
Abstract
We report definitive diagnosis and effective chenodeoxycholic acid (CDCA) treatment of two Japanese children with 3β-hydroxy-Δ5-C27-steroid dehydrogenase/isomerase deficiency. Findings of cholestasis with normal serum γ-glutamyltransferase activity and total bile acid concentration indicated the need for definitive bile acid analysis. Large amounts of 3β-hydroxy-Δ5bile acids were detected by gas chromatography-mass spectrometry. HSD3B7 gene analysis using peripheral lymphocyte genomic DNA from the patients and their parents identified four novel mutations of the HSD3B7 gene in the patients. One had a homozygous mutation, 314delA; the other had compound heterozygous mutations: V132F, T149I, and 973_974insCCTGC. Interestingly, the second patient's mother had V132F and T149I mutations in one allele. Excessive 3β-hydroxy-Δ5-bile acids such as 3β,7α-dihydroxy- and 3β,7α,12α-trihydroxy-5-cholenoic acids were detected in the first patient's urine; the second patient's urine contained large amounts of 3β-hydroxy-5-cholenoic acid. Liver dysfunction in both patients decreased with ursodeoxycholic acid treatment, but unusual bile acids were still detected. Normalization of the patients' liver function and improvement of bile acid profiles occurred with CDCA treatment. Thus, we found mutations in the HSD3B7 gene accounting for autosomal recessive neonatal cholestasis caused by 3β-hydroxy-Δ5-C27-steroid dehydrogenase/isomerase deficiency. Early neonatal diagnosis permits initiation of CDCA treatment at this critical time, before the late cholestatic stage.
Similar content being viewed by others

Main
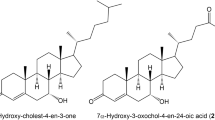
Deficiency of 3β-hydroxy-Δ5-C27-steroid dehydrogenase/isomerase (3β-HSD) was first described by Clayton et al. in 1987 (1). This inborn error of bile acid synthesis is very rare and shows autosomal recessive inheritance. The main findings in 3β-HSD deficiency are low or normal concentrations of total bile acid and normal activity of γ-glutamyltransferase (GGT) in serum, as well as absence of pruritus despite conjugated hyperbilirubinemia, elevated alanine aminotransferase (ALT), and fatty stools. In the synthesis of bile acids from cholesterol, 3β-HSD catalyzes the second of a series of reactions leading to excretion of 3β,7α-dihydroxy-5-cholenoic acid (Δ5-3β,7α-diol) and 3β,7α,12α-trihydroxy-5-cholenoic acid (Δ5-3β,7α,12α-triol) in the urine.
In the first reported patient, complete absence of 3β-HSD activity was found by Buchmann et al. in 1990 (2) based on the study of cultured fibroblasts. In 2000, Schwarz et al. (3) reported that the same patient had a homozygous mutation representing a 2-bp deletion in exon 6 of the 3β-HSD gene (HSD3B7) on chromosome 16p11.2-12. The human HSD3B7 gene contains six coding exons and encodes 369 amino acids; so far, 13 distinct mutations causing 3β-HSD deficiency have been reported (4,5).
Here, we report genetic analyses of two Japanese patients with 3β-HSD deficiency: one previously reported patient (6,7) was diagnosed with 3β-HSD deficiency by bile acid analysis and the other newly reported patient showed different results in the bile acid analysis. Here, we describe definitive diagnosis by bile acid analysis using gas chromatography-mass spectrometry (GC-MS) and effective chenodeoxycholic acid (CDCA) treatment in two patients with 3β-HSD deficiency.
This study was approved by the Kurume University Review Board, and informed consent for the patient report including bile acid and gene analysis was obtained from the two patients and four parents.
PATIENTS AND METHODS
Patient 1.
