
Abstract
For studying the presence of programmed cell death in the lungs of infants with fatal respiratory distress syndrome (RDS) and the possible contribution of postnatal glucocorticoid administration on this cell destruction, lung tissue samples from autopsies of 16 premature infants with fatal RDS were studied. The infants had neither been exposed to antenatal steroids nor received surfactant therapy, but seven of these infants had been subjected to postnatal dexamethasone treatment. Lung autopsy samples of seven term and two preterm neonates without any obvious lung disease served as controls. Lungs were studied histologically, and apoptotic cell death was identified using DNA nick end-labeling assay and caspase-related M30 antibody staining (CytoDeath). Lung tissue from the RDS infants showed elevated leukocyte infiltration, histologic injury score, and number of apoptotic cells, located mainly in the respiratory epithelium, when compared with controls. In contrast, lungs from infants who had RDS and received dexamethasone demonstrated markedly reduced tissue leukocyte accumulation and injury score and lower rates of epithelial apoptosis than the lungs of infants who had RDS and did not receive dexamethasone. These results suggest that significant epithelial apoptosis is present in the lungs of newborn infants with fatal RDS and that this apoptosis may be attenuated by steroid administration.
Similar content being viewed by others


Main
Respiratory distress syndrome (RDS) is a severe pulmonary disease with high mortality and morbidity in premature infants. The pathogenesis of this lung disorder is associated with lung immaturity and insufficient surfactant biosynthesis. However, lung inflammation has also been suggested to contribute to respiratory failure in infants with RDS (1, 2). Therapy for RDS with mechanical ventilation and hyperoxia might worsen the lung injury and contribute to respiratory failure in patients with RDS by activating proinflammatory cells and directly injuring the alveolar epithelium or endothelium. Pharmacologic therapy with glucocorticoids and surfactant have reduced but not eliminated morbidity and mortality from RDS (1–5).
Apoptosis, a noninflammatory programmed cell death, has recently been observed in tissue injuries of various organs, including the lungs (6–9). Although generally regarded as a cell clearance mechanism promoting resolution of inflammation (10), apoptosis triggered by diverse extrinsic or intrinsic signals, such as oxidative stress, endotoxin, or bleomycin, may contribute to the pathophysiology of the acute injury in adult lungs (6, 9, 11, 12). Experimental studies in our laboratory have recently described significant epithelial apoptosis in meconium-contaminated neonatal lungs (13), but the occurrence of pulmonary apoptosis in human neonatal RDS is still unexplored. We thus decided to investigate the presence of apoptotic cell death in lung tissue of infants with fatal RDS using the terminal transferase mediated DNA nick end-labeling assay (TUNEL) and the caspase-related M30 antibody-staining (CytoDeath, Boehringer Mannheim, Mannheim, Germany).
METHODS
Patients.
The patient register of the neonatal intensive care unit of the Department of Pediatrics, University of Turku, from a 10-year period (1986-1996) was reviewed for deaths caused by severe RDS without any clinical evidence of infection. Because postnatal dexamethasone and surfactant therapy have been standard treatment procedures for RDS infants in the local neonatal intensive care unit since 1992, lung tissue samples were selected from autopsy material obtained between the years 1986 and 1991. From this material, tissue samples of consecutive infants with severe RDS and no ante- or postnatal steroid or surfactant treatment (n = 9) were collected for analysis (Table 1). From the same time period, lung tissue samples of another group of consecutive infants with fatal RDS but with a postnatal course of i.v. dexamethasone (n = 7) were similarly analyzed. Five lung autopsy samples from infants with no evidence of lung disease or steroid therapy served as controls (Table 1). All infants were mechanically ventilated, except one control infant (patient 18). The severity of RDS was assessed by a scoring system (0 = no, 1 = mild, 2 = moderate, and 3 = severe RDS) based on five clinical items (Table 1) (14). Time from death to autopsy could not be defined exactly in every instance but varied between 1 and 3 d. Samples with obvious cytolysis were excluded. The study was approved by the local Ethics Committee, and informed consent was obtained from the parents.
