mass concentrations, scaled to ambient air particle number concentrations, also approached an equivalent concentration of 4 μg m−3 (actual concentrations were more than 20 μg m−3). For the ambient enrichment bursts, environmental and biological proxy data were comparable with marginally higher Chl-a concentrations and marginally-lower wind speeds (see Figure legend) while in the mesocosm experiments, phytoplankton abundance, Chl-a, Dissolved Organic Matter (DOM) and Particulate Organic Matter (POM) were all significantly higher in the non-explosion case.

Left column and Right column plots relate to low OMss experiments and high OMss ‘explosions’, respectively.
(Top row) OM and sea salt mass concentration and OMss for ambient air events. These events are 10 days apart for marine air during August 2009 under conditions of similar biological activity, using Chl-a as an indicator and sea spray production as quantified by wind speed: for the low enrichment event, Chl-a was 0.82 μg L−1 and wind speed was 14 m s−1 while for the high enrichment case, Chl-a was 0.95 μg L−1 and wind speed was 11 m s−1. (2nd Row) OM and sea salt mass concentration and OMss for mesocosm experiments. For the low enrichment mesocosm experiment, cell density, Chl-a, DOC and POC were 108 mL−1, 20.6 μg L−1, 111 μM and 332 μM, respectively while for the high enrichment mesocosm experiment, cell density, Chl-a, DOC and POC were 105 cells mL−1, 5 μg L−1, 140 μM and 21 μM, respectively. (3rd Row) Percentage OM and sea salt as a function of spray particle size sampled by the Berner Impactor during the mesocosm experiments. (Bottom Row) Hygroscopic Growth Factor (GF) frequency distribution for mesocosm experiments. GF is the increase in particle size as the carrier air relative humidity (RH) is increased from <10% RH to 90% RH. For pure sea salt, the GF is 2.2 while GF for pure POM is 1.1. The figure illustrates that for the low enrichment experiment, the particles are predominantly sea salt with a smaller fraction of OM leading to a GF slightly lower than pure sea salt (i.e. 1.85 for the larger sized particles and 1.6 for the smallest). For the high enrichment experiment, where the percentage mass contribution increases with particle size, a more dominant very hydrophobic mode at GF = 1.1 becomes more prominent.
Also shown is the chemical composition (OM and sea salt) measured in four size ranges sampled using a Cascade Impactor15 covering 60 nm to 1 μm size and the hygroscopic growth factor (GF-see methods) for 35 nm, 50 nm, 75 nm, 110 nm and 165 nm particles for the two selected bubble-tank experiments. In the impactor data, without the enrichment explosion, OMss is between 3% and 18% over the four stages whereas, for the enrichment explosion case, the enrichment increases from 25% at the largest sizes to more than 90% at the smallest sizes. A similar pattern in hygroscopic growth is seen with a monomodal frequency distribution of GF centred on 1.85 (very hygroscopic) reflecting an internal mix of predominantly sea salt and a minority of OM matter for the case of low enrichment and for high enrichment, the profound shift in GF to 1.1 (i.e. barely hygroscopic) as particle size reduces and enrichment increases are seen.
The significance of these experiments is that they demonstrate, for the first time, that the OMss bursts occur both in ambient air experiments and laboratory experiments and not only do they reaffirm that OMss contributions can reach 95% submicron mass while simultaneously leading to ambient air OM mass contributions 10-fold higher than other studies reported, but they also show approximately an order of magnitude variability in OMss , again in stark contrast to the Quinn et al.3 study. Perhaps even more striking is that the bursts occur in a pattern independent of instantaneous productivity markers and wind-generated forcing, both for the laboratory and ambient air experiments. While the lack of correlation between OMss and instantaneous productivity markers is in agreement with the findings of Quinn et al.3, both the large absolute OM concentration, variation in OMss and magnitude of OMss is in stark contrast to their results.
To explore further any correlation between enrichment, phytoplankton biomass (as Chl-a concentrations) and productivity (as NPP), the multi-year dataset was analysed for a selected region (between 47°N- 57°N and 14°W-24°W, see Fig. 2) upwind of Mace Head. The cleanest air mass with least continental and/or anthropogenic contamination encountered in the region is a maritime polar air mass, therefore, only these air masses were selected for the sea spray composition analysis. An additional black carbon criterion of 1 hour average mass concentration not exceeding 15 ng m−3 was applied to the data in order to ensure the cleanliness of the marine air masses. Chl-a averages over the selected region were filtered for the same periods that corresponded to these clean air criteria. The air mass characterisation was conducted on a 6 hour time resolution and, consequently, the enrichment factor was averaged over the 1 hour period around the air mass classification. Since the highest Chl-a time resolution was 24 hours, its concentration was assumed to be constant for that particular day. Also shown in the Fig. 2 is the correlation coefficient between Chl-a and OMss with an 8-day time lag, illustrating the regions of biomass abundance strongly correlated with enrichment measured at Mace Head. An 8-day time-lag was previously found10 to lead to a maximum correlation between Chl-a and OMss and was regarded10 as a biological time-lag between peak biomass abundance and peak OMss in spray.

