
Abstract
Resident memory CD8 T (TRM) cells in the lung parenchyma (LP) and airways provide heterologous protection against influenza virus challenge. However, scant knowledge exists regarding factors necessary to establish and maintain lung CD8 TRM. Here we demonstrate that, in contrast to mechanisms described for other tissues, airway, and LP CD8 TRM establishment requires cognate antigen recognition in the lung. Systemic effector CD8 T cells could be transiently pulled into the lung in response to localized inflammation, however these effector cells failed to establish tissue residency unless antigen was present in the pulmonary environment. The interaction of effector CD8 T cells with cognate antigen in the lung resulted in increased and prolonged expression of the tissue-retention markers CD69 and CD103, and increased expression of the adhesion molecule VLA-1. The inability of localized inflammation alone to establish lung TRM resulted in decreased viral clearance and increased mortality following heterosubtypic influenza challenge, despite equal numbers of circulating memory CD8 T cells. These findings demonstrate that pulmonary antigen encounter is required for the establishment of lung CD8 TRM and may inform future vaccine strategies to generate robust cellular immunity against respiratory pathogens.
Similar content being viewed by others
Introduction
Viral clearance following acute respiratory virus infection results in the establishment of three CD8 T-cell memory reservoirs: central memory T (TCM) cells, largely found in secondary lymphoid organs; effector memory T (TEM) cells, which systemically circulate, transiently entering peripheral tissues; and resident memory T (TRM) cells, a non-circulating, self-renewing population found in peripheral tissues exposed to the acute pathogenic assault.1,2,3 Tissue-resident CD8 T cells act as sentinels of the adaptive immune system at sites of pathogen entry, rapidly inducing strong local chemokine and cytokine expression when they interact with their cognate antigen, leading to the rapid accumulation of circulating immune cells in the infected tissue.4 Most importantly, TRM in peripheral tissues have been shown to provide nearly sterilizing immunity when present in sufficient numbers, likely through their ability to rapidly respond to the invading pathogen before it can replicate to high titers.5,6,7,8 In the case of the lung, we have shown that the airway CD8 TRM population is sufficient to mediate protection from heterologous influenza challenge, working in concert with the lung parenchyma (LP) CD8 TRM population to rapidly produce cytokines and kill infected cells.9 Importantly, the number of lung CD8 TRM correlates with the efficacy of cellular immunity to influenza.10,11 Despite their importance for cell-mediated protection against respiratory infections, scant knowledge exists as to factors necessary to establish and maintain airway and lung parenchymal CD8 TRM populations.
Upon entering the tissue microenvironment, effector CD8 T cells are exposed to cytokines, such as TGF-β, that induce a transcriptional program leading to TRM differentiation.12,13 TGF-β signaling downregulates T-bet and promotes expression of CD103, the alpha chain of integrin αEβ7, which binds to E-cadherin expressed by epithelial cells and promotes the retention or survival of T cells in the tissue. The transcription factor Krüppel-like factor 2 (KLF2) is also downregulated during TRM differentiation, resulting in decreased expression of the tissue egress molecules S1PR1 and CCR7.14 The inhibition of S1PR1 expression is further reinforced by the expression of CD69 on TRM, which binds to S1PR1 on the cell surface and mediates its degradation.15,16 Together these studies have supported the idea that TRM differentiation is an antigen-independent process mediated by cytokines within the tissue microenvironment. Indeed, CD8 T cells expressing a dominant-negative form of the TGF-β receptor are defective in the establishment of CD103 + TRM in tissues17, and CD8 T cells lacking CD69 expression fail to be retained in the tissue long-term in the skin.18 Furthermore, it has been shown for the skin, gut, and female genital tract that the recruitment of circulating effector CD8 T cells into the tissue is sufficient to induce the differentiation of TRM in an antigen-independent manner.7,19,20,21 In contrast, a recent study has shown that local antigen encounter in the skin was important for the formation of CD8 TRM following vaccinia virus infection.22 Furthermore, it is known the route of infection influences the formation of CD8 TRM in the lung, suggesting that anatomical distribution of antigen may impact local TRM differentiation in the lung.23 However, compared to other mucosal sites, the requirement of localized inflammation and/or local antigen in the tissue for the differentiation of effector T cells into long-lived resident CD8 T-cell memory in the lung airways and lung parenchyma remains unknown.
