
Abstract
Changes in ocean ventilation have been pivotal in regulating carbon sequestration and release on centennial to millennial timescales. However, paleoceanographic reconstructions documenting changes in deep-ocean ventilation using 14C dating, may bear multidimensional explanations, obfuscating the roles of ocean ventilation played on climate evolution. Here, we show that previously inferred poorly ventilated conditions in the North Atlantic were linked to enhanced pre-aged organic carbon (OC) input during Heinrich Stadial 1 (HS1). The 14C age of sedimentary OC was approximately 13,345 ± 692 years older than the coeval foraminifera in the central North Atlantic during HS1, which is coupled to a ventilation age of 5,169 ± 660 years. Old OC was mainly of terrigenous origin and exported to the North Atlantic by ice-rafting. Remineralization of old terrigenous OC in the ocean may have contributed to, at least in part, the anomalously old ventilation ages reported for the high-latitude North Atlantic during HS1.
Similar content being viewed by others

Introduction
Rapid changes in ocean-circulation dynamics during the last deglaciation may provide fundamental constraints for understanding the past and predicting future variations in atmospheric pCO21,2. Many paleoceanographic reconstructions are consistent with a more poorly ventilated high-latitude deep North Atlantic Ocean during the last ice age and early deglaciation, with ventilation ages typically ranging between ~1000 and ~5000 yr (the benthic-atmosphere radiocarbon age offset)3, significantly exceeding modern values (~400 yr)4. Reduced ventilation-defined here as the degree of air-sea equilibration - in the North Atlantic has commonly been attributed to dynamic changes in the Atlantic Meridional Overturning Circulation (AMOC) and the northward advection of poorly ventilated water masses from the Southern Ocean5,6. When pronounced local ventilation anomalies are taken into consideration, such as ventilation ages exceeding 5000 years in the intermediate-depth eastern equatorial Pacific, the dominant paradigm involving the advection of poorly ventilated water masses becomes, however, potentially inconsistent7,8. Indeed, 14C-depleted (local) carbon sources, such as inputs of 14C-dead hydrothermal carbon to the abyssal ocean7,9, may account for some of the old ventilation ages inferred based on marine authigenic carbonate phases7. As changes in ocean circulation dynamics have been pivotal in modulating abrupt climate variability since the Last Glacial Maximum (LGM)10,11,12, it is essential to understand the mechanisms that may have caused changes in ocean ventilation in the past and in particular across the last glacial termination.
Paleoceanographic evidence gleaned through the testimony of Northwestern Atlantic sediment cores indicates that catastrophic events episodically occurred during the last deglaciation, including outburst floods13, abrupt fluvial runoff14, and ice-rafting events15,16. The most prominent of these perturbations relate to Heinrich Events (HEs), during which large amounts of freshwater and terrigenous material13,14,15,16 surged to the high-latitude North Atlantic and were discharged through icebergs17, which transiently altered the intensity of the AMOC18 and led to abrupt cooling in the northern hemisphere15,16. Modern oceanographic studies indicate that melting glaciers and ice sheets export large volumes of dissolved organic carbon (DOC, 1.04 ± 0.18 Tg·yr−1, 1 T = 1012) and particulate organic carbon (POC, 1.97 Tg·yr−1) to the ocean19, with both fractions being prone to bacterial degradation20,21. Given the widespread influence of HEs in the high-latitude North Atlantic15,22, the impact of the pre-aged terrigenous OC supply and subsequent remineralization may significantly have affected the regional (radio)carbon inventory. Enhanced terrigenous OC supply associated with HEs could thus provide a potential source of 14C-depleted carbon to the ocean and account for, at least in part, the anomalously old ventilation ages reported for the high-latitude North Atlantic during the last deglaciation.
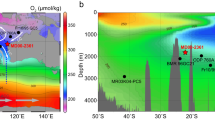
In this study, we measured the 14C ages of sedimentary OC and paired planktic and benthic foraminifera in three North Atlantic sediment cores (Sites U1302, 3560 m; U1308, 3870 m; and U1314, 2820 m; Fig. 1a and Methods) to investigate the impact the remineralization of pre-aged terrigenous OC imposed on North Atlantic ventilation age reconstructions across the last glacial termination. These three sites are located in the western, central, and northern North Atlantic, respectively, enabling us to investigate the spatiotemporal relationships between pre-aged terrigenous OC input associated with ice-rafting events and the evolution of subsurface ocean ventilation. Our data indicate that a large amount of terrigenous OC was exported to the North Atlantic during Heinrich Stadial 1 (HS1) and - to a lesser extent - the Younger Dryas (YD), which was substantially older than co-occurring foraminifera. The pre-aged terrigenous OC input was directly coupled with a decline in subsurface ocean ventilation inferred based on the 14C age difference between coeval benthic-planktic (B-P) foraminifera in the central North Atlantic. Thus, our findings provide a perspective for understanding the multi-facetted mechanisms underlying reduced ocean ventilation in the high-latitude North Atlantic and their implications for the oceanic carbon cycle.