The correlation map of sea spray enrichment, measured at Mace Head and Chl-a over the North East Atlantic.
Pixel size is 0.042° (~5 km). Correlation was calculated from daily mass-fraction enrichment and Chl-a values for the period of 2009–2011. Only clean maritime polar air masses were taken into account. The 8 day lag between the enrichment and Chl-a was used in this analysis. Grey box represents an area of 47°N-57°N and 14°W-24°W10. The map was created by IGOR Pro 6.
The magnitude and variation of OMss is also clearly seen in the analysis of the 3 year dataset as displayed in Fig. 3 where OMss values increase from 4% in winter to greater than 90% in summer as Chl-a increased from 0.2 mg m−3 to 0.96 mg m−3 and NPP from 161 mgC m−2 d−1 to 1234 mgC m−2 d−1. Of course, the OM fraction derived from AMS measurements would include secondary OM contribution as well. So far, there is no reliable method to routinely separate primary and secondary marine aerosol OM, since they could bear the same properties, especially, after aging16. Nevertheless, even high secondary contribution would not change the whole picture very much. By way of illustrations, during the warm, sunny, period of summer, where high secondary contribution could be expected, an extraordinly high 70% classification of OM as secondary organic aerosol would result in only ~15% reduction in OMSS, as high OMSS in such periods is determined by contextually-high OM and low sea salt concentrations. OM variations could have larger impact on low OMSS values occurring during the winter period. Nevertheless, the contribution from secondary OM during winter is expected to be minumum, resulting eventually in insignificant effect on OMss. In other words, secondary contribution could scale down the OMss by no more than 15% over all seasons, leaving the observed yearly trend unchanged. This is illustrated in Fig. 3.

Temporal seasonality of OMss in sea spray aerosol, sea water chlorophyll-a (Chl-a) and Net Primary Production (NPP).
OMss bold lines represent median concentrations, boxes- 25–75% percentile and whiskers demonstrate 10–90% percentile. Grey area represents a reduction in primary OMss due to possible secondary OM contribution to total organics. The secondary OM contribution was derived from data presented in Ceburnis et al.55 and ranges from 22% to 74% depending on the month.
The shortest timescale analysed using the satellite data was one day and led to a correlation coefficient between Chl-a and OMss of R = 0.67 (Fig. 4) while no correlation in instantaneous timescales was observed from the bubble-tank experiments. Moreover, Fig. 4 illustrates that as the timescale was lengthened to one week, the correlation increased to 0.76 and up to 0.85 for a month-long timescale while NPP exhibited a correlation of 0.93, again for a month–long timescale, confirming that OMss is neither constant nor independent of biological activity.

(a) Scatter plot of sea spray OMss as a function of Chl-a for daily, weekly and monthly temporal resolutions and NPP for monthly temporal resolution; (b) correlation coefficient as a function of temporal resolution of biological indicators.
The aforementioned correlation maximum for monthly average concentrations, combined with the observations of a 26-day “boom-bust” biological cycle timescale associated with bloom dynamics as recently illustrated by Lehahn et al.17, suggests that the maximum correlation occurs at a timescale longer than 8 days and perhaps the peak in OMss is more associated with the demise of the bloom rather than any growth phase.
This suggestion is supported here through examination of the correlation coefficient frequency function for NPP, taken over the grid region from 44° and 50° N and −10° and −29°W (the area overwhich trajectories advect during the strongest OM plumes) for time lags of 0, 8, 16 and 24 days. Figure 5 illustrates the correlation between NPP and OMss over the chosen grid and illustrates that the correlation coefficient distribution function both narrows and increases in both amplitude and peak distribution value as the time lag is increased from 0 to 16–24 days. This suggests a maximum correlation between OMss and NPP for the latter third of a typical bloom life span, or, in other words, maximum correlation is seen for bloom demise rather than bloom decay. The width of the distribution function and the lack of a mean correlation as high as that seen for averages over weekly and monthly timescales shown in Figs 3 and 4 is most likely due to blooms, typically active at the meso-scale of approximately 200 km in size16, waxing and waning out of synchronization with each other.

Correlation coefficient spatial distribution between OMss and NPP for 0 days time lag (left) and for 24 days time lag (middle) at each grid cell upwind of Mace Head.
The correlation coefficient frequency distribution is also shown (right) for area comprising grid coordinates 44° and 50° N and −10° and −30°W (the area overwhich trajectories advect during the strongest OM plumes) for time lags of 0, 8, 16 and 24 days. The maps were created by IGOR Pro 6.