Histologic examination.
Pulmonary tissue samples from the autopsies, fixed in 10% buffered formalin, embedded in paraffin, and stained with hematoxylin and eosin, were studied to determine the severity of the lung injury. Samples were assessed by a pathologist blinded to the grouping of the infants. A score from 0 to 4 represented the percentile of affected area of the lung section (0 = 0%; 1 = 1%-25%; 2 = 26%-50%; 3 = 51%-75%; 4 = 76%-100%) and was assigned for three different characteristics: 1) extension of pulmonary leukocyte infiltration, 2) amount of intra-alveolar leukocytes, and 3) amount of exudative debris and edema fluid. The calculated total injury score represents the sum of these scores (15).
In situ detection of apoptotic cells.
In situ detection of apoptotic cells in paraffin-embedded 3-μm-thick sections was performed as described earlier (8), with slight modifications. Briefly, deparaffinized sections were treated with 10 μg/mL proteinase K (Boehringer, Mannheim, Germany) at 37°C in 2 mM CaCl2 and 20 mM Tris-HCL (pH 7.4) for 30 min. The endogenous peroxidase activity was blocked by incubating the slides in 0.3% hydrogen peroxide in water for 10 min at 25°C. DNA 3′-end labeling was performed after 10 min of incubation with terminal transferase buffer (Promega, Madison, WI, U.S.A.). The labeling mixture contained fresh terminal transferase buffer, 5 μM nonradioactive digoxigenin-dideoxy-UTP (dig-ddUTP; Boehringer), 45 μM ddATP (Pharmacia, Uppsala, Sweden), and 0.34 U/μL terminal transferase (Promega). The reaction was allowed to continue for 1 h at 37°C in a humidified chamber. After washing, the slides were incubated with blocking buffer containing 2% (wt/vol) blocking reagent and 0.05% (wt/vol) sodium azide (Boehringer) for 30 min. Antidigoxigenin antibody, conjugated to alkaline phosphatase (1:3000, Boehringer), in 2% (wt/vol) blocking buffer was added and incubated for 2 h in a humidified chamber. The slides were treated with alkaline phosphatase buffer (0.1 M NaCl, 0.05 M MgCl, and 0.1 M Tris-HCl [pH 9.5]) for 10 min. Thereafter, 337 μg/mL nitroblue tetrazolium salt (Boehringer) and 175 μg/mL 5-bromo-4-chloro-3-inodyl-phosphate (Boehringer) were added in fresh alkaline phosphatase buffer, and the reaction was terminated 3 h and 45 min later by several washes with 1 mM EDTA and 10 mM Tris-HCl (pH 8.0). Finally, the slides were mounted with Gurr Aquamount (BDH Chemicals, Poole, England). For controls, terminal transferase, dig-ddUTP, or antidigoxigenin antibody was omitted from reaction. Staining of lymphocytes undergoing apoptosis in the lymph nodes served as a positive control.
Apoptotic cells were counted in lung sections stained with the antidigoxigenin antibody. A distinct color reaction within the cells was regarded to represent apoptotic DNA fragmentation. The results are expressed as the number of positive cells per mm2 of tissue section area in at least 10 fields of view of a 10× objective lens. The in situ detection of free DNA 3′ ends is a well-established method in the detection of apoptotic cellular changes and has been validated by simultaneous electrophoretic DNA analysis in pancreatic tissue (8).
Identification of apoptosis in the epithelial cells.