In the present study, we developed an intramuscular (i.m.) priming and intranasal (i.n.) dosing strategy to dissect the relative contributions of local antigen and inflammation for the establishment of influenza-specific lung CD8 TRM. Similarly to previous studies of vaccination against Mycobacterium tuberculosis and RSV, we find that a non-pulmonary route of influenza priming was unable to induce tissue-resident CD8 T-cell memory in the lung airways and parenchyma despite similar numbers of systemic circulating flu-specific memory CD8 T cells compared to i.n. priming.24,25 Pulmonary inflammation alone was able to draw i.m.-primed effector CD8 T cells into the lung transiently, but these cells failed to persist as lung-resident memory cells. In contrast, combining peptide antigen with pulmonary inflammation resulted in the establishment of antigen-specific airway and LP CD8 TRM populations on par with immune responses generated by a native influenza infection. The ability of effector CD8 T cells to recognize their cognate antigen in the lung resulted in increased and prolonged expression of the tissue-retention markers CD69 and CD103, and increased expression of the collagen-binding integrin VLA-1. Finally, mice in which lung TRM were generated through i.n. peptide administration were protected from lethal heterosubtypic influenza challenge similarly to mice that were primed by native influenza infection. Overall, these results demonstrate that local antigen encounter, and not simply the lung microenvironment itself, is required for the differentiation of antigen-specific effector CD8 T cells into lung TRM.
Materials and methods
Mice
C57BL/6J mice were purchased from The Jackson Laboratory and housed under specific ABSL2 conditions at Emory University. All experiments were completed in accordance with the Institutional Animal Care and Use Committee guidelines of Emory University.
Infections and intranasal vaccination
Intranasal infections were performed with influenza A/HKx31 (H3N2) at 30,000 50% egg infectious doses (EID50); intramuscular infections were performed with influenza A/HKx31 (H3N2) at 1 × 106 EID50.26 Influenza A/PR8 (H1N1) at 6000 EID50 was used for heterologous challenge of vaccinated and infected mice. Peptides used for lung antigen dosing were InfluenzaNP311–325 (QVYSLIRPNENPAHK) and InfluenzaNP366–374 (ASNENMETM) at 5 µg, as indicated. TLR agonist ODN 1826 (CpG) (InVivoGen) was administered at 5 µg to induce inflammation. Peptides and/or TLR agonist were administered in 30 µl of PBS intranasally. For influenza A/PR8 (H1N1) challenge, animals reaching defined endpoints of <75% original weight were humanely killed by 2,2,2-tribromoethanol (Avertin) overdose (600 mg/kg) followed by brachial exsanguination. No other analgesics or anesthetics were administered during the time course.
Intravital cell labeling and cellular isolation
To delineate T cells resident in tissue and those in the vasculature, mice were intravenously injected with a fluorophore-conjugated antibody (1.5 µg of fluorophore-conjugated α-CD3ε antibody in 200 µl PBS) 5 min before killing with Avertin and exsanguination; after which point, tissues were harvested and processed as previously described.9 Bronchoalveolar lavage (BAL) was harvested directly from killed mice; mediastinal lymph nodes and spleen were mechanically dissociated into single cell suspensions; lungs were mechanically dissociated and digested in Collagenase D (Sigma) and DNase (Roche).
Cellular stimulation and intracellular cytokine staining (ICCS)
Mouse lung-derived lymphocytes were stimulated for 5 h with 1 µM InfluenzaNP366–374 (ASNENMETM) with Brefeldin A included in the incubation. Following stimulation, cells were then stained with Zombie Yellow (BioLegend) to exclude dead cells from the downstream analysis. Staining for intracellular cytokines was performed as previously described.26
Flow cytometry
Antibodies and staining reagents used for flow cytometry include Biolegend CD62L [MEL-14], CD8α [53–6.7], CD69 [H1.2F3], CD4 [RM4-5], CD11b [M1/70], CD45 [30/F11]; eBioscience CD11a [M17/4], CD44 [IM7], TNFα [MP6-XT22]; BD Biosciences CD3ε [145-2C11], CD103 [M290], CD49a [Ha31/8], IFN-γ [XMG1.2]. Staining for intracellular markers was performed with the BD cytofix/cytoperm kit according to manufacturer’s instructions. Tetramers used for detection of antigen-specific cells include H-2Db Influenza A NP366–374 (ASNENMETM) and H-2Db Influenza A PA224–233 (SSLENFRAYV) and were provided by the NIH tetramer core. Samples were run on a BD Biosciences LSR II flow cytometer and sorted on an Aria II cell sorter (BD Biosciences) with analysis by FlowJo software.
Data analysis and statistics
All analysis was performed in GraphPad Prism 6. Figures represent mean and SEM unless specified otherwise. The specific statistical test for each analysis is noted in the respective figure legend.