The family history and initial presentation of this patient have been described previously (6). This 2-mo-old male infant underwent medical therapy for cholestasis and liver dysfunction [total bilirubin (T Bil), 9.3 mg/dL; direct bilirubin (D Bil), 5.9 mg/dL; aspartate aminotransferase (AST), 587 IU/L; ALT, 596 IU/L; GGT, 23 IU/L; prothrombin time, 12.2 s] using ursodeoxycholic acid (UDCA, 12.5 mg/kg/d). Hepatic histologic findings indicated giant cell hepatitis with fibrosis. During treatment, cholestasis and liver dysfunction improved. However, bile acid profiles in urine did not change, indicating that hepatotoxic 3β-hydroxy-Δ5-bile acids such as Δ5-3β,7α-diol and Δ5-3β,7α12α-triol accumulated in the patient's hepatocytes despite UDCA treatment. After treatment with CDCA (8.3 mg/kg/d), however, concentrations of 3β-hydroxy-Δ5-bile acids significantly decreased, suggesting that CDCA treatment of this disease would be useful in preventing cirrhosis (7). At present, the patient who is 12-yr-old is in good condition without liver dysfunction or excessive 3β-hydroxy-Δ5-bile acids in urine or serum on his current CDCA dose of 5.9 mg/kg/d. The patient did not have a follow-up liver biopsy performed.
Patient 2.
A male infant with a birth weight of 3310 g was delivered by spontaneous vaginal delivery without complications at a GA of 38 wk, after an uneventful pregnancy. He was noted to have jaundice at the age of 1 and 3 mo. The jaundice was mild when first noted and it gradually worsened. At the age of 6 mo, the patient was referred to Toho University Hospital because of hyperbilirubinemia and liver dysfunction (T Bil, 5.2 mg/dL; D Bil, 3.1 mg/dL; AST, 362 IU/L; ALT, 384 IU/L; GGT, 33 IU/L; total bile acids, 0.7 μM; and prothrombin activity, 21%). Initial physical examination on admission was nearly unremarkable, without hepatomegaly, obvious jaundice, or dark urine. Serial technetium-99m (99mTC)-DISIDA cholescintigraphy visualized intestinal radioactivity. After the bile acid analysis, we started UDCA treatment (19 mg/kg/d), after which T Bil and ALT gradually decreased to the normal range, from 5.2 mg/dL and 384 IU/L to 0.3 mg/dL and 34 IU/L, respectively (Fig. 1). We chose not to perform a liver biopsy because the patient was in good health without liver dysfunction. After clinical diagnosis and UDCA treatment, we substituted CDCA treatment (15.6 mg/kg/d). Subsequently, liver function test results have remained within the normal range.

Clinical course of patient 2. Responses of the serum D Bil (▪) and ALT (▴) to treatment with UDCA and CDCA are shown.
The initial bile acid analysis detected large amounts of 3β-hydroxy-5-cholenoic acid (Δ5-3β-ol), representing 84% of total urinary bile acids (Table 1). This led us to suspect oxysterol 7α-hydroxylase deficiency (8,9), but we could not detect a mutation of the CYP7B1 gene. In the second urinary bile acid analysis during UDCA treatment, we detected large proportions of 3β-hydroxy-Δ5-bile acids such as Δ5-3β-ol and Δ5-3β,7α,12α-triol, when UDCA was excluded from calculations (Table 1). We ultimately diagnosed this patient with 3β-HSD deficiency.
After CDCA treatment, the concentration of 3β-hydroxy-Δ5-bile acids in urine decreased significantly during 4 mo, from 7.3 to 2.8 μmol/mmol creatinine (Cr) (Table 1).
The patients had neither steatorrhea nor pruritus. The parents of patients 1 and 2 were all in good health, without liver dysfunction.
Qualitative and quantitative bile acid analysis.
Serum and urine samples were collected and stored at −25°C until analysis. Concentrations of individual bile acids in the urine were corrected for Cr concentration and expressed as micromoles per millimoles of Cr.
