
Abstract
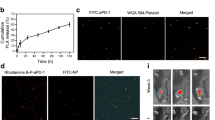
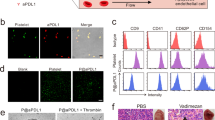
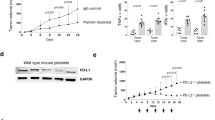
Cancer recurrence after surgical resection remains a significant challenge in cancer therapy. Platelets, which accumulate in wound sites and interact with circulating tumour cells (CTCs), can however trigger inflammation and repair processes in the remaining tumour microenvironment. Inspired by this intrinsic ability of platelets and the clinical success of immune checkpoint inhibitors, here we show that conjugating anti-PDL1 (engineered monoclonal antibodies against programmed-death ligand 1) to the surface of platelets can reduce post-surgical tumour recurrence and metastasis. Using mice bearing partially removed primary melanomas (B16-F10) or triple-negative breast carcinomas (4T1), we found that anti-PDL1 was effectively released on platelet activation by platelet-derived microparticles, and that the administration of platelet-bound anti-PDL1 significantly prolonged overall mouse survival after surgery by reducing the risk of cancer regrowth and metastatic spread. Our findings suggest that engineered platelets can facilitate the delivery of the immunotherapeutic anti-PDL1 to the surgical bed and target CTCs in the bloodstream, thereby potentially improving the objective response rate.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$99.00 per year
only $8.25 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others
References
Baker, D., Masterson, T., Pace, R., Constable, W. & Wanebo, H. The influence of the surgical wound on local tumor recurrence. Surgery 106, 525–532 (1989).
Lukianova-Hleb, E. Y. et al. Intraoperative diagnostics and elimination of residual microtumours with plasmonic nanobubbles. Nat. Nanotech. 11, 525–532 (2016).
Stephan, S. B. et al. Biopolymer implants enhance the efficacy of adoptive T-cell therapy. Nat. Biotechnol. 33, 97–101 (2015).
Demicheli, R., Retsky, M., Hrushesky, W., Baum, M. & Gukas, I. The effects of surgery on tumor growth: a century of investigations. Ann. Oncol. mdn386 (2008).
Ceelen, W., Pattyn, P. & Mareel, M. Surgery, wound healing, and metastasis: recent insights and clinical implications. Crit. Rev. Oncol. Hematol. 89, 16–26 (2014).
Klevorn, L. E. & Teague, R. M. Adapting cancer immunotherapy models for the real world. Trends Immunol. 37, 354–363 (2016).
O’Sullivan, D. & Pearce, E. L. Targeting T cell metabolism for therapy. Trends Immunol. 36, 71–80 (2015).
Robert, C. et al. Pembrolizumab versus ipilimumab in advanced melanoma. N. Engl. J. Med. 372, 2521–2532 (2015).
Postow, M. A. et al. Nivolumab and ipilimumab versus ipilimumab in untreated melanoma. N. Engl. J. Med. 372, 2006–2017 (2015).
Sharma, P. & Allison, J. P. The future of immune checkpoint therapy. Science 348, 56–61 (2015).
Wang, C., Ye, Y., Hochu, G. M., Sadeghifar, H. & Gu, Z. Enhanced cancer immunotherapy by microneedle patch-assisted delivery of anti-PD1 antibody. Nano Lett. 16, 2334–2340 (2016).
Zou, W., Wolchok, J. D. & Chen, L. PD-L1 (B7-H1) and PD-1 pathway blockade for cancer therapy: mechanisms, response biomarkers, and combinations. Sci. Transl. Med. 8, 328rv4 (2016).
Buchbinder, E. I. & Hodi, F. S. Melanoma in 2015: immune-checkpoint blockade—durable cancer control. Nat. Rev. Clin. Oncol. 13, 77–78 (2016).
Smyth, E. C. & Cunningham, D. Encouraging results for PD-1 inhibition in gastric cancer. Lancet Oncol. 17, 682–683 (2016).
Rosenberg, J. E. et al. Atezolizumab in patients with locally advanced and metastatic urothelial carcinoma who have progressed following treatment with platinum-based chemotherapy: a single-arm, multicentre, phase 2 trial. The Lancet 387, 1909–1920 (2016).
Naidoo, J. et al. Toxicities of the anti-PD-1 and anti-PD-L1 immune checkpoint antibodies. Ann. Oncol. 26, 2375–2391 (2015).
Mellati, M. et al. Anti-PD-1 and anti-PDL-1 monoclonal antibodies causing type 1 diabetes. Diabetes Care 38, e137–e138 (2015).