Results
Intramuscular infection fails to generate CD8 TRM in the lung parenchyma and airways
It has been previously shown that the route of infection impacts generation of CD8 TRM within the lung airways.23 However, the requirements for generating lung TRM are still unknown. Given that the majority of vaccination methods employ an i.m. inoculation route, we investigated the ability of an i.m. vs. i.n. infection to establish resident memory CD8 T cells in the lung parenchyma (LP) and lung airways. As i.m. inoculation with influenza virus results in an abortive infection, we adjusted the dose in the i.m. inoculation to generate equal numbers of systemic flu-specific CD8 T cells when compared to an i.n. infection in order to directly compare the ability of these different routes to establish lung TRM (Fig. 1a). Prior to killing, we performed intravital labeling to distinguish cells that were resident in the parenchyma of the lung from those in the pulmonary vasculature (Fig. 1b). At day 45 post-infection (PI), both the i.n. and i.m. routes of infection had established similar numbers of FluNP-specific and FluPA-specific memory CD8 T cells in the spleen and lung vasculature (Fig. 1c, d). In contrast, mice receiving an i.m. infection had significantly fewer flu-specific CD8 TRM in the lung parenchyma and airways compared to i.n.-infected mice (Fig. 1d). This difference was even more dramatic when comparing the number of flu-specific cells in the lung that co-expressed the canonical TRM markers CD69 and CD103 (Fig. 1e). Thus, the ability of an i.m. infection to establish TRM in the lung parenchyma and airways was severely compromised compared to mice that received an i.n. infection despite similar numbers of systemic, circulating flu-specific memory CD8 T cells.

Intramuscular infection results in compromised establishment of resident memory CD8 T cells in the lung parenchyma and airways. a Experimental design. b Representative intravascular staining used to discriminate vascular (i.v. CD3+) and tissue-resident (i.v. CD3−) flu-specific CD8 T cells. c Representative staining of distribution of FluNP-specific CD8 T cells in the airway, lung parenchyma, lung vasculature, and spleen of mice 45 days after IN or IM infection. d The number of FluNP- and FluPA-specific memory CD8 T cells in the lung airways, lung parenchyma (LP), and spleen 45 days after IN (black bar) or IM (white bar) infection. e The number of flu-specific CD8 T cells expressing CD69 and CD103 in the lung airways and lung parenchyma 45 days after IN (black bar) or IM (white bar) infection. Data are shown as the mean ± SEM with 3–5 mice per time point and representative of five individual experiments. Statistical comparisons were performed using an unpaired t-test with Holm–Sidak-corrected multiple comparisons
Local antigen encounter in the lung is required to establish long-lived CD8 TRM in the lung parenchyma and lung airways
The inability of i.m. infection to establish lung TRM may result from a lack of inflammatory signals directing the migration of systemic effector CD8 T cells into the lung tissue. Previous studies have shown that the migration of systemic effector T cells into inflamed skin and genital tract was sufficient to generate local TRM, but the requirements for generating lung TRM are unknown. To test whether local inflammation is sufficient to establish lung TRM, or whether antigen encounter in the tissue is also required, we used a prime-pull strategy to induce local inflammation in the lungs of i.m.-infected mice in the presence or absence of cognate antigen (Fig. 2a). Mice were dosed i.n. with CpG or CpG/Ag seven days after i.m. infection and the number of FluNP-specific CD8 T cells in the resident BAL and lung were analyzed on days 10 and 45. On day 10, 3 days after i.n. dosing, each group showed similar numbers of FluNP-specific CD8 T cells in the BAL and lung (Fig. 2b). However, by day 45 mice given CpG only had a much lower frequency of FluNP-specific lung TRM than either i.n.-infected or i.m.-infected CpG/Ag mice (Fig. 2c). This decreased frequency associated with significant decreases in the total number of FluNP-specific BAL and lung TRM and the number of FluNP-specific lung TRM expressing the canonical residency markers CD69 and CD103 in i.m.-infected mice that were dosed with CpG alone (Figs. 2d,e). In contrast to the tissue-resident cells lung, there was no difference in the number of FluNP-specific memory CD8 T cells in the lung vasculature (LV) or spleen among the treatment groups, but there was a significant decrease number of FluNP-specific memory CD8 T cells in the mediastinal lymph node in i.m.-infected CpG alone mice (Fig. 2f). To further define the requirements for generating lung TRM, we investigated the ability of antigen alone, or CpG/Ag administered intraperitoneally (i.p.), to generate lung TRM following i.m. infection. As shown in Fig. 2g, neither intranasal Ag alone nor i.p. CpG/Ag was sufficient to generate FluNP-specific TRM in the BAL and lung despite similar numbers of number of FluNP-specific memory CD8 T cells in the spleen compared to an i.n. infection. Together these data show that, unlike other mucosal sites, drawing effector CD8 T cells into the lung microenvironment was insufficient to generate TRM. Rather, the establishment of lung TRM required that effector T cells encounter their cognate antigen in the pulmonary environment.