After synthesizing relevant unusual bile acids such as 3β-hydroxy-Δ5 (10), 3-oxo-Δ4 (11), and allo-bile acids (11), which occur in inborn errors of bile acid synthesis, analysis of the bile acids in urine and serum was undertaken by GC-MS using selected ion monitoring of characteristic fragments of methylester-dimethylethylsilyl ether-methoxime derivatives of bile acids as described previously (11). Before GC-MS analysis, the samples were prepared by enzymatic hydrolysis (cholylglycine hydrolase, 30 U) and solvolysis (sulfatase, 150 U; Sigma Chemical Co. Chemical, St. Louis, MO). We did not use N-acetylglucodamine.
Genetic analysis.
With informed consent, HSD3B7 gene analysis was performed using genomic DNA from peripheral lymphocytes from the two patients and four parents, as well as 100 healthy individuals using a QIAamp Mini Kit (Qiagen, Hilden, Germany). DNA fragments spanning the six coding regions of the HSD3B7 gene were amplified by PCR using Gene Taq (Nippon Gene, Toyama, Japan) and five sets of primers to obtain DNA fragments of the optimal length for direct sequence analysis (Table 2). The PCR program included an initial denaturation step at 94°C for 3 min, followed by 30 cycles with denaturation at 94°C for 1 min, annealing at 62°C for 1 min, and extension at 72°C for 1 min. A final extension step of 72°C for 10 min was performed using a T-Gradient Thermoblock (Biometra, Goettingen, Germany).
After enzyme processing with ExoSAP-IT (USB, Cleveland, OH), direct sequencing of the amplified PCR products was carried out with a DTCS Quick Start Kit (Beckman Coulter, Fullerton, CA) according to the manufacturer's protocol, using the same primers as for PCR amplification. The sequencing reaction product was analyzed electrophoretically using an SEQ2000XL analyzer (Beckman Coulter, Brea, CA).
After the four putative mutations were found in the patients, their parents and 100 healthy individuals were screened for these four mutations by direct sequence analysis.
RESULTS
Patient profile.
Both patients were diagnosed with 3β-HSD deficiency by bile acid profiles and HSD3B7 gene analysis. Liver dysfunction, such as T Bil, D Bil, AST, and ALT, improved in both patients with UDCA treatment (Ref. 7 and Fig. 1); however, the excess level of 3β-hydroxy-Δ5-bile acid in urine did not change. After CDCA treatment, concentrations of 3β-hydroxy-Δ5-bile acids in urine and serum gradually decreased to the normal range. With CDCA treatment, these patients have maintained good condition without liver dysfunction, showing normal bile acid profiles with no 3β-hydroxy-Δ5-bile acids detected.
Biochemical identification of the inborn error of bile acid synthesis.
Results of urine and serum bile acid analysis for patient 2 are shown in Table 1. The serum bile acid concentration was normal in the initial analysis. We detected large amounts of Δ5-3β-ol in serum and urine (65% and 84% of total bile acids, respectively) as well as an evidence of oxysterol 7α-hydroxylase deficiency in the initial bile acid analysis. In a second bile acid analysis during UDCA treatment, the main bile acid in serum was UDCA (100% of total bile acids). The main bile acid in urine was UDCA (73% of total bile acids), and we detected small amounts of 3β-hydroxy-Δ5-bile acids (13% of total bile acids), such as Δ5-3β-ol, Δ5-3β,7α-diol and Δ5-3β,7α,12α-triol, in urine. When we excluded urinary UDCA from consideration at the second bile acid analysis, the main bile acids in urine were 3β-hydroxy-Δ5-bile acids (47% of total bile acids). After diagnosis, UDCA treatment was changed to CDCA treatment, after which the concentrations of 3β-hydroxy-Δ5-bile acids in urine gradually decreased.
Patient 1 showed large amounts of 3β-hydroxy-Δ5-bile acids (93% of total bile acids), such as Δ5-3β,7α-diol and Δ5-3β,7α,12α-triol, in urine and serum (7) at the age of 18 mo.
Identification of HSD3B7 gene defects.