Boutros, C. et al. Safety profiles of anti-CTLA-4 and anti-PD-1 antibodies alone and in combination. Nat. Rev. Clin. Oncol. 13, 473–486 (2016).
Larkin, J. et al. Combined nivolumab and ipilimumab or monotherapy in untreated melanoma. N. Engl. J. Med. 373, 23–34 (2015).
Chen, L. & Han, X. Anti-PD-1/PD-L1 therapy of human cancer: past, present, and future. J. Clin. Invest. 125, 3384–3391 (2015).
Weber, J. S., Kahler, K. C. & Hauschild, A. Management of immune-related adverse events and kinetics of response with ipilimumab. J. Clin. Oncol. 30, 2691–2697 (2012).
Woo, S. R., Corrales, L. & Gajewski, T. F. The STING pathway and the T cell-inflamed tumor microenvironment. Trends Immunol. 36, 250–256 (2015).
Hegde, P. S., Karanikas, V. & Evers, S. The where, the when, and the how of immune monitoring for cancer immunotherapies in the era of checkpoint inhibition. Clin. Cancer Res. 22, 1865–1874 (2016).
Spranger, S. et al. Up-regulation of PD-L1, IDO, and Tregs in the melanoma tumor microenvironment is driven by CD8+ T cells. Sci. Transl. Med. 5, 200ra116 (2013).
Gajewski, T. F., Schreiber, H. & Fu, Y. X. Innate and adaptive immune cells in the tumor microenvironment. Nat. Immunol. 14, 1014–1022 (2013).
Fesnak, A. D., June, C. H. & Levine, B. L. Engineered T cells: the promise and challenges of cancer immunotherapy. Nat. Rev. Cancer 16, 566–581 (2016).
Yoo, J. W., Irvine, D. J., Discher, D. E. & Mitragotri, S. Bio-inspired, bioengineered and biomimetic drug delivery carriers. Nat. Rev. Drug Discov. 10, 521–535 (2011).
Tamagawa-Mineoka, R. Important roles of platelets as immune cells in the skin. J. Dermatol. Sci. 77, 93–101 (2015).
Franco, A. T., Corken, A. & Ware, J. Platelets at the interface of thrombosis, inflammation, and cancer. Blood 126, 582–588 (2015).
Hu, C. M. et al. Nanoparticle biointerfacing by platelet membrane cloaking. Nature 526, 118–121 (2015).
Textor, J. in Platelet-Rich Plasma (eds Lana, J. F., Santana, M. H. A., Belangero, W. D., & Luzo, A. C. M. ) 61–94 (Springer, 2014).
Harker, L. A. et al. Effects of megakaryocyte growth and development factor on platelet production, platelet life span, and platelet function in healthy human volunteers. Blood 95, 2514–2522 (2000).
Nurden, A. T., Nurden, P., Sanchez, M., Andia, I. & Anitua, E. Platelets and wound healing. Front. Biosci. 13, 3532–3548 (2008).
Gay, L. J. & Felding-Habermann, B. Contribution of platelets to tumour metastasis. Nat. Rev. Cancer 11, 123–134 (2011).
Nash, G. F., Turner, L. F., Scully, M. F. & Kakkar, A. K. Platelets and cancer. Lancet Oncol. 3, 425–430 (2002).
Hu, Q. et al. Anticancer platelet-mimicking nanovehicles. Adv. Mater. 27, 7043–7050 (2015).
Garraud, O. Editorial: platelets as immune cells in physiology and immunopathology. Front. Immunol. 6, 1–3 (2015).
Morrell, C. N., Aggrey, A. A., Chapman, L. M. & Modjeski, K. L. Emerging roles for platelets as immune and inflammatory cells. Blood 123, 2759–2767 (2014).
Semple, J. W., Italiano, J. E. & Freedman, J. Platelets and the immune continuum. Nat. Rev. Immunol. 11, 264–274 (2011).
Elzey, B. D. et al. Platelet-mediated modulation of adaptive immunity: a communication link between innate and adaptive immune compartments. Immunity 19, 9–19 (2003).
Seifert, L. et al. The necrosome promotes pancreatic oncogenesis via CXCL1 and Mincle-induced immune suppression. Nature 532, 245–249 (2016).
Topalian, S. L., Drake, C. G. & Pardoll, D. M. Targeting the PD-1/B7-H1(PD-L1) pathway to activate anti-tumor immunity. Curr. Opin. Immunol. 24, 207–212 (2012).
Siljander, P. R. M. Platelet-derived microparticles—an updated perspective. Thromb. Res. 127, S30–S33 (2011).
Li, J., Sharkey, C. C., Wun, B., Liesveld, J. L. & King, M. R. Genetic engineering of platelets to neutralize circulating tumor cells. J. Control. Release 228, 38–47 (2016).