Local antigen encounter in the lung environment is required to establish lung-resident CD8 T-cell memory. a Experimental design to test the relative contributions of inflammation and antigen for establishing lung TRM. b The number of FluNP-specific CD8 T cells in the lung airways (BAL) or lung parenchyma (LR) on day 10 after IN influenza infection (black bar) or after i.n. delivery of CpG (green bar) or CpG/Ag (blue bar) on day 7 to IM-infected mice. c Representative staining from the lungs of the three experimental groups on day 45 post-infection showing the frequency of the bulk CD8+ lung-resident population (left column), the frequency of the lung-resident FluNP-specific cells (middle column), and the frequency of CD69+ D103+ cells. d The number of FluNP-specific cells resident in the lung airways (BAL) or lung parenchyma (LP) on day 45 post-infection. e The number of flu-specific CD8 T cells expressing CD69 and CD103 in the lung airways (BAL) and lung parenchyma (LP) on day 45 post-infection. f The number of flu-specific CD8 T cells in the lung vasculature (LV), mediastinal lymph node (MLN) and spleen on day 45 post-infection. g The number of flu-specific CD8 T cells in the lung airways (BAL), lung parenchyma (LP), and spleen on day 45 post-infection. Data are shown as the mean ± SEM with 3–5 mice per time point and representative of five individual experiments. Statistical comparisons were performed using a two-way ANOVA with Tukey-corrected multiple comparisons
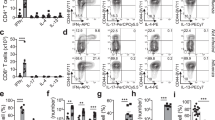
To further support that individual T cells must encounter their cognate antigen in the lung to generate TRM, we analyzed the number of FluPA-specific T cells in the BAL and lungs of mice dosed with CpG or CpG + FluNP peptide. As shown in Fig. 3a, both CpG and CpG/Ag dosed mice show significantly fewer FluPA-specific lung TRM than i.n.-infected mice, and there was no difference in the number of FluPA-specific cells between the CpG and CpG/Ag groups despite the significant increase in FluNP-specific TRM in the CpG/Ag mice. In addition, there is significantly less fold change in the number of FluNP-specific lung TRM from day 10 to day 45 in the CpG/Ag mice compared to the CpG mice (Fig. 3b), whereas there is no difference in the fold change of FluPA-specific lung TRM between these two groups (Fig. 3c). These data demonstrate that the necessity of local antigen recognition for the generation of lung TRM is a cell-intrinsic phenomenon, and that antigen recognition by cells of one specificity do not alter the lung microenvironment to permit cells that have not encountered their antigen in the lung to establish TRM.

The requirement for antigen recognition in establishing lung TRM is cell intrinsic and results in prolonged expression of CD69. a The number of FluPA-specific memory CD8 T cells resident in the lung airways (BAL) or lung parenchyma (LP) on day 45 following IN (black bar), IM + CpG (green bar) or IM + CpG/Ag (blue bar) infection. b, c The fold change in the number of b FluNP-specific CD8 T cells or c FluPA-specific CD8 T cells resident in the lung airways (BAL) or lung parenchyma (LP) from day 10 to day 45 post-infection. d Representative staining of CD69 expression on FluNP-specific CD8 T cells in the lung on days 10 and 17 after IM + CpG (top row) or IM + CpG/Ag (bottom row) infection. e, f The frequency of CD69 + cells among e FluNP-specific or f FluPA-specific lung-resident CD8 T cells on days 10 and 17 following IM + CpG (green bar) or IM + CpG/Ag (blue bar) infection. Data are shown as the mean ± SEM with five mice per time point and representative of three individual experiments. Statistical comparisons were performed (a) using a two-way ANOVA with Tukey-corrected multiple comparisons or (b, c, e, f) using an unpaired t-test with Holm–Sidak-corrected multiple comparisons
Rapid seeding of the lung CD8 TRM pool following pulmonary antigen encounter
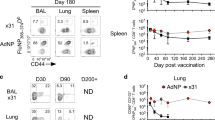
Two potential explanations for the absence of lung TRM in mice receiving only local inflammation are that the lack of local antigen results in: (i) defective initial formation of lung TRM, or (ii) transient formation of lung TRM that rapidly decline and are not maintained into memory. To investigate the initial generation of lung TRM, we compared i.n.-infected, CpG alone, and CpG/Ag i.m.-infected mice between days 7 and 17 after initial infection. All groups showed a robust lung-resident FluNP-specific CD8 T-cell population on days 10 and 14, but only the i.n.-infected and CpG/Ag groups showed a subset of cells expressing CD69 and CD103 by day 14 (Fig. 4a). The total number of resident FluNP-specific CD8 T cells in the BAL and lung on day 10, 3 days following CpG or CpG/Ag dosing, are similar between all the groups (Fig. 4b). However, by day 14 there is a significant divergence in the BAL and lung between the CpG and CpG/Ag treated groups, where the number of FluNP-specific cells in the CpG/Ag group remained similar to the i.n.-infected group, whereas the CpG group showed a rapid decline in numbers. This difference was also observed among BAL and lung FluNP-specific cells expressing CD69 and CD103, as the number of these cells in the CpG/Ag group continued to rise between days 10-14, whereas declining after day 10 in the CpG group (Fig. 4c). Based on this evidence, it appears as if the CpG only group fails to initially generate lung CD8 TRM from the effector T-cell pool in the lung, whereas the addition of local antigen (CpG/Ag) leads to the rapid formation of CD69+/CD103+ long-lived TRM within the lung parenchyma and airways.