We identified four novel mutations in these two patients. In patient 1, a single homozygous mutation was found in exon 2, at nucleotide number 314, deletion A (314delA), resulting in a frameshift leading to formation of a stop codon at position 112. The mutation was detected in heterozygous form in the parents, whereas being absent in 100 healthy individuals (Fig. 2).

Pedigree for patient 1 shown with genomic DNA sequences in exon 2 of the HSD3B7 gene in this patient, his parents, and a control. The arrows identify homozygous 314delA in the patient, heterozygous (314delA/normal) in his parents, and intact A in a control subject. The reverse strand sequence shows the same result. This represents a 314delA mutation causing a frameshift. Such a nucleotide deletion was not observed in 100 controls.
Patient 2 showed three heterozygous mutations. The first was in exon 3, at nucleotide 412, representing a G-to-T substitution, causing an amino acid change from valine to phenylalanine (V132F). The second mutation was in exon 4, at nucleotide number 464, representing a single C-to-T substitution, leading to an amino acid change from threonine to isoleucine (T149I). The third mutation in exon 6, between nucleotides 973 and 974, was a 5-bp insertion, CCTGC (973_974insCCTGC), causing a frameshift leading to formation of a stop codon at position 321. Interestingly, both V132F and T149I mutations in heterozygous form were detected in the patient's mother but were absent in the father and in 100 healthy individuals. A heterozygous 973_974insCCTGC mutation was detected in the father but was absent in the mother and in the 100 healthy individuals (Fig. 3).

Pedigree for patient 2 shown with genomic DNA sequences in exons 3, 4, and 6 of the HSD3B7 gene in this patient, his patients, and a control. The arrow in exon 3 identifies G/T in the patient and his mother, but G in his father and a control subject. The reverse strand sequence shows the same result. This represents a GTT-to-TTT mutation, affecting valine at position 132, where it is replaced by phenylalanine. The arrow in exon 4 identifies C/T in the patient and his mother, but C in his father and a control subject. The reverse strand sequence shows the same results. This represents an ACC-to-ATC mutation, affecting threonine at position 149, where it is replaced by isoleucine. Such nucleotide substitutions were not observed in 100 controls. The arrows and squares in exon 6 identify nucleotide number 974 (T/C) and heterozygous insertion of 5 bp, CCTGC, in the patient and his father; in his mother and a control subject, only T is present. The reverse strand sequence shows the same result. This represents a 973_974insCCTGC mutation causing a frameshift. Such a nucleotide insertion was not observed in 100 controls.
Above nucleotide numbers indicating positions of individual mutations are based on those determined from human 3β-HSD cDNA (GenBank accession no. AF277719).
DISCUSSION
In the clinical course of our two patients with 3β-HSD deficiency, UDCA treatment was very effective for treating conjugated hyperbilirubinemia and elevation of aminotransferase, such as ALT (Ref. 7, Fig. 1), in addition for treating idiopathic neonatal hepatitis. Therefore, the pediatricians may misdiagnose patients as having idiopathic neonatal hepatitis if UDCA restores apparent good health. We encountered these patients with normal values for GGT, total bile acids in serum, and suspected 3β-HSD deficiency. We also obtained prompt analysis of bile acids in serum and urine using GC-MS to detect any inborn errors of bile acid synthesis (12). Previous reports described two adult patients with 3β-HSD deficiency who were diagnosed in this manner (5,13).
Therefore, in a cholestatic patient, if the serum GGT activity is normal and the total serum bile acid concentration determined using 3α-hydroxysteroid dehydrogenase is normal or low, one needs to analyze bile acids in serum and urine using methods such as GC-MS.
