
Abstract
Neonates have a lower serum bicarbonate level than adults, which is caused by a lower renal threshold for bicarbonate. Eighty percent of bicarbonate reabsorption occurs in the proximal tubule, in which proton secretion is predominantly mediated by a luminal Na+/H+ antiporter. Previous studies have demonstrated that there is a maturational increase in apical membrane rabbit proximal convoluted tubule Na+/H+ antiporter activity. However, in rat brush border membrane vesicles, Na+/H+ activity was higher in neonates than that in adult rats. To examine the maturation of Na+/H+ antiporter activity in rat proximal convoluted tubules, we perfused rat proximal convoluted tubules in vitro. Na+/H+ antiporter activity was assayed as the proton secretory rate on luminal sodium removal. Na+/H+ antiporter activity was 121.2 ± 18.4 pmol/mm·min in neonatal and 451.8 ± 40.6 pmol/mm·min in adult proximal convoluted tubules (p < 0.001). We next examined whether the increase in Na+/H+ antiporter activity was associated with changes in renal cortical NHE3 mRNA and brush border membrane NHE3 protein abundance. Adult renal cortical NHE3 mRNA abundance was 10-fold greater than that in 1-d-old neonates (p < 0.001). There was a comparable developmental increase in renal brush border membrane vesicle NHE3 protein abundance (p < 0.001). In summary, this study demonstrates that there is a maturational increase in rat apical membrane Na+/H+ antiporter activity, renal cortical NHE3 mRNA, and brush border membrane vesicle NHE3 protein abundance.
Similar content being viewed by others
Main
The proximal tubule reabsorbs 80% of filtered bicarbonate. Luminal proton secretion in this segment is mediated predominantly by an apical membrane Na+/H+ antiporter and to a lesser extent by an H+-ATPase (1, 2). Although a number of isoforms for the Na+/H+ exchanger have been cloned, there is compelling evidence that NHE3 mediates most luminal proton secretion in this segment (3–8).
The lower serum bicarbonate level in neonates is in large part the result of an immature proximal tubule with lower rates of bicarbonate reabsorption compared with adults (9, 10). Most of luminal proton secretion by the proximal tubule is mediated by the luminal Na+/H+ antiporter. Thus, the maturational changes that occur in proximal tubule Na+/H+ antiporter activity are key to our understanding of postnatal renal acidification (11). In addition to mediating proton secretion for bicarbonate reclamation, the Na+/H+ antiporter, in parallel with Cl−/base exchange, mediates active NaCl transport in this segment (12, 13). There is also evidence that the developmental increase in proximal tubule Na+/H+ antiporter activity is responsible for inducing the postnatal increase in basolateral Na+-K+-ATPase activity (14).
We have previously demonstrated that there is a maturational increase in rabbit PCT Na+/H+ antiporter activity (2, 15), and others have confirmed this using rabbit BBMV (16). However, Na+/H+ antiporter activity in rat renal BBMV was found to be higher in neonates than in adults (17), raising the possibility that our findings (2, 15) and those of Beck et al. (16) may be peculiar to the rabbit. The purpose of the present study was to determine Na+/H+ antiporter activity in intact rat proximal tubules. To this end, we perfused rat PCTs in vitro to measure apical membrane Na+/H+ antiporter activity. In addition, we compared Na+/H+ antiporter activity to the maturational changes in renal cortical NHE3 mRNA and BBMV NHE3 protein abundance.
METHODS
In vitro microperfusion.
We have previously tried to dissect tubules from 3- to 6-mo-old rats without success. The dense connective tissue does not permit freehand dissection of PCTs. We have also tried to perfuse proximal tubules from rats after incubation with collagenase. Although the tubules were easily dissected, the tubules fragmented after placement in the microperfusion pipettes. However, 0.2- to 0.5-mm tubules from 14- to 40-d-old Sprague-Dawley rats can be dissected freehand without collagenase and perfused in vitro.
Isolated segments of neonatal and adult rat PCTs were perfused by concentric glass pipettes using techniques previously described for rabbit proximal tubules (2, 13, 15). Briefly, neonatal (13–15 d of age) and adult (33–37 d of age) tubules were dissected in Hanks' balanced salt solution containing (in mM) 137 NaCl, 5 KCl, 0.8 MgSO4, 0.33 Na2HPO4, 0.44 KH2PO4, 1 MgCl2, 10 Tris, 0.25 CaCl2, 2 glutamine, 2 heptanoic acid, and 2 lactate at 4°C. Tubules were transferred to a 0.2-mL chamber in which the bathing solution was preheated to 38°C.
Measurement of pHi.