Rapid induction of cells with a TRM phenotype following antigen encounter in the lung. a Representative staining of FluNP366–374-specific CD8 T cells resident in the lung parenchyma on days 10 and 14 following IN, IM + CpG, or IM + CpG/Ag infection (top rows) and the expression of CD69 and CD103 on FluNP366–374-specific CD8 T cells in the lung (bottom rows). b The number of FluNP366–374-specific CD8 T cells resident in the lung airways and lung parenchyma (LP) at day 7, 10, 14 and 17 after IN infection (black circle) or IM infection followed by i.n. delivery of CpG (green square) or CpG/Ag (blue triangle) on day 7 to IM-infected mice. c The number of CD69+ CD103+ cells among the FluNP366–374-specific population in the airways and lung parenchyma. Data are shown as the mean ± SEM with five mice per time point and representative of three individual experiments. Statistical comparisons were performed using a two-way ANOVA with Tukey-corrected multiple comparisons and significance is shown comparing the IM + CpG and IM + CpG/Ag groups (p-values (**<0.01; ***<0.001; ****<0.0001))
Pulmonary antigen promotes expression of tissue-retention molecule CD49a on lung CD8 T cells
It has been shown that expression of CD49a, the alpha subunit of the integrin VLA-1, is necessary for protection during a heterologous influenza virus challenge due to its role in retaining memory CD8 T cells in the lung.27 Given the importance of CD49a in facilitating the maintenance of lung memory T cells, we wanted to determine whether antigen encounter in the lung altered CD49a expression. As shown in Fig. 5a and b, CD49a was highly expressed on the total CD8 TRM pool in the lungs of i.n.-infected mice and CpG/Ag i.m.-infected mice, whereas CpG i.m.-infected mice showed low expression of CD49a on the few lung CD8 TRM present in these animals. In addition, the FluNP-specific CD8 TRM in the lung of CpG/Ag i.m.-primed mice had significantly higher expression of CD49a compared to CpG i.m.-infected mice on day 45 (Fig. 5c). Together, these data show that effector CD8 T cells unable to recognize their cognate antigen in the lung fail to maintain or increase expression of tissue-retention molecules, including CD49a, which may account for the inability of localized inflammation to establish lung TRM by itself.

Antigen encounter in the lung promotes the expression of CD49a on tissue-resident memory CD8 T cells. a Representative staining for CD49a on lung-resident CD8 TRM at day 45 following IN (black histogram), IM + CpG (gray histogram) or IM + CpG/Ag (open histogram) infection. b CD49a geometric MFI (gMFI) on total lung-resident memory CD8 T cells from the three groups noted in a at day 45 post-infection. c CD49a gMFI on FluNP366–374-specific lung CD8 TRM 45 days post-infection. Data are shown as the mean ± SEM with 10 mice per group and representative of two individual experiments. Statistical comparisons were performed (b) using a two-way ANOVA with Tukey-corrected multiple comparisons or (c) an unpaired t-test (p-values (**<0.01; ****<0.0001))
Lung CD8 TRM established via intranasal inflammation and antigen are protective against heterologous influenza challenge
Although numbers of FluNP-specific lung CD8 TRM between i.n.-infected and i.m. + CpG/Ag mice were similar, it was unclear if these cells had similar effector functions or were able to mediate similar levels of protective cellular immunity. We isolated lung-resident lymphocytes on day 42 from i.n.-infected and i.m. + CpG/Ag mice, stimulated them with FluNP366–374 peptide, and evaluated their production of TNFα and IFNγ (Fig. 6a). We observed no difference between groups in the ability of lung TRM to produced IFNγ or TNFα alone or in combination (Fig. 6b). Mice were also challenged with influenza PR8 to assess the efficacy of heterosubtypic cellular immunity. As shown in Fig. 6c, i.m. + CpG and i.m. + i.p. CpG/Ag mice showed rapid weight loss following challenge, whereas i.m. + CpG/Ag mice were protected from weight loss similarly to i.n.-infected mice. Survival was also significantly improved in mice i.m. + CpG/Ag mice and was similar to i.n.-primed mice (Fig. 6d). The improved clinical outcome of i.m. + CpG/Ag mice corresponded with decreased viral titers after challenge compared to i.m. + CpG mice, and i.m. + CpG/Ag mice had viral titers similar to i.n.-infected mice. Notably, the administration of CpG during the priming phase did not adversely impact clearance of the challenge virus, as mice only given an i.m. infection and i.m. + CpG mice showed similar viral titers after challenge (Fig. 6f). Thus, establishing lung TRM by intranasal delivery of cognate antigen to the lung environment is able to generate a robust and protective CD8 TRM population in the airways and lung parenchyma that provides protective cellular immunity rivaling that observed following a native respiratory infection.