a Map showing the core sites considered in this study (Supplementary Table 1)4,5,11,37,38,39,40,43,51,61. The grey shaded area represents Ruddiman’s IRD belt15,16. The dashed lines indicate the maximum ice sheet extent at the LGM67. bi North Greenland Ice Core Project (NGRIP) δ18O serves as a reference chronology68, the grey line represents the raw data and the black line reflects the 10-point running mean; ii sedimentary terrigenous Organic Carbon (OC) content (indicated by the δ13Corg data) at sites U1302 (magenta), U1308 (green), and U1314 (blue); iii 14C ages targeting the different sedimentary fractions, namely Total Organic Carbon (TOC, dots), planktic foraminifera (triangles), and benthic foraminifera (diamonds) at sites U1302 (magenta), U1308 (green), and U1314 (blue). The percentage of terrigenous OC is derived based on δ13Corg, and the old OC is indicated by the older 14C age of TOC when compared to the co-deposited foraminifera. BA Bølling–Allerød, HS1 Heinrich Stadial 1, LGM Last Glacial Maximum, YD Younger Dryas. The earth map is drawn by Ocean Data View (Schlitzer, Reiner, Ocean Data View, odv.awi.de, 2023)69.
Results and discussion
Enhanced old terrigenous OC input to the North Atlantic during HS1
The 14C ages of sedimentary OC vary between 18,450 ± 150 and 30,200 ± 310 yr during HS1. These 14C ages are significantly older than those of co-occurring foraminifera, which typically range between 13,000 ± 50 and 18,950 ± 500 yr (Fig. 1b). On average, sedimentary OC is thus 8,564 ± 305 yr older than planktic foraminifera and 6,650 ± 478 yr older than benthic foraminifera during HS1 (Supplementary Dataset S2). Beyond HS1, the age difference between OC and biogenic carbonate phases is much more subdued, with sedimentary OC on average only 2,782 ± 174 yr older than coeval foraminifera (Fig. 2b, Supplementary Dataset S2). Furthermore, the δ13Corg and δ15Norg values at sites U1302 and U1308 are significantly lower during HS1, in line with the maximum C/N ratio (Supplementary Figs. 6 and 7), consistent with increased sedimentary burial of terrigenous OC in the western and central North Atlantic23. Microscopic observations suggest that the sediment layer is enriched with ice-rafted debris (IRD) (Supplementary Fig. 9), indicating that OC is intimately related to ice-rafting events characteristic of HS15,16,24. Rocks and sediments underlying the Laurentide ice sheet have been suggested to provide the main sources of IRDs in the western and central North Atlantic15,16,17 (Fig. 1a). Site U1314 is located south of Iceland, implying that the northern North Atlantic may also have received terrestrial material from Iceland, Greenland, and/or Scandinavia15. Furthermore, our results indicate that the sedimentary OC is ~4000 yr older than the coeval foraminifera at Site 1314, suggesting that pre-aged terrigenous OC may also have been transported to the northern North Atlantic (Fig. 2b). While pilot studies reported that hydrodynamic processes could lead to OC aging about thousands years in the relatively-shallow oceans25,26,27 and bioturbation may also affect the OC 14C ages28, it is difficult to interpret the such large 14C age difference between OC and coeval planktonic foraminifera (~4240–15,440 yr) during HS1 in northern North Atlantic (More information see Supplementary Materials). To sum up, it is more reasonable that enhanced old terrigenous OC input is mainly attributed to the observed large 14C offset need the contribution of pre-age OC remineralization.

The data for sites U1302, U1308, and U1314 were generated in the context of this study; the data for sites 984, NEAP and BOFS were generated previously38,61. a Binary diagrams for δ13Corg and 14C offsets between Total Organic Carbon (TOC) and planktic foraminifera were used to identify the pre-aged, terrigenous OC fraction (with associated 1σ-uncertainties). The samples spanning HS1, HS2, and YD are indicated in red shading. b Simulated 14C offset between TOC and benthic/planktic foraminifer versus sample age. Significant 14C offsets between TOC and foraminifera were observed during HS1, HS2, and the YD (with associated 1σ-uncertainties), which indicate that pre-aged terrigenous OC input may have been derived from HEs. The few anomalous 14C values of benthic foraminifera are indicated by question marks. HS1 Heinrich Stadial 1, HS2 Heinrich Stadial 2, YD Younger Dryas.
Remineralization of old terrigenous OC and its influence on ventilation ages
Our data reveal that the pre-aged terrigenous OC deposited during HS1, both in the central (U1308 Site) and northern (U1314 Site) North Atlantic, correspond to reduced ocean ventilation reconstructed based on B-P 14C ages with age differences ranging between 1950 ± 135 and 4190 ± 545 yr (Fig. 1b, Supplementary Figs. 6 and 8). As pre-aged terrigenous OC degrades, it may substantially alter the (radio)carbon inventory of ambient seawater29, thereby influencing the 14C ages of benthic foraminifera. Stable carbon-isotope evidence suggests that the deep North Atlantic was characterised by enhanced sequestration of remineralized carbon during the last deglaciation29,30, consistent with the hypothesis that pre-aged terrigenous OC degradation may have occurred in the ocean interior.