For identifying the occurrence of apoptosis in the pulmonary epithelium, thin (3 μm) successive sections of paraffin-embedded lung tissue samples were studied. The first section was pretreated with pepsin and stained with a monoclonal mouse antibody MNF-116 (Dakopatts, Glostrup, Denmark), which recognizes type II epithelial cells (16, 17). The sections were blocked with Tris-buffered saline (TBS) containing 1 mg/mL BSA (TBS/BSA) for 30 min and incubated with the antibody, diluted 1:10 in TBS, for 30 min. After incubation, the sections were washed three times with TBS and stained with the secondary antibody. The consecutive section was stained by the TUNEL-method for the detection of the DNA-fragmentation, as described earlier. Apoptosis was identified in the epithelium by comparing the staining in both sections.
For further confirming apoptotic changes in the respiratory epithelium, separate sections of the lungs were stained with a caspase-related monoclonal antibody M30 (CytoDeath). Formalin-fixed, paraffin-embedded sections were prepared and processed according to the protocol described earlier (18). Briefly, deparaffinized sections were pretreated by microwave irradiation at 350 W for 5 min in citrate buffer (0.01 M citrate buffer at pH 6.0). Sections were incubated with M30 antibody at 1 μg IgG/mL overnight at room temperature. Sections were incubated with biotinylated rabbit anti-mouse Ig (DAKO A/S, Glostrup, Denmark) followed by streptavidin-HRP (Roche Molecular Biochemicals). Sections were visualized with diaminobenzidine and counterstained with hematoxylin. M30 is a new antibody that detects caspase-cleaved cytokeratin 18, and it has been regarded to be specific for apoptotic epithelial cells (18). For quantitative analysis, the number of M30-positive cells was counted in at least 10 fields of view of a 10× objective lens.
Statistical analysis.
The results are expressed as means (SD). One-way ANOVA was used to compare the number of apoptotic cells or lung injury scores in lung tissue between the groups. When the overall ANOVA was significant, comparison between the groups was made using Newman-Keuls post hoc test. Pearson analysis was used to evaluate further the correlation between apoptosis and lung injury parameters or gestational age. A level of p < 0.05 was considered statistically significant.
RESULTS
The clinical data of the study infants are shown in Table 1. The gestational age, birth weight, and Apgar scores of the RDS infants with or without dexamethasone treatment were similar, whereas the control infants were more mature and had higher birth weight.
Histologic analysis of the RDS lungs showed significantly higher level of leukocyte infiltration, alveolar exudate, and total injury score compared with controls (Table 2). In contrast, leukocyte infiltration and total injury score in the dexamethasone-exposed RDS lungs were significantly lower than in the nonexposed lungs, but the amount of alveolar exudate was similar (Table 2).
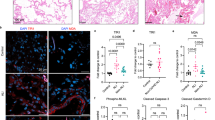
In the autopsy lung samples, apoptosis was detected mainly (>90%) in the epithelial cells, confirmed with TUNEL, MNF-116, and M30 stainings (Figs. 1 and 2). The number of apoptotic epithelial cells was significantly (p < 0.01) higher in the lungs of infants with RDS, when compared with controls (Figs. 3 and 4). In contrast, lung samples of infants with RDS and dexamethasone treatment showed a significantly (p < 0.01) lower number of TUNEL-positive cells than those of the nontreated infants with RDS, being similar to the finding in the control group (Figs. 3 and 4). Because the control infants in our study were more mature than the infants with RDS, we also studied the lungs of two preterm infants without RDS for pulmonary tissue apoptosis. The first infant was born at 24 wk of gestation and died of immaturity at 5 h of age, and the second infant was born at 26 wk of gestation and died of intraventricular hemorrhage at 2 d of age. Neither of these infants had significant respiratory disorder, and they had not received steroid or surfactant therapy. The lungs of both infants showed an apoptotic rate that was in the range of the mature controls (32 and 8 apoptotic cells/mm2 versus 16.3 ± 9.7 apoptotic cells/mm2 in the term controls). The apoptotic incidence did not correlate with the gestational age, age at death, birth weight, leukocyte infiltration, alveolar exudate, or total injury score in any of the study groups.