In the initial bile acid analysis, we detected large amounts of 3β-hydroxy-Δ5-bile acids such as Δ5-3β-ol in serum and urine from patient 2, a finding also reported in oxysterol 7α-hydroxylase deficiency (8,9). We speculate that the main pathway of bile acid synthesis was the acidic pathway, based on the results of initial bile acid analysis in serum and urine. Low or absent activity of cholesterol 7α-hydroxylase enzyme in this patient reflected previous observations that cholesterol 7α-hydroxylase enzyme activity physiologically is low or absent in fetal and neonatal life (8,14), and that low or absent activity of oxysterol 7α-hydroxylase enzyme in this patient could be physiologic (as in knockout mice lacking a functional cholesterol 7α-hydroxylase enzyme). Cholestasis precedes up-regulation of oxysterol 7α-hydroxylase activity in the acidic pathway (15,16). Oxysterol 7α-hydroxylase activity was first detected in 3- to 4-wk-old mice and remained detectable in the livers of older mice (16). We detected increased 3β-hydroxy-Δ5-bile acids such as Δ5-3β,7α-diol and Δ5-3β,7α,12α-triol in our patient's second urinary bile acid analysis (Table 1, Fig. 4).

Effect of the defect of 3β-HSD. Reduced primary bile acid synthesis from cholesterol and increased synthesis of 3β-hydroxy-Δ5-bile acids are shown in the flow chart of the classic and acidic pathways.
CDCA treatment was recommended for patients with 3β-HSD deficiency by Ichimiya et al. (17,18), who reported that treatment with CDCA was very effective in improving the clinical status and liver function, reflecting reduced cholesterol catabolism because of inhibition of cholesterol 7α-hydroxylase. As a result, hepatotoxic 3β-hydroxy Δ5 bile acids were decreased. The treatment should also improve absorption of cholesterol from the gut. Together, these two mechanisms might contribute to an increased serum cholesterol concentration. The importance of bile secretion stimulation for avoiding hepatotoxicity could be evaluated by treatment with UDCA, which does not inhibit cholesterol 7α-hydroxylase and therefore does not prevent further synthesis of hepatotoxic 3β-hydroxy Δ5 bile acids (18,19). Even with short-term UDCA treatment available, primary bile acid treatment should be the first choice upon definitive diagnosis. Actually, our patients showed less liver dysfunction with short-term UDCA treatment (Ref. 7, Fig. 1). After diagnosis was made using GC-MS bile acid analysis, both patients, especially the first, maintained good condition without liver dysfunction. Unusual bile acids such as 3β-hydroxy-Δ5-bile acids gradually decreased on long-term CDCA treatment. Subramaniam et al. (20) support cholic acid (CA) and CDCA treatment in 3β-HSD deficiency, but CDCA cannot be used when patients with 3β-HSD deficiency present late with chronic liver disease, at which point CDCA can be hepatotoxic. Alternatively, Jacquemin et al. (21) and Gonzales et al. (22) found oral CA treatment to be safe and effective in treating most common inborn errors of bile acid synthesis, including 3β-HSD deficiency. Therefore, in 3β-HSD deficiency, CA treatment may be better because CA activates negative feedback regulation of bile acid synthesis to inhibit production of hepatotoxic metabolites and is not itself hepatotoxic. Unfortunately, however, CA is not available for clinical use in Japan. We think that CDCA treatment may be effective for 3β-HSD deficiency because of the high potency of CDCA in suppressing bile acid synthesis relative to CA (23–25).
Previous reports of mutations in the HSD3B7 gene in 17 patients identified 13 distinct mutations causing 3β-HSD deficiency (4,5). This enzyme deficiency has been characterized as showing autosomal recessive transmission. Here, we describe genetic analysis of the HSD3B7 gene in two patients with 3β-HSD deficiency. Patient 1 had a homozygous 314delA mutation. The protein encoded by the 314delA mutation is composed of 98 amino acids from the normal protein fused to a 13-residue extension. Patient 2 had three heterozygous mutations: V132F, T149I, and 973_974insCCTGC. The protein encoded by the 973_974insCCTGC mutation is composed of 318 amino acids from the normal protein fused to a 2-residue extension. We screened for the two potentially informative substitution mutations, V132F and T149I, in 100 healthy individuals by direct sequence analysis; neither was found. As for interspecies comparisons corresponding protein, the valine 132 and threonine 149 residues are conserved among humans, cattle, rats, mice, and zebrafish as well as in the previously reported missense mutations G19S, E147 K, and S162R (4,5). Moreover, amino acids adjoining valine 132 and threonine 149 are conserved across species (Fig. 5), which suggests an important catalytic or structural role in the dehydrogenase/isomerase. Accordingly, we concluded that either or both of the V132F and T149I mutations could have contributed to loss of 3β-HSD enzyme function in the proband, considering that patient 2 received one allele with V132F and T149I mutations from the mother, whereas the other allele from his father contained 973_974insCCTGC, also affecting the HSD3B7 gene. His mother was asymptomatic despite having the two missense mutations because both occurred together on only one allele, representing heterozygosity. In the two patients, in this study, we identified a total of four novel mutations.