The spatial heterogeneity in the B-P 14C ages amongst the three sites highlights regional sedimentological characteristics (Fig. 1). In particular, the B-P 14C age differences at Site U1308 indicate anomalously reduced ventilation in the central North Atlantic (4190 ± 545 yr, HS1). This may be related to the location of Site U1308, which lies within the IRD belt (Fig. 1), where the terrigenous OC input flux associated with IRDs may have been the highest15,16. While no significant increase in TOC burial was observed at Site U1308 during the deglaciation, enhanced supply of terrigenous OM has been reported previously based on biomarkers and reconstructions of past changes in sediment accumulation31,32,33, suggesting that the remineralization of old OC may be intensive, impacting on ventilation age reconstructions at site U1308. Site U1314 is located in the northern reaches of the IRD belt, which was likely less affected by old terrigenous carbon input. This may have contributed to the comparatively lower ventilation ages at site U1314 when compared to site U1308. We did not observe reduced ventilation as inferred by 14C age reconstructions during HS1 at Site U1302 (Fig. 1 and Supplementary Fig. 6), possibly owing to the high sedimentation rate (ranging between 16.5–17.3 cm/ka) and spatial differences of IRD sources34. The high sediment accumulation at U1302 may have prevented extensive remineralization of old OC due to its rapid burial and lower bacterial-respiration rates (i.e. reduced oxygen exposure time)35,36 (Supplementary Fig. 6). Therefore, we suggest that the differential remineralization of old terrigenous OC may have contributed to the spatial heterogeneity in the ventilation ages reported for HS1 in the North Atlantic.
Spatial Atlantic ventilation compilations
To further document the potentially obfuscating influence of old terrigenous OC remineralization on ventilation-age reconstructions, we compile the ventilation ages available for the entire North Atlantic basin (Fig. 3). Taking changes in surface reservoir ages into consideration, we hereafter report ventilation ages as the 14C age difference between benthic foraminifera and the contemporaneous atmosphere (B-atm). Relatively well-ventilated intermediate-depth (<2000 yr) waters prevailed from low- to mid-latitudes5,37 in the North Atlantic basin across the last deglaciation (Fig. 3c, e). In contrast, the high-latitude North Atlantic was characterised by generally more poorly ventilated conditions (with B-atm ~5000 yr) at both abyssal (Fig. 3f) and intermediate38 (Fig. 3g) depths during HS1. This spatial pattern is difficult to reconcile with the prevailing notion that old (poorly ventilated) southern-sourced water (SSW) bathed large swaths of the deep North Atlantic at the end of the last ice age5,39,40,41. The ventilation ages of the SSW endmember typically range between ~2000 and 4000 yr5,37 during HS1, somewhat younger than the old ventilation ages of the high-latitude North Atlantic (~5000 14C yr). Emerging evidence indicates that the ventilation of the South Atlantic, and more generally, the Southern Ocean, began to recover at the onset of HS15,37,42. Would SSW have substantially intruded the deep North Atlantic during HS1, it would have resulted in ventilation ages (B-P 14C ages) ranging between ~2000 and 4000 yr, which is arguably incompatible with a sluggishly ventilated high-latitude North Atlantic at that time.

a Greenland Ice Core from the NGRIP δ18O68. b South Atlantic B-atm ages at water depth of 3.8 km5,37, where the shaded area indicates the maximum/minimum range of B-atm offsets. c Coral-derived B-atm ages in the Equatorial Atlantic11. d B-atm ages at the Iberian Margin (data of Site SHAK06-5K (2.6 km, pink) and Site MD99-2334 (3.1 km, sky blue)51. The data for Site MD99-2334 (3.1 km, brown) were also reported previously5,40. The shaded area indicates the maximum/minimum range of B-atm offsets at site MD99-2334. e B-atm ages (coral-derived data, orange diamonds; benthic foram-derived data, light green diamonds) from the northwest Atlantic seamounts at water depths ranging 1.1–4.6 km39. f B-atm ages of the high-latitude North Atlantic at 2.8–3.9 km (three sites in this study). The anomalous 14C values of benthic foraminifera are indicated by question marks. In term of the overall Atlantic ventilation compilation, we assembled our data from three sites to represent the situation of the high-latitude deep North Atlantic. g B-atm ages of the south Iceland Rise at depth of 1.2–2.3 km3. h, B-atm ages of the Arctic Mediterranean at a water depth of 2.7 km43. i 231Pa/230Th and εNd proxies showed variations of the AMOC and NADW, respectively. The orange lines indicate 231Pa/230Th data17,44, and the grey lines indicate εNd data45,46 measured at Bermuda Rise. All of the above have an associated 1σ-uncertainty.
The poorly ventilated Arctic-sourced water (ventilation ages up to 10,400 yr) may have contributed to reduce ventilation in the high-latitude North Atlantic43 during the last ice age; however, whether it can also influence the deep North Atlantic remains questionable. To some extent, sedimentary 231Pa/230Th and εNd evidence is consistent with a weaker AMOC state and reduced North Atlantic Deep Water (NADW) formation during HS117,44,45,46 (Fig. 3i), which may suggest that the contribution of overflow water from the Arctic Mediterranean to the North Atlantic remained limited. Furthermore, millennial-scale variations in ocean ventilation in the (sub)polar North Atlantic (Fig. 3f–g) and Arctic Mediterranean (Fig. 3h) coincided with those reported in the mid- (Fig. 3d,e) and low-latitudes North- (Fig. 3c) and South Atlantic (Fig. 3b), albeit to a much smaller magnitude. Such abrupt variations in ventilation ages challenge the mechanisms proposed to account for water mass advection from the south.