Apoptotic epithelial cells in the lungs of infant with RDS. Dark staining represents DNA fragmentation in TUNEL-stained lung section (A1 and A2). Consecutive section of the same lung stained with a specific epithelial marker MNF-116 (B1 and B2). Apoptosis was identified in the epithelium by comparing the staining in both sections. Magnification: A1 and B1, ×260; A2 and B2, ×780.

Lung tissue section of infant with RDS. Brown staining represents early apoptotic changes in lung epithelial cells stained with M30 antibody, an indicator of caspase activation. Magnification: ×780.

Number of apoptotic cells in lung tissue as indicated by TUNEL staining. Control, infants with no lung injury; RDS, infants with respiratory distress syndrome; RDS/D, infants with RDS and dexamethasone therapy. Results are expressed as means (SD). *p < 0.01 vs control and RDS/D group.

TUNEL-positive cells in lung tissue. (A) Control. (B) Infant with respiratory distress syndrome (RDS). (C) Infant with RDS and dexamethasone therapy. Dark staining represents DNA fragmentation. Magnification: ×260.
DISCUSSION
A characteristic feature of the acute lung injury in adults and neonates is widespread destruction of the alveolar epithelium leading to pulmonary edema formation and gas exchange abnormalities with impairment of the clinical outcome (19, 20). Although necrotic injury to the airway epithelium with a significant inflammatory component is widely recognized in neonatal distressed lungs, especially when not treated with surfactant (19), anti-inflammatory treatment strategies have been unsuccessful (1, 4, 5). Recent experimental evidence, however, suggests that epithelial apoptotic cell death could play a role in the pathogenesis of acute injury in adult and supposedly also in neonatal lungs (6, 7, 13, 21). Our present data demonstrate significant epithelial apoptosis in the lungs of newborn infants with fatal RDS. This programmed cell death may potentiate the detrimental effects of the inflammatory necrotic injury and surfactant insufficiency on immature pulmonary function and is thus proposed to be involved in the pathogenesis of neonatal RDS.
The mechanisms of the pulmonary apoptosis remain unclear, although previous experimental data have indicated oxidative stress, endotoxin, and cytokines as mediators of apoptosis in acute lung injury (6, 9, 22, 23). The inflammatory response with cytokine surge in the lungs and therapeutic use of high inspiratory concentrations of oxygen and mechanical ventilation thus may predispose the lungs of the distressed infants to epithelial apoptosis. In addition, physiologic fetal development and birth itself may be associated with increased pulmonary apoptosis (24, 25), and postmortal changes in lung tissue samples may interfere, but these likely do not explain our present findings, especially because postnatal dexamethasone treatment was associated with diminished apoptosis in the severely distressed lungs. Furthermore, because of the different gestational ages of the infants with RDS and the controls in our study, we cannot totally exclude some gestation-related effects on apoptosis, but the low number of apoptotic cells in the two premature control lungs renders it unlikely. Earlier human and experimental data similarly indicate no correlation between the amount of pulmonary apoptosis and gestational age (24, 25).
Glucocorticoids have powerful cell membrane-stabilizing, anti-inflammatory, and maturational effects on the lungs (22, 26). Accordingly, antenatal and early postnatal administration of steroids may diminish airway inflammatory reaction and edema formation in preterm infants with RDS (1, 4). This pulmonary effect of steroids was also apparent in our study. There is also evidence indicating an increase in the lung surfactant protein synthesis in preterm infants with RDS after early postnatal dexamethasone treatment (27, 28). Furthermore, corroborating our recent experimental findings in meconium-contaminated neonatal lungs (29), the present study indicates that early dexamethasone therapy is associated with prevention of the lung epithelial apoptosis in RDS. The inhibition of apoptotic cell death in the respiratory epithelium may contribute to diminished pulmonary damage and may possibly also improve pneumocyte surfactant production in the immature distressed lungs. Although the apoptosis-inhibiting mechanisms of dexamethasone still remain unclear and its multiple effects on the neonatal lung cannot be evaluated from the present data, previous in vitro studies with cultured lung epithelial cells suggest a steroid-related blockage of caspases (22). On the basis of our observations, inhibition of lung epithelial apoptosis thus could be considered as one of the potent effects by which steroids modulate the pulmonary function and the clinical course of RDS in premature infants. In conclusion, our results demonstrate significant epithelial apoptosis in lung tissue from newborn infants with RDS and suggest that postnatal dexamethasone treatment may attenuate apoptotic death in the lung epithelium. These data suggest that alveolar epithelial apoptosis may be important in the pathogenesis of RDS.