Discussion
Both theoretical and experimental studies suggest6,8,18,19,20,21 that OMss in sea spray is driven by insoluble gel-colloids forming material characterized by surface-active properties and nanosized particles (<200 nm), which are components of DOM, such as viruses and cellular fragments due to cell death or cell burst induced by viral infection. This means that self-aggregation, forming insoluble surfactants, is the key process driving OMss in sea spray. DOM and POM in the ocean are in a dynamic equilibrium as the enrichment in DOM induces aggregation processes leading to the production of POM, while particle dissolution by extracellular enzymatic activities determines the shift from POM to DOM22. The aggregation properties of marine OM can be due to a number of factors, including presence of metals, dynamic conditions of the oceans, UV irradiation and chemical composition23. Moreover, the role of prokaryotic activity and cell lysis in forming such colloidal gel-forming material is proposed in several studies24,25,26. Indeed, our bubble bursting experiments support the above suggestions through revealing enrichment factors in fine mode aerosol of the order of 200 for viruses, 30 for prokrayotes, 100,000 for lipids, carbohydrates and proteins and 15,000 for DNA. The enrichment factor in the fine, or submicron sea-spray was typically more than 10-fold that in the coarse mode.
While these processes are poorly understood at present, increasing evidence indicates that biological processes determining cell lysis, including viral infections could play a pivotal role in the process of transformation of living biomass into cell debris and in DOM dynamics (e.g. cytoplasmic material)17,27,28,29,30. Among biological processes responsible for the release of cell debris and DOM from phytoplankton blooms, grazing and viral infections can play a dominant role30,31. Marine phytoplankton blooms generally last for a few weeks and these biological processes can be responsible for the abatement of 50–100% of the phytoplankton. Zooplankton during grazing on phytoplankton, by sloppy feeding, can release approximately 30% of the ingested biomass as DOM.
Viruses play an important role in both global and small-scale biogeochemical cycling, influencing community structure and phytoplankton bloom termination. Viruses are indeed the most numerous ‘lifeforms’ in aquatic systems, at least 10 times the total number of Bacteria and Archaea and an abundance in marine waters ranging from about 108 to 1010 L−1. Given that the vast majority of the biomass in oceans comprises microorganisms, it is expected that viruses and other prokaryotic and eukaryotic microbes will play important roles in biogeochemical processes.
Viral infections in marine plankton can be lysogenic (viral genomes integrated into the genomes of their hosts) or lytic (causing host cell bursting), frequently followed by host death. Thus, lytic infections represent an important source of mortality of marine plankton.
Most marine viruses infect primarily prokaryotes, followed by microalgae, which are among the most abundant eukaryotic organisms in the ocean27,32. Virus-mediated mortality of bacterioplankton and phytoplankton is often in the range of 10–30%, but can reach 100%33,34. Epidemiological models predict that viral infection rates increase with increasing host cell density because infection is a direct function of the encounter rate between a pathogen and its host and it is thought that host density drives much of the lytic response. For this reason, available studies provided evidence that viral abundance and the importance of viruses in infecting and killing their hosts increases with the productivity of water masses, with the highest percentage of infected cells occurring in highly eutrophic ecosystems35. Phytoplankon bloom collapse occurs when viral lysis rates exceed the specific host growth rates. This type of interaction has been referred to as control by “reduction30”. Compelling evidence that viruses are involved in algal bloom demise comes from observations of high proportions (10–50%) of cells being visibly infected (using TEM) at the end of Aureococcus anophagefferens36, Heterosigma akashiwo37 and Emiliania huxleyi38 blooms. Further evidence for viral control of phytoplankton comes from dividing empirical estimates of virus production by burst size, which indicates that viral lysis can be responsible for 7 to 100% of the mortality of Phaeocystis globosa39 and E. huxleyi40 during the bloom events. When viral infections of phytoplankton provoke bursting of infected cells, their cellular content is released30,41, resulting in sub-micron sized particles and colloids (e.g., cell wall fragments and intracellular organic compounds) being released into surface waters27,33. Phytoplankton blooms are also known to produce exopolymeric substances30, which add to the variable and diverse OM pool, as a response to stress.
Wind-driven dynamics, resulting in bubble generation, is perhaps the most effective way of promoting self-aggregation and the formation of ocean surface foam-surfactant layers leading to OMss. The significant difference between our results and those reporting low and invariant enrichment controlled by the DOC reservoir3 could be due to lack of wind-driven dynamics in those experiments. The difference could also be, in part, species driven: in the blooming northeast Atlantic waters, characterized by lower Chl-a and similar POC and DOC concentrations (1.4 ± 0.8 μg L−1, 12 ± 6 μM C, 54 ± 20 μM C, respectively) compared to the northwest Atlantic and Pacific waters , OMss, variable and far greater than 35% was encountered, pointing to the possibility that the very low and constant enrichment observed3 in regions of very high Chl-a concentration could be due to the production of OM which is lacking in aggregating surface-active properties. Such a scenario could arise, for instance, as a result of a lack of significant viral infections and the subsequent release of colloidal material and cell debris.