Assuming that the input of terrigenous OC to the high-latitude North Atlantic via ice-rafting amounted ~50 Tg·yr−1 during HS1 – 50 times the OC-input flux by the modern Greenland Ice Sheet (~1 Tg·yr−1)18,20 – sustained for ~1500 yr47, we estimate that ~75 Pg (1 Pg = 1015 g) of terrigenous OC may have been exported to the North Atlantic during HS1. We estimate that at least ~18 Pg terrigenous OC would have been remineralized in the North Atlantic during HS1. Thus, the remineralized OC may have significantly impacted the 14C ages of benthic foraminifera (14CBenthic), which are commonly used for ventilation-age reconstructions. Based on our semi-quantitative framework (Method and Fig. 4), assuming that ~10.7–18.1 Pg pre-aged terrigenous OC was remineralized and efficiently assimilated by benthic foraminifera during HS1, the overestimation of ventilation age may amount up to thousand years during HS1. Furthermore, marine-sourced, labile OC may be pre-aged in the water column before deposition48. The aged marine OC degradation may further accelerate the remineralization of pre-aged terrigenous refractory OC49,50. Considering the duration of the events, the maturation and degradation of marine OC and its potential impact on ventilation age reconstructions may be a target for research in the future. Hence, researchers should not ignore the influence of pre-aged remineralized OC on reconstructed ventilation ages5,37,40,51.

14C ages of surface water and bottom water serve as a basis for developing the theoretical ventilation-age model described below (Methods; theoretical ventilation age). Pre-aged Organic Carbon (OC) contribution to benthic foraminifera (\({{{{{{\rm{f}}}}}}}_{{14}_{{{{{{\rm{C}}}}}}-{{{{{\rm{dead}}}}}}-{{{{{\rm{OC}}}}}}}}\)) tended to negatively correlate with the ventilation agemodel in the context of an old ventilation age (4190 ± 545 yr of B-P, with associated 1σ-uncertainties) at site U1308 during HS1. The results of the ventilation agemodel were dramatically overprinted by pre-aged OC remineralization in the North Atlantic during HS1. The positive seawater-ventilation age trend can be theoretically tuned via remineralization of pre-aged OC (insert square). The ordinary ventilation age could be hundreds, or even a thousand years (extreme value up to 2000 yr)3 may be achieved based on pre-aged OC inputs ranging between 14.2 ± 6.3% and 24.1 ± 5.2% \({{{{{{\rm{f}}}}}}}_{{14}_{{{{{{\rm{C}}}}}}-{{{{{\rm{dead}}}}}}-{{{{{\rm{OC}}}}}}}}\).
In addition, the terrigenous OC supplied by ice-rafting to the high-latitude North Atlantic may have provided a hitherto unaccounted for source of carbon to the atmosphere during the last deglaciation. Were the respired carbon derived from the remineralization terrigenous OC released to the atmosphere, then it may have contributed, at least in part, to the rapid rise in atmospheric CO2 concentrations1. HEs or Heinrich-like events have been proposed to have occurred during each glacial termination, since at least ~640 ka BP16. It is therefore possible that the terrigenous OC input associated with HEs or Heinrich (-like) events and its subsequent remineralization and release may have contributed to the rapid rises in atmospheric CO2 that characterised previous glacial terminations52. Further investigations are needed to better quantify the impact of catastrophic outflow events on ocean ventilation and circulation during deglaciations and their implication for the global carbon cycle.
Methods
Sites and samples
Integrated Ocean Drilling Program (IODP) Site U1314 is located on the southwest flank of the Reykjanes Ridge (Iceland) on the southern Gardar Drift (2820 m, 56°21.89′N, 27°53.28′W) (Fig. 1). The Gardar Drift is largely influenced by the lower part of the AMOC, the Iceland‐Scotland Overflow Water (ISOW), or the Northeast Atlantic Deep Water (NEADW) and seafloor topography53. Site U1308 (3870 m, 49°52.67′N; 24°14.28′W) is located in the central North Atlantic and Ruddiman’s IRD belt, making it a key site for investigating ocean–atmosphere–ice sheet interactions, which have a profound impact on global climate change. Site U1302 (3560 m, 50°10.01′N; 45°38.32′W) is located on the crest of a small ridge, east of a fault scarp marking the eastern side of Orphan Knoll. Site U1302 is bathed by Labrador Sea Water which may transport terrestrial materials from the North American continent54,55.
At these sites, IRDs were commonly deposited during the last deglaciation. We sampled the upper 3.12 mcd composite core of U1314B/C at 10 cm resolution, the upper 1.40 mcd of U1308B at 2 cm resolution, and the upper 3.76 mcd of U1302D/E at 4 cm resolution (which spans the last ~30 ka BP, thousand years before present), in order to investigate to which degree old terrigenous OC input may have affected 14C-based ocean ventilation reconstructions in the North Atlantic.