Abbreviations
- RDS:
-
respiratory distress syndrome
- TUNEL:
-
terminal deoxynucleotidyl transferase-mediated nick end-labeling
REFERENCES
Jobe AH, Ikegami M 1998 Mechanisms initiating lung injury in the preterm. Early Hum Dev 53: 81–94.
Brus F, van Oeveren W, Okken A, Bambang SO 1996 Activation of circulating polymorphonuclear leukocytes in preterm infants with severe idiopathic respiratory distress syndrome. Pediatr Res 39: 456–463.
Jones CA, Cayabyab RG, Kwong KY, Stotts C, Wong B, Hamdan H, Minoo P, deLemos RA 1996 Undetectable interleukin (IL)-10 and persistent IL-8 expression early in hyaline membrane disease: a possible developmental basis for the predisposition to chronic lung inflammation in preterm newborns. Pediatr Res 39: 966–975.
Wang JY, Yeh TF, Lin YJ, Chen WY, Lin CH 1997 Early postnatal dexamethasone therapy may lessen lung inflammation in premature infants with respiratory distress syndrome on mechanical ventilation. Pediatr Pulmonol 23: 193–197.
Sanders RJ, Cox C, Phelps DL, Sinkin RA 1994 Two doses of early intravenous dexamethasone for the prevention of bronchopulmonary dysplasia in babies with respiratory distress syndrome. Pediatr Res 36: 122–128.
Kawasaki M, Kuwano K, Hagimoto N, Matsuba T, Kunitake R, Tanaka T, Maeyama T, Hara N 2000 Protection from lethal apoptosis in lipopolysaccharide-induced acute lung injury in mice by a caspase inhibitor. Am J Pathol 157: 597–603.
Thompson CB 1995 Apoptosis in the pathogenesis and treatment of disease. Science 267: 1456–1462.
Laine VJO, Nyman KM, Peuravuori HJ, Henriksen K, Parvinen M, Nevalainen TJ 1996 Lipopolysaccharide induced apoptosis of rat pancreatic acinar cells. Gut 38: 747–752.
Kazzaz JA, Xu J, Palaia TA, Mantell L, Fein AM, Horowitz S 1996 Cellular oxygen toxicity. Oxidant injury without apoptosis. J Biol Chem 271: 15182–15186.
Bardales RH, Xie SS, Schaefer RF, Hsu SM 1996 Apoptosis is a major pathway responsible for the resolution of type II pneumocytes in acute lung injury. Am J Pathol 149: 845–852.
Fine A, Janssen-Heininger Y, Soultanakis RP, Swisher SG, Uhal BD 2000 Apoptosis in lung pathophysiology. Am J Physiol 279:L423–L427.
Wang R, Ibarra-Sunga O, Verlinski L, Pick R, Uhal BD 2000 Abrogation of bleomycin-induced epithelial apoptosis and lung fibrosis by captopril or by a caspase inhibitor. Am J Physiol 279:L143–L151.
Holopainen R, Aho H, Laine J, Peuravuori H, Soukka H, Kaapa P 1999 Human meconium has high phospholipase A2 activity and induces cellular injury and apoptosis in piglet lungs. Pediatr Res 46: 626–632.
Korvenranta H, Kero P, Valimaki I 1983 Cardiovascular monitoring in infants with respiratory distress syndrome. Biol Neonate 44: 138–145.