The increased correlation between OMss and biological activity indicators, as the timescale is increased, supports the concept that the production of OM and its mass fraction enrichment in spray is, indeed, strongly correlated to the degree of surface water biological activity; however, the production of OM appears to be more linked to the bloom’s decay and cell lysis rather that life cycle – the waning of prokaryotes (bacteria and archaea) and phytoplankton due to viral infections after the waxing stage. In particular, these results are consistent with the time lag between the initiation of the phytoplankton bloom (generally assumed to occur when Chl-a concentrations pass the threshold of 1 μg L−1), the increase of prokaryotes that can exploit the phytoplankton exudates and the ability of viruses to infect living cells33,42. These aspects could contribute to explain why the phytoplankton biomass (here measured through the proxy Chl-a concentration) in surface waters is a pre-requisite for the release of sub-micron organic particles, mainly water insoluble, in the aerosol, but are not sufficient to determine the process directly. Aggregated biogenic materials are evident in both sea spray and filtered water images; however, it is predominantly fine (<10 μm) POC that contributes to sea spray OMss as evidenced by 1H-Nuclear Magnetic Resonance (1HNMR) fingerprinting of both fine POC and OM samples8. Colloids and nanogels, which are sufficiently small to enter the DOC operational definition (organic matter small enough in size to pass through a 0.2 μm pore filter) contribute to sea spray enrichment as well. We hypothesise that the viruses, which have been reported to increase exponentially along with the waning of E. huxleyi in the experimental systems show a size ranging from 120 to 180 nm and contribute to the DOC pool thought they are evidently submicron particles while the rest of the dissolved OM pool is less likely to contribute to enrichment in sea spray aerosol. The identification of the biological processes involved in the termination of the phytoplankton blooms, including the impact of viruses, is crucial to quantify their role in DOM dynamics. Further investigations are needed to provide mechanistic evidence of the interactions amongst these biotic components, able to provide production “pulses” of living and dead biomass. The bloom demise, or its death disco, not only forms an essential component of the climate-mediated plankton dance43, but also plays an essential role in the organic carbon cycle.
Methods
Aerosol Measurements
Aerosol chemical composition was measured with the Aerodyne Research Inc. high resolution time-of-flight aerosol mass spectrometer (AMS) which provides real-time size resolved composition analysis of volatile and semi-volatile particulate matter12. The combination of size and chemical analysis of PM 1.0 aerosol mass loading with fast time resolution makes the AMS unique and the theory of operation is well established. In summary, the AMS quantifies non-refractory aerosol chemical composition, covering major inorganic species such as ammonium, sulphate, nitrate, plus organic species13. In addition, recent work has demonstrated that both sea salt mass and primary marine organic mass can be retrieved from the high resolution AMS14. GF properties of aerosol were measured using a Hygroscopic Tandem Differential Mobility Analyser (H-TDMA)44. A higher GF indicates more hygroscopic particles resulting from a higher affinity for water. The Mace Head H-TDMA covered a “dry-size” RH of 20% to a humidified RH of 90%. GFs were measured for particle sizes of 35, 50, 75, 110 and 165 nm. An inversion algorithm was used to retrieve the GF Probability Density Function (PDF), adjusted for multiple charging effects and other required associated corrections45.
Bubble-mediated experiments
A series of bubble-mediated aerosol production experiments were carried out using different cell densities and different stages of the of the microalgal species Emiliania huxleyi growth phase was examined. The cell densities chosen were typical of the abundance associated with natural bloom conditions. During each experiment, the cell number and viability was tested periodically; cell number was noted at the start of the experiment using a haemocytometer. The numbers of dead cells were counted during or at the end of the experiment by adding 2–3 drops of 0.1% Evan’s blue dye to 20 mL of the sample46. Samples were also collected for Chl-a (HPLC), nutrient and DOC and POC analysis. WSOM and WIOM for selected experiments were also analysed by 1HNMR analysis.
In the experiments, an airtight high grade stainless steel tank (200 L in volume) was filled with 100 L of filtered and UV-sterilised seawater. This seawater was freshly collected close to Mace Head in Carna Bay, Co. Galway, on the day each experiment. Samples were taken from approximately 30 m off-shore. Light was provided by cool white fluorescent lamps (L 18W/865 lumilux fluorescence lamps, Osram, Germany) fitted above the water surface inside the tank which provided the phytoplankton cultures a constant light of 80 μmol photons m−2 s−1 (measured using a Li Cor 1400 data logger) at 12 h:12 h photoperiod. Air was entrained into water from the top of the tank by a vertical re-circulating water jet at a flow rate of 20 L min−1. The entrained air formed bubbles that in turn burst at the surface producing sea spray aerosol.
The tank had several ports on the top where different analytical instruments were connected: (1) an AMS, which measured sea spray chemical composition; (2) two 8 stage Berner impactors mounted for approximately 6 hours (0.060–16 μm diameter), with different sampling substrates (quartz and tedlar) in order to analyse the organic and inorganic components; (3) a Scanning Mobility Particle Sizer (SMPS), which measured aerosol size distributions (10–500 nm); (4) a Humidified Tandem Differential Mobility Analyser (HTDMA), which measured aerosol growth factor (35, 50, 75, 110 and 165 nm).