Lung TRM generated by IM infection + CpG/Ag provide robust protection against heterologous influenza challenge. a Representative staining of IFNγ and TNFα production by lung-derived CD8 T cells from mice 42 days after IN infection or IM infection + CpG/Ag following stimulation with FluNP366–374 peptide; gated on CD11b-/CD4-/CD8+/CD44+ cells. b Percentage of the FluNP366–374-specific CD8 T cells, described in (a), that express IFNγ, TNFα, or both cytokines. c Weight loss over 7 days of mice challenged with 10 LD50 PR8 45 days following IN (black circle), IM + CpG (green square), IM + CpG/Ag (blue triangle), or IM + IP CpG/Ag (red triangles) infection. d Survival curve of the same mice for which weight loss was measured. e Viral load (influenza PA RNA copies/100 mg of lung tissue) on day 6 following PR8 influenza challenge of IN (black circle), IM + CpG (black square) or IM + CpG/Ag (black triangle) primed mice. Mice were challenged 45 days after initial priming. Data shown are compiled from two independent experiments. f Viral load (influenza PA RNA copies/100 mg of lung tissue) on day 6 following PR8 influenza challenge of IM only (black squares) or IM + CpG primed mice, compiled from two independent experiments. Data shown as the mean ± SEM with 5–8 mice per time point and representative of 2–3 independent experiments. Statistical comparisons were performed using a two-way ANOVA with b Sidak or c Tukey-corrected multiple comparisons, d Log-rank Mantel–Cox test, e one-way ANOVA with Tukey-corrected multiple comparisons, and f and unpaired t-test (p-values (**<0.01; ****<0.0001))
Discussion
Although cellular immunity in the lung has been shown to be effective in limiting the severity of respiratory viral infections, the factors that regulate the establishment of lung-resident T-cell memory have not been defined. In the present study, we have shown that exposure to the lung environment itself is insufficient to promote the differentiation of effector CD8 T cells into long-lived lung TRM. Rather, TRM differentiation in the lung airways and lung parenchyma required cognate antigen recognition by effector CD8 T cells in the lung environment. Exposure to antigen in the lung tissue resulted in prolonged expression of CD69 and increased expression of VLA-1, which together promote the retention of CD8 T cells in the lung. Finally, we showed that lung TRM generated via intranasal inflammation and antigen administration are highly protective against heterologous influenza challenge, confirming the importance of lung TRM for combating respiratory viral infections and demonstrating the utility of the intranasal dosing strategy for inducing efficacious cellular immunity in the lung.
The factors that control the differentiation of tissue-resident memory T cells have been intensely studied. In contrast to mechanisms described for the skin and FRT, where administration of local inflammatory signals was sufficient to establish a robust and protective TRM population from a systemic effector T-cell pool, both local antigen and inflammation are required for the establishment of robust and protective LP and airway TRM populations.5,7 The unique requirements among different tissues raises interesting questions regarding whether there may be tissue-specific factors that regulate the induction of a tissue residency program in CD8 T cells. Prior studies have found that common residency cues, such as TGFβ signaling, are also required for the establishment of lung CD8 TRM.28,29 However, the insufficiency of local cytokine signals by themselves to induce a robust population of lung TRM demonstrates that additional signals are required in the lung. Several possible clues for the unique requirements of lung TRM come from the comparison of flu-specific CD8 T cells in the lung that do or do not encounter their cognate antigen. In the lungs of i.m. + CpG/Ag treated mice, fluNP-specific cells (which can encounter their antigen in the lung) maintained prolonged expression of CD69 compared to fluPA-specific cells (which do not encounter their antigen in the lung). Owing to the highly vascularized nature of the lung, it is possible that extended antagonism of S1P1R by CD69 is necessary to inhibit the S1P-mediated exit of T-cell into the bloodstream prior to the establishment of tissue residency. We also observed that only flu-specific CD8 T cells encountering their cognate antigen in the lung express CD49a, and the absence of this integrin has been previously shown to decrease the number of flu-specific CD8 T cells in the lung.27 Likely, it is the combined expression of these, and potentially other, molecules that enable CD8 T cells encountering their antigen in the local environment to establish long-term resident memory in the lung.
Another interesting possibility our findings suggest is the potential for distinct lung antigen presenting cell (APC) subsets to regulate the establishment of lung TRM. It was previously shown that targeting vaccination to CD103+ dendritic cells in the lung could promote the establishment of lung CD8 TRM,29 and a similar study found that alveolar macrophages expressing cognate antigen could promote lung TRM.30 Although all APC subsets in the lung can present antigen to CD8 T cells, it is not known whether all APC subsets harbor influenza antigens following viral clearance when we first observe the appearance of flu-specific CD8 T cells with a TRM (CD69+ CD103+) phenotype. Furthermore, it is possible that specific lung APC subsets, in addition to presenting antigen, may also supply additional signals through cytokines or cell surface ligands that promote TRM development. Thus, future investigations of T cell—APC interactions in the lung after viral clearance are needed determine whether effector CD8 T cells must interact with specific APC subsets displaying antigen in the lung to develop into TRM.