Holopainen R, Aho H, Laine J, Halkola L, Kaapa P 1999 Nitric oxide inhalation inhibits pulmonary apoptosis but not inflammatory injury in porcine meconium aspiration. Acta Paediatr 88: 1147–1155.
Schittny JC, Djonov V, Fine A, Burri PH 1998 Programmed cell death contributes to postnatal lung development. Am J Respir Cell Mol Biol 18: 786–793.
Kasper M, Rudolf T, Verhofstad AA, Schuh D, Muller M 1993 Heterogeneity in the immunolocalization of cytokeratin-specific monoclonal antibodies in the rat lung: evaluation of three different alveolar epithelial cell types. Histochemistry 100: 65–71.
Leers MP, Kolgen W, Bjorklund V, Bergman T, Tribbick G, Persson B, Bjorklund P, Ramaekers FC, Bjorklund B, Nap M, Jornvall H, Schutte B 1999 Immunocytochemical detection and mapping of a cytokeratin 18 neo-epitope exposed during early apoptosis. J Pathol 187: 567–572.
Pinar H, Makarova N, Rubin LP, Singer DB 1994 Pathology of the lung in surfactant-treated neonates. Pediatr Pathol 14: 627–636.
Ware LB, Matthay MA 2000 The acute respiratory distress syndrome. N Engl J Med 342: 1334–1349.
Matute-Bello G, Liles WC, Steinberg KP, Kiener PA, Mongovin S, Chi EY, Jonas M, Martin TR 1999 Soluble Fas ligand induces epithelial cell apoptosis in humans with acute lung injury (ARDS). J Immunol 163: 2217–2225.
Wen LP, Madani K, Fahrni JA, Duncan SR, Rosen GD 1997 Dexamethasone inhibits lung epithelial cell apoptosis induced by IFN-gamma and Fas. Am J Physiol 273:L921–L929.
Buckley S, Barsky L, Driscoll B, Weinberg K, Anderson KD, Warburton D 1998 Apoptosis and DNA damage in type 2 alveolar epithelial cells cultured from hyperoxic rats. Am J Physiol 274:L714–L720.
Kresch MJ, Christian C, Wu F, Hussain N 1998 Ontogeny of apoptosis during lung development. Pediatr Res 43: 426–431.
Scavo LM, Ertsey R, Chapin CJ, Allen L, Kitterman JA 1998 Apoptosis in the development of rat and human fetal lungs. Am J Respir Cell Mol Biol 18: 21–31.
Jobe AH, Ikegami M 2000 Lung development and function in preterm infants in the surfactant treatment era. Annu Rev Physiol 62: 825–846.
Wang JY, Yeh TF, Lin YC, Miyamura K, Holmskov U, Reid KB 1996 Measurement of pulmonary status and surfactant protein levels during dexamethasone treatment of neonatal respiratory distress syndrome. Thorax 51: 907–913.
Ogasawara Y, Kuroki Y, Tsuzuki A, Ueda S, Misaki H, Akino T 1992 Pre- and postnatal stimulation of pulmonary surfactant protein D by in vivo dexamethasone treatment of rats. Life Sci 50: 1761–1767.
Holopainen R, Laine J, Halkola L, Aho H, Kaapa P 2001 Dexamethasone treatment attenuates pulmonary injury in piglet meconium aspiration. Pediatr Res 49: 162–168.
Acknowledgements
We thank Sinikka Kollanus for technical assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
This study is financially supported by the Turku University Hospital, the Sigrid Juselius Foundation, and the Pediatric Research Foundation, Finland.
Rights and permissions
About this article
Cite this article
Lukkarinen, H., Laine, J. & Kääpä, P. Lung Epithelial Cells Undergo Apoptosis in Neonatal Respiratory Distress Syndrome. Pediatr Res 53, 254–259 (2003). https://doi.org/10.1203/01.PDR.0000047522.71355.08
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/01.PDR.0000047522.71355.08