Aligned amino acid sequences for HSD3B7, comparing human with bovine, rat, mouse, and zebrafish sequences. Amino acids identical with those in humans are in black as opposed to gray letters. Amino acids are numbered at the right. Black arrows identify the two novel missense mutations, V132F and T149I, described in this study. Gray arrows identify previously reported missense mutations, G19S, E147K, and S162R. GenBank accession numbers for human, cattle, rats, mice, and zebrafish are NM_025193, BC105259, NM_139329, BC132605, and NM_199809, respectively.
Patient 1 was a homozygote and patient 2 was a compound heterozygote. Both parents of each patient were the heterozygous for a mutation, strongly suggesting that the patient inherited one or more mutated genes from each parent.
Finally, the results of HSD3B7 gene analysis led us to identify four novel mutations in the HSD3B7 gene that can underlie 3β-HSD deficiency, an autosomal recessive form of neonatal cholestasis. Because the liver function in this disease is improved by UDCA treatment, patients may be misdiagnosed with idiopathic neonatal hepatitis. However, a low or normal serum concentration of total bile acid and a normal serum GGT concentration in a neonate with cholestasis should lead us to suspect inborn errors of bile acid synthesis such as 3β-HSD deficiency. After diagnosis of 3β-HSD deficiency, we recommend prompt initiation of primary bile acid treatment using CDCA (and/or CA, where available) in the early neonatal period, before the late stage of chronic cholestatic liver dysfunction.
Abbreviations
- 3β-HSD:
-
3β-hydroxy-Δ5-C27-steroid dehydrogenase/isomerase
- Δ5-3β-ol:
-
3β-hydroxy-5-cholenoic acid
- Δ5-3β,7α-diol:
-
3β,7α-dihydroxy–5-cholenoic acid
- Δ5-3β,7α,12α-triol:
-
3β,7α,12α-trihydroxy-5-cholenoic acid
- ALT:
-
alanine aminotransferase
- AST:
-
aspartate aminotransferase
- CA:
-
cholic acid
- CDCA:
-
chenodeoxycholic acid
- Cr:
-
creatinine
- D Bil:
-
direct bilirubin
- GC-MS:
-
gas chromatography-mass spectrometry
- GGT:
-
γ-glutamyltransferase
- T Bil:
-
total bilirubin
- UDCA:
-
ursodeoxycholic acid
References
Clayton PT, Leonard JV, Lawson AM, Setchell KD, Andersson S, Egestad B, Sjövall J 1987 Familial giant cell hepatitis associated with synthesis of 3β,7α-dihydroxy- and 3β,7α,12α-trihydroxy-5-cholenoic acids. J Clin Invest 79: 1031–1038
Buchmann MS, Kvittingen EA, Nazer H, Gunasekaran T, Clayton PT, Sjövall J, Björkhen I 1990 Lack of 3β-hydroxyΔ5-C27-steroid dehydrogenase/isomerase in fibroblasts from a child with urinary excretion of 3β-hydroxy-Δ5-bile acids. J Clin Invest 86: 2034–2037
Schwarz M, Wright AC, Davis DL, Nazer H, Björkhem I, Russell DW 2000 The bile acid synthesis gene 3β-hydroxyΔ5-C27-steroid oxidoreductase is mutated in progressive intrahepatic cholestasis. J Clin Invest 106: 1175–1184