Age model
The age models developed in this study were primarily based on 14C dating of planktonic foraminifera in the three IODP cores. The age model used for U1314 was based on seven planktic foraminifera (G. bulloides) 14C dates and Bacon modelling56. The reservoir ages and standard errors at Site 1314 were obtained from the literature57. The stratigraphy for site U1308 used here was also published previously47, with 24 planktic foraminifera (G. bulloides) used for 14C dating at sites U1308A47 and 60958. We recalculated calendar ages based on the latest Marine20 curve and Bacon modelling59. We used magnetic susceptibility to graphically align U1308A, U1308B, and U1308C and propose a revised metre composite depth (rmcd) model. The age model for U1302 was based on four planktonic foraminifera (N. pachyderma) 14C dates. Owing to the lack of constraints on the reservoir age during the last deglaciation in the vicinity of site U1302, modern ΔR values were used to determine calendar ages using the Bacon model. See the Supplementary Information for additional details (Supplementary Figs. 1 and 4).
TOC, TN, and δ13Corg, δ15Norg analysis
The samples were freeze-dried, weighed (0.5 g), homogenised, and acidified using 1 M HCl to remove carbonate until no bubbles were present. The samples were then rinsed repeatedly with Milli-Q water until the pH reached neutrality, after which the samples were freeze-dried again. All glassware used was pre-combusted at 450 °C for 5 h prior to use. Carbon and nitrogen content and isotope analyses were performed with an EA-IsoLink elemental analyser coupled to a MAT253 plus isotope ratio mass spectrometer via a ConFlo IV universal interface (all from Thermo Fisher Scientific, Bremen, Germany) at the Key Lab of Submarine Geosciences and Prospecting Techniques (Ministry of Education, Ocean University of China). The δ13Corg data are reported relative to the Vienna Pee Dee Belemnite (V-PDB) standard with an external analytical precision of ±0.1 ‰, and the δ15N data are reported relative to air with an external analytical precision of ±0.5 ‰. The external analytical precisions of the TOC and TN data were ±5 ‰ and ±10 ‰, respectively.
14C analysis
In this study, we measured the 14C contents of TOC and foraminifera at the National Ocean Sciences Accelerator Mass Spectrometry Facility at Woods Hole Oceanographic Institution (WHOI). Acid pre-treatment was conducted at the WHOI. Planktonic foraminifera (Globigerina bulloides and Neogloboquadrina pachyderma (sin.)) were subjected to five interations of ultrasonic treatment in Milli-Q water to remove excess clay minerals60, as shown in Supplementary Dataset S1. The benthic foraminifera for conducting 14C analysis included Cibicidoides wellerstorfi, Uvigerina spp. and Melonis spp.
Source apportionment of OC
The following two-end-member model (terrigenous and marine OC) for δ13C was used to identify the OC source (Supplementary Dataset S3–S5):
where δ13CTOC is the δ13C of bulk sediments and fterr and fmar represent the relative proportions of terrigenous OC and marine OC, respectively. The δ13Cterr value was assigned to an average value of −27‰, reflecting the −26 to −29‰ δ13C values of TOC and the −26 to −30‰ δ13C values for Phanerozoic OC-rich rocks underlying these soils61,62. The δ13Cmar endmember value amounts to −20‰, representative of the values reported previously61,63.
Furthermore, the bulk sedimentary δ15N and C/N values were used to constrain terrigenous OC input. According to previous studies, C/N ratios >1064 and δ15N values of <1‰65 are diagnostic for a terrigenous provenance for the OC.
Ventilation by B-P foraminifera and the B-atm
In this study, two methods were used to quantify past changes in ocean ventilation40, namely B-P foraminifera age differences and the 14C offset between benthic foraminifera and the contemporary atmosphere (B-atm). To investigate the potential contrasts in ventilation age reconstructions among the three sites in this study, we used the B-P approach to avoid the uncertainties related to constraining calendar ages. When comparing ventilation ages across the entire Atlantic, the B-atm approach was used to unify the available datasets and overcome the drawback pertaining to the lack of planktonic data, which was reported as a limitation in previous studies3,11,39,43. In this study, when considering the western North Atlantic39, we used atmospheric 14C ages derived from IntCal2059 (see the data in Supplementary Dataset S1 and S2 for further details). We refer to the original data from the literature to obtain the ventilation ages mentioned in the main text. We believe that the influence of using different IntCal datasets on ventilation age reconstructions remained within the normal error range (generally <500 14C yr) and did not affect our argumentation.
Theoretical ventilation age
First, two end-member models (OC and bottom water) for Δ14C were applied to quantify the 14C contribution from OC to the benthic foraminifera
where Δ14CBenthic, Δ14COC, and Δ14CBW represent Δ14C values derived from benthic foraminifera, OC, and bottom water, respectively; fOC and fBW represent the relative Δ14C proportions of OC and the bottom water to benthic foraminifera. Based on the 14C age offsets between the OC and benthic/planktic foraminifers shown in Fig. 2B, we selected values for Δ14CBenthic (−905.8%) and Δ14COC (−976.9%) to analyse the relationship between fOC and Δ14CBW. These values for Δ14CBenthic and Δ14COC were defined at Site U1308 where the ventilation ages were the oldest (4,190 ± 545 yr, B-P 14C age).
Second, we substituted Eqs. (3) and (4) to obtain the relationship between fOC and Δ14CBW, using Eq. (5).
The theoretical ventilation age (B-P 14C age) was considered as representing the offset in 14C age between bottom and surface waters, and AgeSW was considered a fixed value for the 14C age of planktonic foraminifera during HS1.
Where, AgeBW and AgeSW represented the 14C age of the bottom water and surface water, repsectively. AgeBW would be expressed as follows:
FmBW was the Fm value of bottom water.