Each experiment was carried out for up to several days until after the phytoplankton culture was declared dead. Two experiments were specifically designed to study algae-virus interaction using Emiliania huxleyi (CCMP 1516) and cocolithovirus EhV-86 with corresponding densities of 105 cells ml−1 and 104 viruses ml−1 which were cultured in separate batches and inoculated 4 hours before being transferred into experimental tank. A separate control tank was used in algae-virus experiments using same phytoplankton culture and density as in experimental tank, but without inoculation with viruses.
Description of the Satellite Products
Daily satellite sea surface Chl-a concentration data were acquired from MyOcean47. The global Chl-a dataset at 4 km of resolution produced by the ESA OC-CCI Project was used and is a climate-quality consistent dataset produced using the latest and most complete knowledge of satellite sensor calibration, characterization and attitude, complete ancillary data sets, latest versions of models and algorithms. SeaWiFS, MODIS and MERIS Remote sensing reflectances were corrected for atmospheric transmission, band shifted to SeaWiFS wavebands and merged. Chl-a concentration were calculated applying the NASA OC4.V6 algorithm on the OC-CCI merged remote sensing reflectances48. Standard masking criteria for detecting clouds or other contamination factors were applied to the sensor data, i.e., land, cloud, sun glint, atmospheric correction failure, high total radiance, large solar zenith angle (70°), large spacecraft zenith angle (56°), coccolithophores, negative water leaving radiance and normalized water leaving radiance at 555 nm 0.15 Wm−2 sr−1 49. The quality of chlorophyll product (mg m−3) was evaluated by comparing the satellite retrieval with corresponding in situ data. The analysis, performed on the log10 transformed data, results in a bias of −0.01910 and an RMS 0.30300 (see MyOcean Quality Information Document: MYO2-OC-QUID-009–056–057–058–059-V2.050).
From this global dataset we extracted the chlorophyll field our study area (29°-7°W; 44°-60°N) from 1st January 2009 to 31st December 2011. Multi-channel singular spectral analysis (M-SSA) was used to fill daily gaps in the chlorophyll maps due to cloud cover or other environmental factors. The method uses temporal as well as spatial correlation to fill the missing points (see Appendix A of Rinaldi et al.10 for method details).
The NPP dataset used in this paper were acquired from the Ocean Productivity webpage51. We selected the PP Standard product computed using the Vertically Generalized Production Model (VGPM)52. For the VGPM, net primary production is a function of chlorophyll, available light and the photosynthetic efficiency. This Standard Product is computed using MODIS chlorophyll and temperature data, SeaWiFS PAR and estimates of euphotic zone depth from a model developed by Morel and Berthon53 and based on chlorophyll concentration.
Analyses of viruses and prokaryotes in the seawater and marine aerosol
Seawater, samples were filtered onto Anodisc 25 mm (20 nm pore size) filters. To determine viral and prokaryotic abundance in the aerosol, each size class quartz filter (collected for approximately 3–12 h during bubble bursting experiments by using an 80 L min−1 five stage Berner impactor in the size range 0.05–10 μm) was homogenised in sterile and virus free MilliQ water (pre-filtered onto 0.02 μm pore-size filters) and samples were then treated by ultrasounds and pyrophosphate (5 mM final concentration) to detach viruses and prokaryotes. Aerosol samples were then centrifuged at 800 g for 10 minutes and the supernatant filtered onto Anodisc 25 mm (20 nm pore size) filters. All Anodisc filters were then stained with 20 μl of SYBR Green I (diluted 20 fold in MilliQ water, optical density at 495 nm ca. 1.3) for 15 minutes in the dark, rinsed twice with 1 ml MilliQ water (in order to eliminate fluorescence background noise). The filters were mounted on slides using an anti-fade solution (50% phosphate buffer [6.7 mM, pH 7.8] and 50% glycerol containing 0.25% ascorbic acid) and analyzed by epifluorescence microscopy using a Zeiss Axioplan microscope, equipped with a 50W lamp. To reduce the uncertainties on viral and prokaryotic counts due to the low abundance in aerosol samples, at least 100 optical fields at 1000× magnification were examined.