Cheng JB, Jacquemin E, Gerhardt M, Nazer H, Cresteil D, Heubi JE, Setchell KD, Russell DW 2003 Molecular genetics of 3β-hydroxyΔ5-C27-steroid oxidoreductase deficiency in 16 patients with loss of bile acid synthesis and liver disease. J Clin Endocrinol Metab 88: 1833–1841
Fischler B, Bodin K, Stjernman H, Olin M, Hansson M, Sjövall J, Björkhen I 2007 Cholestatic liver disease in adults may be due to an inherited defect in bile acid biosynthesis. J Intern Med 262: 254–262
Terazawa S, Kimura A, Inoue T, Murai T, Kurosawa T, Takao A 1998 An infant with 3β-hydroxyΔ5-C27-steroid dehydrogenase/isomerase deficiency presenting with typical neonatal hepatitis syndrome: the first Japanese case. Acta Paediatr Jpn 40: 638–640
Yamato Y, Kimura A, Murai T, Yoshimura T, Kurosawa T, Terazawa S, Takao A, Maeda K, Nakashima E, Yamashita Y, Kato H 2001 3β-HydroxyΔ5-C27-steroid dehydrogenase deficiency: diagnosis and treatment. J Paediatr Child Health 37: 516–519
Setchell KD, Schwarz M, O'Connell NC, Lund EG, Davis DL, Lathe R, Thompson HR, Weslie Tyson R, Sokol RJ, Russell DW 1998 Identification of a new inborn error in bile acid synthesis: mutation of the oxysterol 7alpha-hydroxylase gene causes severe neonatal liver disease. J Clin Invest 102: 1690–1703
Ueki I, Kimura A, Nishiyori A, Chen H-L, Takei H, Nittono H, Kurosawa T 2008 Neonatal cholestatic liver disease in an Asian patient with a homozygous mutation in the oxysterol 7α-hydroxylase gene. J Pediatr Gastroenterol Nutr 46: 465–469
Tohma M, Mahara R, Takeshita H, Kurosawa T 1986 A convenient synthesis of 3β,12α-, 3β,7α-, and 3β,7β-dihydroxy-5-cholen-24-oic acids: unusual bile acids in human biological fluids. Steroids 48: 331–338
Kimura A, Mahara R, Inoue T, Nomura Y, Murai T, Kurosawa T, Tohma M, Noguchi K, Hoshiyama A, Fujisawa T, Kato H 1999 Profile of urinary bile acids in infants and children: developmental pattern of excretion of unsaturated ketonic bile acids and 7β-hydroxylated bile acids. Pediatr Res 45: 603–609
Sundaram SS, Bove KE, Lovell MA, Schol RJ 2008 Mechanisms of disease: inborn errors of bile acid synthesis. Nat Clin Pract Gastroenterol Hepatol 5: 456–468
Kobayashi M, Koike M, Sakiyama M, Okuda S, Okuda M, Tanaka T, Unno A, Nittono H, Takei H, Murai T, Yoshimura T, Kurosawa T 2000 3β-Hydroxy-Δ5-C27 dehydrogenase/isomerase deficiency in a 23-year-old woman. Pediatr Int 42: 685–688
Gustafsson J 1986 Bile acid biosynthesis during development: hydroxylation of C27-sterols in human fetal liver. J Lipid Res 27: 801–806
Ishibashi S, Schwarz M, Frykman PK, Herz J, Russell DW 1996 Disruption of cholesterol 7α-hydroxylase gene in mice. I. Postnatal lethality reversed by bile acid and vitamin supplementation. J Biol Chem 271: 18017–18023
Schwarz M, Lund EG, Setchell KD, Kayden HJ, Zerwehk JE, Björkhem I, Herz J, Russell DW 1996 Disruption of cholesterol 7α-hydroxylase gene in mice. II. Bile acid deficiency is overcome by induction of oxysterol 7α-hydroxylase. J Biol Chem 271: 18024–18031