By substituting Eqs. (5), (7), and (8) into Eq. (6), we obtained Eq. (9) that related the ventilation agemodel and fOC values
Here, AgeplanktonicHS1 was a fixed value for the 14C age of planktonic foraminifera during HS1.
Finally, we resolved the relationship between fOC and \({{{{{{\rm{f}}}}}}}_{{14}_{{{{{{\rm{C}}}}}}-{{{{{\rm{dead}}}}}}-{{{{{\rm{OC}}}}}}}}\) during HS1 to determine the function relating ventilation the Agemodel and \({{{{{{\rm{f}}}}}}}_{{14}_{{{{{{\rm{C}}}}}}-{{{{{\rm{dead}}}}}}-{{{{{\rm{OC}}}}}}}}\) values (14C-dead contribution of OC to benthic foraminifera; Fig. 4).
To quantify the amount of pre-aged OC exported into the North Atlantic that caused the largest ventilation age during HS1 at U1308, we adopted Eqs. (10) and (11):
Δ14COC, Δ14Cdead carbon and Δ14CAgeHS1 represented fixed Δ14C dates for OC, 14C dead carbon, and the 14C age model, respectively; fdead carbon represented the percentage of dead 14C in OC.
We derived that OC was composed of 82.9% dead carbon during HS1.
Furthermore, we estimated the pre-aged OC flux based on Eq. (12):
where the OC flux was estimated as described above in the main text as 75 Pg in the North Atlantic during HS1; we estimated that the pre-aged OC flux ranged from 10.7 to 18.1 Pg.
Data availability
The data generated in this study are provided in the Supplementary Information and Figshare data repository (https://doi.org/10.6084/m9.figshare.22634719)66.
References
Marcott, S. A. et al. Centennial-scale changes in the global carbon cycle during the last deglaciation. Nature 514, 616–619 (2014).
Shakun, J. D. et al. Global warming preceded by increasing carbon dioxide concentrations during the last deglaciation. Nature 484, 49–54 (2012).
Thornalley, D. J. R., Barker, S., Broecker, W. S., Elderfield, H. & McCave, I. N. The deglacial evolution of North Atlantic deep convection. Science 331, 202–205 (2011).
Adkins, J. F., Cheng, H., Boyle, E. A., Druffel, E. R. M. & Edwards, R. L. Deep-sea coral evidence for rapid change in ventilation of the deep North Atlantic 15,400 years ago. Science 280, 725–728 (1998).
Skinner, L. C., Waelbroeck, C., Scrivner, A. E. & Fallon, S. J. Radiocarbon evidence for alternating northern and southern sources of ventilation of the deep Atlantic carbon pool during the last deglaciation. Proc. Natl Acad. Sci. USA 111, 5480–5484 (2014).
Martrat, B. et al. Four climate cycles of recurring deep and surface water destabilizations on the Iberian Margin. Science 317, 502–507 (2007).
Stott, L. D. & Harazin, K. M. & Quintana Krupinski, N. B. Hydrothermal carbon release to the ocean and atmosphere from the eastern equatorial Pacific during the last glacial termination. Environ. Res. Lett. 14, 025007 (2019).
Stott, L., Southon, J., Timmermann, A. & Koutavas, A. Radiocarbon age anomaly at intermediate water depth in the Pacific Ocean during the last deglaciation. Paleoceanography 24, PA2223 (2009).
Zhao, N. & Keigwin, L. D. An atmospheric chronology for the glacial-deglacial Eastern Equatorial Pacific. Nat. Commun. 9, 3077 (2018).
Skinner, L. C. et al. Radiocarbon constraints on the glacial ocean circulation and its impact on atmospheric CO2. Nat. Commun. 8, 16010 (2017).
Chen, T. et al. Persistently well-ventilated intermediate-depth ocean through the last deglaciation. Nat. Geosci. 13, 733–738 (2020).
Li, T. et al. Rapid shifts in circulation and biogeochemistry of the Southern Ocean during deglacial carbon cycle events. Sci. Adv. 6, eabb3807 (2020).
Nicholl, J. A. L. et al. A Laurentide outburst flooding event during the last interglacial period. Nat. Geosci. 5, 901–904 (2012).
Ménot, G. et al. Early reactivation of European rivers during the last deglaciation. Science 313, 1623–1625 (2006).
Hemming, S. R. Heinrich events: massive late Pleistocene detritus layers of the North Atlantic and their global climate imprint. Rev. Geophys. 42, 1–43 (2004).
Naafs, B. D. A., Hefter, J. & Stein, R. Millennial-scale ice rafting events and Hudson Strait Heinrich(-like) Events during the late Pliocene and Pleistocene: a review. Qua. Sci. Rev. 80, 1–28 (2013).
Hefter, J., Naafs, B. D. A. & Zhang, S. Tracing the source of ancient reworked organic matter delivered to the North Atlantic Ocean during Heinrich Events. Geochim. Cosmochim. Acta 205, 211–225 (2017).
McManus, J. F., Francois, R., Gherardi, J.-M., Keigwin, L. D. & Brown-Leger, S. Collapse and rapid resumption of Atlantic meridional circulation linked to deglacial climate changes. Nature 428, 834–837 (2004).
Hood, E., Battin, T. J., Fellman, J., O’Neel, S. & Spencer, R. G. M. Storage and release of organic carbon from glaciers and ice sheets. Nat. Geosci. 8, 91–96 (2015).