Current research continues to reveal that naturally occurring cross-reactive T cells to influenza virus have the capacity to reduce morbidity and mortality, resulting in largely asymptomatic responses in some cases, upon pathogen challenge.6,31,32 Although cellular immunity has been shown to be effective against heterologous influenza challenge, the efficacy of this protection wanes over time due to the loss of flu-specific lung TRM, and thus the development of approaches to transiently increase lung TRM numbers could prove effective in boosting heterosubtypic immunity during influenza epidemics. The intranasal dosing strategy we describe here could be employed to transiently boost the number of flu-specific lung TRM, and as we have demonstrated the lung TRM generated through this approach are protective against heterosubtypic influenza challenge. In addition, this approach could be useful for establishing lung TRM populations against a number of respiratory pathogens. Antigen-specific memory T cells in the lung are known to be important for protection against respiratory pathogens such as RSV and Mycobacterium tuberculosis, and the addition of an intranasal antigen-dosing regimen may enhance protective immunity following parenteral vaccination through the induction of lung TRM without direct infection of the respiratory system.
In summary, we have shown that both local inflammation and antigen are required to generate a robust pathogen-specific lung and airway CD8 TRM population from systemic effector CD8 T cells, whereas local inflammation alone is not sufficient. Antigen encounter in the lung prolonged expression of the tissue-retention marker CD69 and induced the expression of the collagen-binding integrin CD49a in a cell-intrinsic manner. Finally, lung TRM generated through the pulmonary delivery of inflammation and antigen provided robust protection against secondary challenge. These findings demonstrate an approach to increase the number of lung TRM and may inform future vaccine design against respiratory pathogens.
References
Mueller, S. N., Gebhardt, T., Carbone, F. R. & Heath, W. R. Memory T cell subsets, migration patterns, and tissue residence. Annu Rev. Immunol. 31, 137–161 (2013).
Schenkel, J. M. & Masopust, D. Tissue-resident memory T cells. Immunity 41, 886–897 (2014).
Woodland, D. L. & Kohlmeier, J. E. Migration, maintenance and recall of memory T cells in peripheral tissues. Nat. Rev. Immunol. 9, 153–161 (2009).
Schenkel, J. M., Fraser, K. A., Vezys, V. & Masopust, D. Sensing and alarm function of resident memory CD8(+) T cells. Nat. Immunol. 14, 509–513 (2013).
Mackay, L. K. et al. Long-lived epithelial immunity by tissue-resident memory T (TRM) cells in the absence of persisting local antigen presentation. Proc. Natl Acad. Sci. USA 109, 7037–7042 (2012).
McMaster, S. R. et al. Memory T cells generated by prior exposure to influenza cross react with the novel H7N9 influenza virus and confer protective heterosubtypic immunity. PLoS ONE 10, e0115725 (2015).
Shin, H. & Iwasaki, A. A vaccine strategy that protects against genital herpes by establishing local memory T cells. Nature 491, 463–467 (2012).
Steinert, E. M. et al. Quantifying Memory CD8 T. Cells Reveals Reg. Immunosurveill. Cell 161, 737–749 (2015).
McMaster, S. R., Wilson, J. J., Wang, H. & Kohlmeier, J. E. Airway-resident memory CD8 T cells provide antigen-specific protection against respiratory virus challenge through rapid IFN-gamma production. J. Immunol. 195, 203–209 (2015).
Liang, S., Mozdzanowska, K., Palladino, G. & Gerhard, W. Heterosubtypic immunity to influenza type A virus in mice. Effector mechanisms and their longevity. J. Immunol. 152, 1653–1661 (1994).
Wu, T. et al. Lung-resident memory CD8 T cells (TRM) are indispensable for optimal cross-protection against pulmonary virus infection. J. Leukoc. Biol. 95, 215–224 (2014).
Mackay, L. K. et al. The developmental pathway for CD103(+)CD8+tissue-resident memory T cells of skin. Nat. Immunol. 14, 1294–1301 (2013).
Zhang, N. & Bevan, M. J. Transforming growth factor-beta signaling controls the formation and maintenance of gut-resident memory T cells by regulating migration and retention. Immunity 39, 687–696 (2013).
Skon, C. N. et al. Transcriptional downregulation of S1pr1 is required for the establishment of resident memory CD8+T cells. Nat. Immunol. 14, 1285–1293 (2013).
Pham, T. H., Okada, T., Matloubian, M., Lo, C. G. & Cyster, J. G. S1P1 receptor signaling overrides retention mediated by G alpha i-coupled receptors to promote T cell egress. Immunity 28, 122–133 (2008).
Shiow, L. R. et al. CD69 acts downstream of interferon-alpha/beta to inhibit S1P1 and lymphocyte egress from lymphoid organs. Nature 440, 540–544 (2006).
Casey, K. A. et al. Antigen-independent differentiation and maintenance of effector-like resident memory T cells in tissues. J. Immunol. 188, 4866–4875 (2012).
Mackay, L. K. et al. Cutting edge: CD69 interference with sphingosine-1-phosphate receptor function regulates peripheral T cell retention. J. Immunol. 194, 2059–2063 (2015).