Ruggeri, Z. M. & Mendolicchio, G. L. Adhesion mechanisms in platelet function. Circ. Res. 100, 1673–1685 (2007).
Mause, S. F., von Hundelshausen, P., Zernecke, A., Koenen, R. R. & Weber, C. Platelet microparticles—a transcellular delivery system for RANTES promoting monocyte recruitment on endothelium. Arterioscler. Thromb. Vasc. Biol. 25, 1512–1518 (2005).
Tripathi, S. & Guleria, I. Role of PD1/PDL1 pathway, and TH17 and treg cells in maternal tolerance to the fetus. Biomed. J. 38, 25–31 (2015).
Headley, M. B. et al. Visualization of immediate immune responses to pioneer metastatic cells in the lung. Nature 531, 513–517 (2016).
Flaumenhaft, R. Formation and fate of platelet microparticles. Blood Cells Mol. Dis. 36, 182–187 (2006).
Rand, M. L., Wang, H., Bang, K. W., Packham, M. A. & Freedman, J. Rapid clearance of procoagulant platelet-derived microparticles from the circulation of rabbits. J. Thromb. Haemost. 4, 1621–1623 (2006).
Lu, Y., Aimetti, A. A., Langer, R. & Gu, Z. Bioresponsive materials. Nat. Rev. Mater. 1, 16075 (2016).
Cazenave, J.-P. et al. in Platelets and Megakaryocytes: Volume 1: Functional Assays (eds Gibbins, J. M. & Mahaut-Smith, M. P. ) 13–28 (Methods In Molecular Biology Series Vol. 272, Humana, 2004).
Janowska-Wieczorek, A. et al. Platelet-derived microparticles bind to hematopoietic stem/progenitor cells and enhance their engraftment after transplantation. Blood 98, 3143–3149 (2001).
Cheville, N. F. & Stasko, J. Techniques in electron microscopy of animal tissue. Vet. Pathol. 51, 28–41 (2014).
Zimmerman, M., Hu, X. & Liu, K. Experimental metastasis and CTL adoptive transfer immunotherapy mouse model. J. Vis. Exp. 45, 2077 (2010).
Fischer, A. H. et al. Hematoxylin and eosin staining of tissue and cell sections. Cold Spring Harb. Protoc. 5, pdb-prot4986 (2008).
Wang, C. et al. In situ activation of platelets with checkpoint inhibitors for post-surgical cancer immunotherapy. figsharehttp://dx.doi.org/10.6084/m9.figshare.4231766 (2016).
Acknowledgements
This work was supported by grants from the Alfred P. Sloan Foundation (Sloan Research Fellowship), NC TraCS, the National Institutes of Health (Clinical and Translational Science Award (CTSA, NIH grant 1L1TR001111) to Z.G.) and a pilot grant from the University of North Carolina (UNC) Cancer Center. We acknowledge L. Huang at UNC at Chapel Hill for providing the B16F10-Luc-GFP and 4T1-Luc-GFP cell lines.
Author information
Authors and Affiliations
Contributions
C.W. and Z.G. designed the project. C.W., W.S., Y.Y and Q.H. performed the experiments. All authors analysed and interpreted the data. All authors contributed to the writing of the manuscript. All authors discussed the results and implications and edited the manuscript at all stages.
Corresponding author
Ethics declarations
Competing interests
Z.G. and C.W. have a pending patent entitled ‘Platelets for delivery of cancer immunotherapeutics’ (patent number, 10620-039PV1).
Supplementary information
Supplementary Information
Supplementary Figures (PDF 4148 kb)
Rights and permissions
About this article

Cite this article
Wang, C., Sun, W., Ye, Y. et al. In situ activation of platelets with checkpoint inhibitors for post-surgical cancer immunotherapy. Nat Biomed Eng 1, 0011 (2017). https://doi.org/10.1038/s41551-016-0011
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41551-016-0011
This article is cited by
-
In-situ-sprayed therapeutic hydrogel for oxygen-actuated Janus regulation of postsurgical tumor recurrence/metastasis and wound healing
Nature Communications (2024)
-
The dynamic role of platelets in cancer progression and their therapeutic implications
Nature Reviews Cancer (2024)
-
Cell-mediated nanoparticle delivery systems: towards precision nanomedicine
Drug Delivery and Translational Research (2024)
-
Bionic immunoactivator copresenting autophagy promoting and costimulatory molecules for synergistic cancer immunotherapy
Nano Research (2024)
-
Cholesterol removal improves performance of a model biomimetic system to co-deliver a photothermal agent and a STING agonist for cancer immunotherapy
Nature Communications (2023)