Hood, E. et al. Glaciers as a source of ancient and labile organic matter to the marine environment. Nature 462, 1044–1047 (2009).
Lawson, E. C. et al. Greenland Ice Sheet exports labile organic carbon to the Arctic oceans. Biogeosciences 11, 4015–4028 (2014).
Bard, E., Rostek, F., Turon, J.-L. & Gendreau, S. Hydrological impact of Heinrich events in the subtropical northeast Atlantic. Science 289, 1321–1324 (2000).
Huon, S., Grousset, F. E., Burdloff, D., Bardoux, G. & Mariotti, A. Sources of fine-sized organic matter in North Atlantic Heinrich Layers: δ13C and δ15N tracers. Geochim. Cosmochim. Acta 66, 223–239 (2002).
Wary, M., Kornilova, O., Russell, M. & Rosell‐Melé, A. Biomarker Fingerprint of Debris Flow Deposits as a Paleoproxy for IRD Sources in the Last Glacial North Atlantic. Paleoceanogr. Paleoclimatol. 35, e2020PA003850 (2020).
Bao, R. et al. Organic carbon aging during across-shelf transport. Geophys. Res. Lett. 45, 8425–8434 (2018).
Magill, C. R. et al. Transient hydrodynamic effects influence organic carbon signatures in marine sediments. Nat. Commun. 9, 4690 (2018).
Ausín, B. et al. Controls on the abundance, provenance and age of organic carbon buried in continental margin sediments. Earth Planet. Sci. Lett. 558, 116759 (2021).
Kim, S. et al. Bioturbation supplying young carbon into West Antarctic continental margin sediment. Palaeogeogr. Palaeoclimatol. Palaeoecol. 602, 111161 (2022).
Oppo, D. W., Curry, W. B. & McManus, J. F. What do benthic δ13C and δ18O data tell us about Atlantic circulation during Heinrich Stadial 1? Paleoceanography 30, 353–368 (2015).
Gu, S. et al. Remineralization dominating the δ13C decrease in the mid-depth Atlantic during the last deglaciation. Earth Planet. Sci. Lett. 571, 117106 (2021).
Madureira, L. A. S. et al. Late Quaternary high-resolution biomarker and other sedimentary climate proxies in a Northeast Atlantic Core. Paleoceanography 12, 255–269 (1997).
Manighetti, B. & McCave, I. N. Depositional fluxes, palaeoproductivity, and ice rafting in the NE Atlantic over the past 30 ka. Paleoceanography 10, 579–592 (1995).
Rosell-Melé, A., Maslin, M. A., Maxwell, J. R. & Schaeffer, P. Biomarker evidence for “Heinrich” events. Geochim. Cosmochim. Acta 61, 1671–1678 (1997).
Barker, S. et al. Icebergs not the trigger for North Atlantic cold events. Nature 520, 333–336 (2015).
Hartnett, H. E., Keil, R. G., Hedges, J. I. & Devol, A. H. Influence of oxygen exposure time on organic carbon preservation in continental margin sediments. Nature 391, 572–575 (1998).
Jessen, G. L. et al. Hypoxia causes preservation of labile organic matter and changes seafloor microbial community composition (Black Sea). Sci. Adv. 3, e1601897 (2017).
Skinner, L. C., Fallon, S., Waelbroeck, C., Michel, E. & Barker, S. Ventilation of the deep Southern Ocean and deglacial CO2 rise. Science 328, 1147–1151 (2010).
Mollenhauer, G., McManus, J. F., Wagner, T., McCave, I. N. & Eglinton, T. I. Radiocarbon and 230Th data reveal rapid redistribution and temporal changes in sediment focussing at a North Atlantic drift. Earth Planet. Sci. Lett. 301, 373–381 (2011).
Robinson, L. F. et al. Radiocarbon variability in the western North Atlantic during the last deglaciation. Science 310, 1469–1473 (2005).
Skinner, L. C. & Shackleton, N. J. Rapid transient changes in northeast Atlantic deep water ventilation age across Termination I. Paleoceanography 19, PA2005 (2004).
Süfke, F. et al. Constraints on the northwestern Atlantic deep water circulation from 231Pa/230Th during the last 30,000 years. Paleoceanogr. Paleocl. 34, 1945–1958 (2019).
Rose, K. A. et al. Upper-ocean-to-atmosphere radiocarbon offsets imply fast deglacial carbon dioxide release. Nature 466, 1093–1097 (2010).
Thornalley, D. J. R. et al. A warm and poorly ventilated deep Arctic Mediterranean during the last glacial period. Science 349, 706–710 (2015).
Lippold, J. et al. Does sedimentary 231Pa/230Th from the Bermuda Rise monitor past Atlantic Meridional Overturning Circulation? Geophys. Res. Lett. 36, L12601 (2009).
Roberts, N. L., Piotrowski, A. M., McManus, J. F. & Keigwin, L. D. Synchronous deglacial overturning and water mass source changes. Science 327, 75–78 (2010).
Gutjahr, M. & Lippold, J. Early arrival of Southern Source Water in the deep North Atlantic prior to Heinrich event 2. Paleoceanography 26, PA2101 (2011).