Bergsbaken, T. & Bevan, M. J. Proinflammatory microenvironments within the intestine regulate the differentiation of tissue-resident CD8(+) T cells responding to infection. Nat. Immunol. 16, 406–414 (2015).
Gebhardt, T. et al. Memory T cells in nonlymphoid tissue that provide enhanced local immunity during infection with herpes simplex virus. Nat. Immunol. 10, 524–530 (2009).
Masopust, D. et al. Dynamic T cell migration program provides resident memory within intestinal epithelium. J. Exp. Med. 207, 553–564 (2010).
Khan, T. N., Mooster, J. L., Kilgore, A. M., Osborn, J. F. & Nolz, J. C. Local antigen in nonlymphoid tissue promotes resident memory CD8+T cell formation during viral infection. J. Exp. Med. 213, 951–966 (2016).
Takamura, S. et al. The route of priming influences the ability of respiratory virus-specific memory CD8+T cells to be activated by residual antigen. J. Exp. Med. 207, 1153–1160 (2010).
Morabito, K. M. et al. Intranasal administration of RSV antigen-expressing MCMV elicits robust tissue-resident effector and effector memory CD8+T cells in the lung. Mucosal Immunol. 10, 545–554 (2017).
Wang, J. et al. Single mucosal, but not parenteral, immunization with recombinant adenoviral-based vaccine provides potent protection from pulmonary tuberculosis. J. Immunol. 173, 6357–6365 (2004).
Kohlmeier, J. E., Cookenham, T., Roberts, A. D., Miller, S. C. & Woodland, D. L. Type I interferons regulate cytolytic activity of memory CD8(+) T cells in the lung airways during respiratory virus challenge. Immunity 33, 96–105 (2010).
Ray, S. J. et al. The collagen binding alpha1beta1 integrin VLA-1 regulates CD8 T cell-mediated immune protection against heterologous influenza infection. Immunity 20, 167–179 (2004).
Hu, Y., Lee, Y. T., Kaech, S. M., Garvy, B. & Cauley, L. S. Smad4 promotes differentiation of effector and circulating memory CD8 T cells but is dispensable for tissue-resident memory CD8 T cells. J. Immunol. 194, 2407–2414 (2015).
Wakim, L. M., Smith, J., Caminschi, I., Lahoud, M. H. & Villadangos, J. A. Antibody-targeted vaccination to lung dendritic cells generates tissue-resident memory CD8 T cells that are highly protective against influenza virus infection. Mucosal Immunol. 8, 1060–1071 (2015).
Macdonald, D. C. et al. Harnessing alveolar macrophages for sustained mucosal T-cell recall confers long-term protection to mice against lethal influenza challenge without clinical disease. Mucosal Immunol. 7, 89–100 (2014).
Hayward, A. C. et al. Natural T cell-mediated protection against seasonal and pandemic influenza. Results of the flu watch cohort study. Am. J. Respir. Crit. Care Med. 191, 1422–1431 (2015).
Sridhar, S. et al. Cellular immune correlates of protection against symptomatic pandemic influenza. Nat. Med. 19, 1305–1312 (2013).
Acknowledgements
We would like to thank the NIH Tetramer Core Facility (contract HHSN272201300006C) for provision of MHC I tetramers. This work was supported by National Institutes of Health grant HL122559 (to J.E.K.) and Centers of Excellence in Influenza Research and Surveillance Contract HHSN272201400004C (to J.E.K.). S.R.M. was supported by National Institutes of Health Grants F30 HL118954 and T32 AI007610. P.R.D. was supported by National Institutes of Health Grant F31 AI124611.
Author information
Authors and Affiliations
Contributions
S.R.M., A.N.W., P.R.D., S.L.H, E.K.C., and J.E.K. performed experiments and analyzed data. T.L.D. provided assistance and contributed reagents and materials. S.R.M. and J.E.K. designed the study and wrote the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing financial interests.
Additional information
Publisher's note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
McMaster, S.R., Wein, A.N., Dunbar, P.R. et al. Pulmonary antigen encounter regulates the establishment of tissue-resident CD8 memory T cells in the lung airways and parenchyma. Mucosal Immunol 11, 1071–1078 (2018). https://doi.org/10.1038/s41385-018-0003-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41385-018-0003-x
This article is cited by
-
Prevention of respiratory virus transmission by resident memory CD8+ T cells
Nature (2024)
-
T cell receptor signaling strength establishes the chemotactic properties of effector CD8+ T cells that control tissue-residency
Nature Communications (2023)
-
Intranasal administration of a recombinant RBD vaccine induces long-term immunity against Omicron-included SARS-CoV-2 variants
Signal Transduction and Targeted Therapy (2022)
-
Spatial, temporal and molecular dynamics of swine influenza virus-specific CD8 tissue resident memory T cells
Mucosal Immunology (2022)
-
Tissue resident memory T cells in the respiratory tract
Mucosal Immunology (2022)