Ichimiya H, Nazer H, Gunasekaran T, Glayton P, Sjövall J 1990 Treatment of chronic liver disease caused by 3β-hydroxy-Δ5-C27 dehydrogenase deficiency with chenodeoxycholic acid. Arch Dis Child 65: 1121–1124
Ichimiya H, Egestad B, Nazer H, Baginski ES, Clyton PT, Sjövall J 1991 Bile acids and bile alcohols in a child with hepatic 3β-hydroxy-Δ5-C27 dehydrogenase deficiency: effect of chenodeoxycholic acid treatment. J Lipid Res 32: 829–841
Jacquemin E, Setchell KD, O'Connell NC, Estrada A, Maggiore G, Schmitz J, Hadchoael M, Bernard O 1994 A new cause of progressive intrahepatic cholestasis: 3β-hydroxy-C27 dehydrogenase/isomerase deficiency. J Pediatr 125: 379–384
Subramaniam P, Clayton PT, Portmann BC, Mieli-Vergani G, Hadžić N 2010 Variable clinical spectrum of the most common inborn error of bile acid metabolism-3β-hydroxy-Δ5-C27-steroid dehydrogenase deficiency. J Pediatr Gastroenterol Nutr 50: 61–66
Jacquemin E, Gerhardt M, Cresteil D, Fabre M, Taburet AM, Hadchouel M, Trivin F, Setchell KD, Bernard O 2001 Long-term effects of bile acid therapy in children with defects of primary bile acid synthesis: 3β-hydroxy-C27-steroid-dehydrogenase/isomerase and Δ4-3-oxosteroid 5β-reductase deficiencies. In: von Berge Henegouwen GP, Keppler D, Leuschner U, Paumgartner G, Stiehl A (eds) Biology of Bile Acids in Health and Disease. Kluwer Academic Publishers, Dordrecht, The Netherlands, pp 278–282
Gonzales E, Gerhardt MF, Fabre M, Setchell KD, Davit-Spraul A, Vincent I, Heubi JE, Bernard O, Jacquemin E 2009 Oral cholic acid for hereditary defects of primary bile acid synthesis: a safe and effective long-term therapy. Gastroenterology 137: 1310–1320
Makishima M, Okamoto AY, Repa JJ, Tu H, Learned MR, Luk A, Hull MV, Lustig KD, Mangelsdorf DJ, Shan B 1999 Identification of a nuclear receptor for bile acid. Science 284: 1362–1365
Parks DJ, Blanchard SG, Bledsoe RK, Chandra G, Consier TG, Kliewer SA, Stimmel JB, Willson TM, Zavacki AM, Moore DD, Lehmann JM 1999 Bile acids: natural ligands for an orphan nuclear receptor. Science 284: 1365–1368
Ellis E, Axelson M, Abrahamsson A, Eggertsen G, Thörne A, Nowak G, Ericzon B-G, Björkhem I, Einarsson C 2003 Feedback regulation of bile acid synthesis in primary human hepatocytes: evidence that CDCA is the strongest inhibitor. Hepatology 38: 930–938
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mizuochi, T., Kimura, A., Ueki, I. et al. Molecular Genetic and Bile Acid Profiles in Two Japanese Patients With 3β-Hydroxy-Δ5-C27-Steroid Dehydrogenase/Isomerase Deficiency. Pediatr Res 68, 258–263 (2010). https://doi.org/10.1203/PDR.0b013e3181eb0188
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/PDR.0b013e3181eb0188
This article is cited by
-
Bile Acid Synthesis Disorders in Japan: Long-Term Outcome and Chenodeoxycholic Acid Treatment
Digestive Diseases and Sciences (2021)
-
Two neonatal cholestasis patients with mutations in the SRD5B1 (AKR1D1) gene: diagnosis and bile acid profiles during chenodeoxycholic acid treatment
Journal of Inherited Metabolic Disease (2013)