Hodell, D. A. et al. Anatomy of Heinrich Layer 1 and its role in the last deglaciation: Heinrich event 1. Paleoceanography 32, 284–303 (2017).
Mollenhauer, G. et al. Aging of marine organic matter during cross-shelf lateral transport in the Benguela upwelling system revealed by compound-specific radiocarbon dating. Geochem. Geophys. Geosyst. 8, Q09004 (2007).
Guenet, B., Danger, M., Abbadie, L. & Lacroix, G. Priming effect: bridging the gap between terrestrial and aquatic ecology. Ecology 91, 2850–2861 (2010).
Trevathan-Tackett, S. M., Thomson, A. C. G., Ralph, P. J. & Macreadie, P. I. Fresh carbon inputs to seagrass sediments induce variable microbial priming responses. Sci. Total Environ. 621, 663–669 (2018).
Ausín, B., Sarnthein, M. & Haghipour, N. Glacial-to-deglacial reservoir and ventilation ages on the southwest Iberian continental margin. Qua. Sci. Rev. 255, 106818 (2021).
Lüthi, D. et al. High-resolution carbon dioxide concentration record 650,000–800,000 years before present. Nature 453, 379–382 (2008).
Grützner, J. & Higgins, S. M. Threshold behavior of millennial scale variability in deep water hydrography inferred from a 1.1 Ma long record of sediment provenance at the southern Gardar Drift. Paleoceanography 25, PA4204 (2010).
Channell, J. E. T. et al. A 750-kyr detrital-layer stratigraphy for the North Atlantic (IODP Sites U1302–U1303, Orphan Knoll, Labrador Sea). Earth Planet. Sci. Lett. 317–318, 218–230 (2012).
Channell, J. E. T. & Hodell, D. A. Magnetic signatures of Heinrich-like detrital layers in the Quaternary of the North Atlantic. Earth Planet. Sci. Lett. 369–370, 260–270 (2013).
Blaauw, M. & Christen, J. A. Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 6, 457–474 (2011).
Stern, J. V. & Lisiecki, L. E. North Atlantic circulation and reservoir age changes over the past 41,000 years. Geophys. Res. Lett. 40, 3693–3697 (2013).
Bond, G. et al. Evidence for massive discharges of icebergs into the North Atlantic Ocean during the last glacial period. Nature 360, 245–249 (1992).
Reimer, P. J. et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 cal kBP). Radiocarbon 62, 725–757 (2020).
Barker, S., Greaves, M. & Elderfield, H. A study of cleaning procedures used for foraminiferal Mg/Ca paleothermometry: Mg/Ca Paleothermometry. Geochem. Geophys. Geosyst. 4, 8407 (2003).
Schouten, S., Ossebaar, J., Brummer, G. J. & Elderfield, H. & Sinninghe Damsté, J. S. Transport of terrestrial organic matter to the deep North Atlantic Ocean by ice rafting. Org. Geochem. 38, 1161–1168 (2007).
Verwega, M. T. et al. Description of a global marine particulate organic carbon-13 isotope data set. Earth Syst. Sci. Data 13, 4861–4880 (2021).
Sackett, W. M. Stable carbon isotope studies on organic matter in the marine environment. Handb. Environ. Isot. Geochem. 3, 139–169 (1989).
Meyers, P. A. Organic geochemical proxies of paleoceanographic, paleolimnologic, and paleoclimatic processes. Org. Geochem. 27, 213–250 (1997).
Riethdorf, J.-R. et al. Surface nitrate utilization in the Bering sea since 180 kA BP: Insight from sedimentary nitrogen isotopes. Deep Sea Res. Part II Top. Stud. Oceanogr. 125–126, 163–176 (2016).
Liu, J. et al. Pre-aged terrigenous organic carbon biases ocean ventilation-age reconstructions in the North Atlantic. Dataset. Figshare. https://doi.org/10.6084/m9.figshare.22634719 (2023).
Andrews, J. T. & Voelker, A. H. L. “Heinrich events” (& sediments): a history of terminology and recommendations for future usage. Quat. Sci. Rev. 187, 31–40 (2018).
Svensson, A. et al. A 60 000 year Greenland stratigraphic ice core chronology. Clim. Past 4, 47–57 (2008).
Schlitzer, R. Ocean Data View https://odv.awi.de (2023).
Acknowledgements
This study was supported by the National Natural Science Foundation of China (grant numbers 92058207, 42076037, and 42242601, B.R.) and Fundamental Research Funds for the Central Universities (grant number 202042010, B.R.). This study was also funded by the Taishan Young Scholars (grant number tsqn202103030, B.R.) and the Shandong Natural Science Foundation (grant number ZR2021JQ12, B.R.). We appreciate the IODP community for sampling assistance on cruises and in laboratories.
Author information
Authors and Affiliations
Contributions
R.B. and J.L. designed the research; J.L. and N.W. performed the laboratory analysis; R.B., J.L., and Y.W. analysed the data and wrote the paper. R.B., J.L., Y.W., S.J., X.G., and N.F. contributed to the interpretation of results and commented on the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Blanca Ausin and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Liu, J., Wang, Y., Jaccard, S.L. et al. Pre-aged terrigenous organic carbon biases ocean ventilation-age reconstructions in the North Atlantic. Nat Commun 14, 3788 (2023). https://doi.org/10.1038/s41467-023-39490-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-023-39490-6
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