Analyses of carbohydrates, proteins, lipids and DNA in the seawater and marine aerosol
Seawater samples were pre-filtered onto 10 μm pore-size filters and then onto polycarbonate 0.2 μm pore-size filters. Each size class quartz filter used for collecting aerosol during bubble bursting experiments was homogenised in sterile and virus free MilliQ water (pre-filtered onto 0.02 μm pore-size filters) and samples were then treated with ultrasound. Carbohydrate, protein and lipid determinations, both in aerosol and seawater samples, were carried out spectrophotometrically according to procedures previously described54. The analyses of total DNA in aerosol and seawater samples were performed fluorometrically using SYBR Green I as fluorochrome after digestion of RNA with RNase (DNase free, 1 U mL−1 for 15 minutes at room temperature). Carbohydrate, protein, lipid and DNA concentrations were obtained by calibration curves using standard solutions of glucose (1–25 μg mL−1), bovine serum albumin (0.5–10 μg mL−1), tripalmitin (2.5–25 μg mL−1) and DNA from E. coli (1–100 ng mL−1), respectively. The detection limits were 2.5 μg mL−1, 1 μg mL−1, 5 μg mL−1 and 2.5 ng mL−1 for carbohydrates, proteins, lipids and DNA, respectively. Clean quartz and polycarbonate filters were used as blanks. Carbohydrate, protein, lipid and DNA concentrations obtained from the analyses of 0.05–1.2 μm pore size filters were summed and defined as belonging to the fine fraction (<1.2 μm) of the aerosol, whereas concentrations obtained from the analyses of 1.2–10 μm pore size filters were defined as belonging to the coarse fraction (>1.2 μm).
Additional Information
How to cite this article: O’Dowd, C. et al. Connecting marine productivity to sea-spray via nanoscale biological processes: Phytoplankton Dance or Death Disco? Sci. Rep. 5, 14883; doi: 10.1038/srep14883 (2015).
References
Vaishya, A. et al. Bistable Effect of Organic Enrichment on Sea-Spray Radiative Properties. Geophys Res Lett. 2013GL058452, 10.1002/2013GL058452 (2013).
Latham, J. & Smith, M. H. Effect on global warming of wind-dependent aerosol generation at the ocean surface. Nature 347, 372–373, 10.1038/347372a0 (1990).
Quinn, P. K. et al. Contribution of sea surface carbon pool to organic matter enrichment in sea spray aerosol. Nature Geosci 7, 228–232, 10.1038/ngeo2092 (2014).
O’Dowd, C. D. et al. Biogenically driven organic contribution to marine aerosol. Nature 431, 676–680, 10.1038/Nature02959 (2004).
Keene, W. C. et al. Chemical and physical characteristics of nascent aerosols produced by bursting bubbles at a model air-sea interface. J Geophys Res-Atmos 112, D21202, 10.1029/2007jd008464 (2007).
Ovadnevaite, J. et al. Primary marine organic aerosol: A dichotomy of low hygroscopicity and high CCN activity. Geophys Res Lett 38, 10.1029/2011gl048869 (2011).
Fuentes, E., Coe, H., Green, D. & McFiggans, G. On the impacts of phytoplankton-derived organic matter on the properties of the primary marine aerosol – Part 2: Composition, hygroscopicity and cloud condensation activity. Atmos. Chem. Phys. 11, 2585–2602, 10.5194/acp-11-2585-2011 (2011).
Facchini, M. C. et al. Primary submicron marine aerosol dominated by insoluble organic colloids and aggregates. Geophys Res Lett 35, L17814, 10.1029/2008gl034210 (2008).
O’Dowd, C. D. et al. A combined organic-inorganic sea-spray source function. Geophys Res Lett 35, 10.1029/2007gl030331 (2008).
Rinaldi, M. et al. Is chlorophyll-a the best surrogate for organic matter enrichment in submicron primary marine aerosol? J Geophys Res-Atmos 118, 4964–4973, 10.1002/jgrd.50417 (2013).
O’Connor, T. C., Jennings, S. G. & O’Dowd, C. D. Highlights of fifty years of atmospheric aerosol research at Mace Head. Atmos Res 90, 338–355, 10.1016/j.atmosres.2008.08.014 (2008).
DeCarlo, P. F. et al. Field-deployable, high-resolution, time-of-flight aerosol mass spectrometer. Anal Chem 78, 8281–8289, 10.1021/Ac061249n (2006).
Jimenez, J. L. et al. Ambient aerosol sampling using the Aerodyne Aerosol Mass Spectrometer. J Geophys Res-Atmos 108, 8425, 10.1029/2001JD001213 (2003).
Ovadnevaite, J. et al. On the effect of wind speed on submicron sea salt mass concentrations and source fluxes. J Geophys Res-Atmos 117, 11, 10.1029/2011jd017379 (2012).
Berner, A., Lürzer, C., Pohl, F., Preining, O. & Wagner, P. The size distribution of the urban aerosol in Vienna. Sci Total Environ 13, 245–261 (1979).
Jimenez, J. L. et al. Evolution of Organic Aerosols in the Atmosphere. Science 326, 1525–1529, 10.1126/science.1180353 (2009).
Lehahn, Y. et al. Decoupling Physical from Biological Processes to Assess the Impact of Viruses on a Mesoscale Algal Bloom. Curr Biol 24, 2041–2046, 10.1016/j.cub.2014.07.046 (2014).
Leck, C. & Bigg, E. K. Source and evolution of the marine aerosol - A new perspective. Geophys Res Lett 32, 10.1029/2005gl023651 (2005).
Li, X. et al. Cross-Linked Polysaccharide Assemblies in Marine Gels: An Atomistic Simulation. J. Phys. Chem. Lett. 4, 2637–2642, 10.1021/jz401276r (2013).
Orellana, M. V. et al. Marine microgels as a source of cloud condensation nuclei in the high Arctic. Proc. Natl. Acad. Sci. USA 108, 13612–13617, 10.1073/pnas.1102457108 (2011).
Prather, K. A. et al. Bringing the ocean into the laboratory to probe the chemical complexity of sea spray aerosol. Proc. Natl. Acad. Sci. USA 110, 7550–7555, 10.1073/pnas.1300262110 (2013).
Smith, D. C., Simon, M., Alldredge, A. L. & Azam, F. Intense hydrolytic enzyme activity on marine aggregates and implications for rapid particle dissolution. Nature 359, 139–142, 10.1038/359139a0 (1992).
Verdugo, P. Marine Microgels. Ann Rev Mar Sci 4, 375–400, 10.1146/annurev-marine-120709-142759 (2012).
Simon, M., Grossart, H.-P., Schweitzer, B. & Ploug, H. Microbial ecology of organic aggregates in aquatic ecosystems. Aquat Microb Ecol 28, 175–211, 10.3354/ame028175 (2002).
Jiao, N. et al. Microbial production of recalcitrant dissolved organic matter: long-term carbon storage in the global ocean. Nat Rev Micro 8, 593–599, 10.1038/nrmicro2386 (2010).
Lønborg, C., Middelboe, M. & Brussaard, C. D. Viral lysis of Micromonas pusilla: impacts on dissolved organic matter production and composition. Biogeochemistry 116, 231–240, 10.1007/s10533-013-9853-1 (2013).
Fuhrman, J. A. Marine viruses and their biogeochemical and ecological effects. Nature 399, 541–548, 10.1038/21119 (1999).
Suttle, C. A. Marine viruses - major players in the global ecosystem. Nat Rev Micro 5, 801–812, 10.1038/nrmicro1750 (2007).
Danovaro, R. et al. Major viral impact on the functioning of benthic deep-sea ecosystems. Nature 454, 1084–1087, 10.1038/nature07268 (2008).
Brussaard, C. P. D. Viral Control of Phytoplankton Populations—a Review 1. J. Eukaryot. Microbiol. 51, 125–138, 10.1111/j.1550-7408.2004.tb00537.x (2004).
Brussaard, C. P. D. et al. Global-scale processes with a nanoscale drive: the role of marine viruses. ISME J 2, 575–578 (2008).
Weinbauer, M. G. Ecology of prokaryotic viruses. Fems Microbiol Rev 28, 127–181, 10.1016/j.femsre.2003.08.001 (2004).
Wommack, K. E. & Colwell, R. R. Virioplankton: Viruses in Aquatic Ecosystems. Microbiol. Mol. Biol. Rev. 64, 69–114, 10.1128/mmbr.64.1.69-114.2000 (2000).
Corinaldesi, C., Dell’Anno, A. & Danovaro, R. Viral infection plays a key role in extracellular DNA dynamics in marine anoxic systems. Limnol Oceanogr 52, 508–516, 10.4319/lo.2007.52.2.0508 (2007).
Weinbauer, M. G. & Rassoulzadegan, F. Are viruses driving microbial diversification and diversity? Environ Microbiol 6, 1–11, 10.1046/j.1462-2920.2003.00539.x (2004).
Gastrich, M. D. et al. Assessment of brown tide blooms, caused by Aureococcus anophagefferens and contributing factors in New Jersey coastal bays: 2000–2002. Harmful Algae 3, 305–320, 10.1016/j.hal.2004.06.003 (2004).
Lawrence, J. E., C. A. M. & Suttle, C. A. Viruses causing lysis of the toxic bloom-forming alga Heterosigma akashiwo (Raphydophyceae) are widespread in coastal sediments of British Columbia, Canada. Limnol Oceanogr 47, 545–550 (2002).
Bratbak, G., Jacobsen, A. & Heldal, M. Viral lysis of Phaeocystis pouchetii and bacterial secondary production. Aquat Microb Ecol 16, 11–16, 10.3354/ame016011 (1998).
Brussaard, C. P. D., Kuipers, B. & Veldhuis, M. J. W. A mesocosm study of Phaeocystis globosa population dynamics: I. Regulatory role of viruses in bloom control. Harmful Algae 4, 859–874, 10.1016/j.hal.2004.12.015 (2005).
Jacquet, S. et al. Flow cytometric analysis of an Emiliana huxleyi bloom terminated by viral infection. Aquat Microb Ecol 27, 111–124, 10.3354/ame027111 (2002).
Bratbak, G., Egge, J. K. & Heldal, M. Viral Mortality of the Marine Alga Emiliania-Huxleyi (Haptophyceae) and Termination of Algal Blooms. Mar Ecol Prog Ser 93, 39–48, 10.3354/Meps093039 (1993).
Ackermann, H. W. & DuBow, M. S. Viruses of Prokaryotes: General properties of bacteriophages. (CRC Press, 1987).
Behrenfeld, M. J. Climate-mediated dance of the plankton. Nature Clim. Change 4, 880–887, 10.1038/nclimate2349 (2014).
Nilsson, E. et al. Development of an H-TDMA for long-term unattended measurement of the hygroscopic properties of atmospheric aerosol particles. Atmos Meas Tech 2, 313–318 (2009).
Gysel, M., McFiggans, G. B. & Coe, H. Inversion of tandem differential mobility analyser (TDMA) measurements. J Aerosol Sci 40, 134–151 (2009).
Andersen, R. A. Algal Culturing Techniques. (Elsevier/Academic Press, 2005).
Marine Environment Monitoring Service. Available at: www.myocean.eu. (Accessed: January 2014).
O’Reilly, J. E. et al. In SeaWiFS Postlaunch Calibration and Validation Analyses, Part 3 Vol. 11 (ed. S.B. Hooker ) (NASA Goddard Space Flight Center, 2000).
McClain, C. R. e. a. SeaWiFS Algorithms, Part 1. NASA Tech. Memo. 104566. (NASA Goddard Space Flight Center, Greenbelt, Maryland, 1995).
Santoleri, R., Melin, F. & Volpe, G. For Ocean Colour Global and European Sea, Observation Product, OC-TAC CNR. Quality Information Document. (2013) Available at: http://catalogue.myocean.eu.org/static/resources/myocean/quid/MYO2-OC-QUID-009-056-057-058-059-V2.0.pdf. (Accessed: May 2015).
Ocean Productivity. Available at: http://www.science.oregonstate.edu/ocean.productivity/. (Accessed: January 2014)
Behrenfeld, M. J. & Falkowski, P. G. Photosynthetic rates derived from satellite-based chlorophyll concentration. Limnol Oceanogr 42, 1–20 (1997).
Morel, A. & Berthon, J. F. Surface Pigments, Algal Biomass Profiles and Potential Production of the Euphotic Layer - Relationships Reinvestigated in View of Remote-Sensing Applications. Limnol Oceanogr 34, 1545–1562 (1989).
Danovaro, R. Methods for the Study of Deep-Sea Sediments, Their Functioning and Biodiversity. (CRC Press Taylor & Francis Group, 2010).
Ceburnis, D. et al. Marine submicron aerosol sources, sinks and chemical fluxes. Atmos. Chem. Phys. Discuss. 14, 23847–23889, 10.5194/acpd-14-23847-2014 (2014).
Acknowledgements
The research leading to these results has received funding from the European Space Agency’s Support To Science Element: Oceanflux Sea Spray Aerosol project under grant agreement 4000104514/11/I-AM; the Environmental Protection Agency’s (Ireland) EASI-AQCIS project (grant agreement n° 2007-CCRP-5.3); and the European Union’s Seventh Framework Programme (FP7/2007-2013) project BACCHUS (grant agreement n° 603445); The work was further supported by the CNR (Italy) under AirSEaLab : Progetto Laboratori Congiunti; the Irish Higher Education Authority under the Programme for Research in Third Level Ireland, 4; and the European Commission Horizon 2020 Framework Programme (grant agreement n° 633085) and by the Copernicus Marine Environment Monitoring Service –CMEMS (grant agreement n° 9836100).
Author information
Authors and Affiliations
Contributions
C.O.D. wrote the paper and led the overall study. D.C. coordinated the bubble-bursting culture experiments and field studies and conceived the virus experiments. J.O. conducted the aerosol mass spectrometer measurements and data integration and analysis, J.B. conducted the GF measurements; M.R. conducted the impactor measurements, S.D. conducted the H+NMR measurements, S.M. & R.S. provided the satellite analysis, D.S., M.Z., U.N. and S.C. cultured the algae and participated in the mesocosm experiments, A.D., C.C. and M.T. analysed water and aerosol samples for viruses and biological components. M.C.F. and S.F. coordinated the aerosol and sea water chemical analysis. R.D. coordinated the virus and bacteria analysis.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
O’Dowd, C., Ceburnis, D., Ovadnevaite, J. et al. Connecting marine productivity to sea-spray via nanoscale biological processes: Phytoplankton Dance or Death Disco?. Sci Rep 5, 14883 (2015). https://doi.org/10.1038/srep14883
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep14883
This article is cited by
-
Virus infection of phytoplankton increases average molar mass and reduces hygroscopicity of aerosolized organic matter
Scientific Reports (2023)
-
Regionally sourced bioaerosols drive high-temperature ice nucleating particles in the Arctic
Nature Communications (2023)
-
A critical review on bioaerosols—dispersal of crop pathogenic microorganisms and their impact on crop yield
Brazilian Journal of Microbiology (2023)
-
Factors controlling the transfer of biogenic organic species from seawater to sea spray aerosol
Scientific Reports (2022)
-
Surface ocean microbiota determine cloud precursors
Scientific Reports (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