The solutions used in these experiments are shown in Table 1. The fluorescent dye BCECF was used to measure pHi as described previously (2, 13, 15, 18). pHi was measured using a Nikon inverted epifluorescent microscope (Nikon, Japan) attached to a PTI Ratiomaster (Photon Technology International, South Brunswick, New Jersey, U.S.A.) at a rate of 30 measurements per second. A variable diaphragm was placed over the area to be measured. To calculate pH from the ratio of fluorescence (F500/F450), a nigericin calibration curve was performed as previously described (15, 18, 19). There was a small difference in the calibration curves of adult and neonatal PCTs. Calibrations performed in tubules with EIPA had a significantly different intercept than respective tubules without EIPA. All rates and pHi were calculated with their respective nigericin calibration curves.
Measurement of JH.
Proton flux rates (JH, in pmol/mm·min) resulting from a luminal fluid change were calculated using the following FORMULA : where d pHi/ dt is the rate of initial change in pHi after a luminal fluid change, V is the tubular volume in liters, and β is the buffer capacity.

Tubules were incubated with an ultrafiltrate-like solution in the lumen and bath (solution B containing 5 mM glucose and 5 mM alanine) for at least 5 min before loading with 5 × 10−6 M BCECF, and tubules had a constant pHi for several minutes before the measurement of the transporter activity. d pHi/ dt was measured from the slope of the change in pHi immediately after a luminal fluid change. Steady-state pHi values were reached within 1 min after a luminal fluid exchange, but pHi was measured for several minutes to ensure a steady-state pHi was achieved.
Apparent buffer capacity (β) was measured as previously described using NH3/NH4+ (15, 20). Solutions (D and E) were used for measurement of apparent buffer capacity. These solutions did not contain Na+ or Cl− so as to inhibit all acidification mechanisms owing to Na+- and Cl−-dependent transporters. In the presence of 25 mM HCO3−, buffer capacity was 98.7 ± 16.4 mM/pH in neonatal PCT and 98.9 ± 17.6 mM/pH in adult PCT (p = NS). Tubular volume was calculated from the measured inner and outer tubular diameters at ×400 magnification using an eyepiece reticule. Tubular volume was 453 ± 42 × 10−12 L/mm in neonatal PCT and 1073 ± 45 × 10−12 L/mm in adult PCT (p < 0.001).
Na+/H+ antiporter activity was measured as previously described in adult rat PCTs perfused in vivo and neonatal and adult rabbit PCTs perfused in vitro (15, 18, 20). Neonatal and adult rat tubules were perfused at a rate of ∼2 μL/min with an ultrafiltrate-like solution without glucose and amino acids (solution B). Organic solutes were omitted from the luminal solution because sodium-coupled glucose and amino acid transport depolarizes the basolateral membrane, which may affect bicarbonate exit, an electrogenic process (18). 4-Acetamido-4′-isothiocyano-stilbene-2,2′ disulfonic acid (SITS, 1 mM) was present in the bathing solution to inhibit the sodium bicarbonate cotransporter, a major regulator of intracellular pHi in PCTs (15, 18). The bathing solution had a bicarbonate concentration of 5 mM and a pH of 6.6 to compensate for the cell alkalinization caused by the addition of bath SITS (16, 18). The bathing solution was exchanged at a rate of 5 mL/min. Under these conditions, changes in pHi in response to a change in luminal sodium concentration are a measure of Na+/H+ antiporter activity (15, 18, 20). In the experimental period, luminal sodium was removed (solution C).
BBMV isolation.
Neonatal rat kidneys were rapidly removed and placed in an ice-cold isolation buffer containing 300 mM mannitol, 16 mM HEPES, and 5 mM EGTA, titrated to pH 7.4 with Tris. The isolation buffer contained aprotinin (2 μg/mL), leupeptin (2 μg/mL), and phenylmethylsulfonyl fluoride (100 μg/mL). The cortex was dissected and then homogenized with 20 strokes with a polytetrafluoroethylene-glass homogenizer at 4°C. BBMV were then isolated by differential centrifugation and magnesium precipitation as previously described (21). The final BBMV fraction was resuspended in isolation buffer. Protein was assayed using the bicinchoninic acid assay (BCA Protein Assay, Pierce Chemical Company) using BSA as the standard.
SDS-PAGE and immunoblotting.
BBMV protein (50 μg/lane) was denatured and separated on a 7.5% polyacrylamide gel as previously described (21, 22). The proteins were transferred to polyvinylidene difluoride membrane overnight at 140 mA at 4°C. The blot was blocked with fresh Blotto (5% nonfat milk, 0.1% Tween 20, and PBS, pH 7.4) for 1 h, and then a primary antibody to NHE3 was added at a 1:250 dilution and incubated for 1 h at room temperature. The blot was washed with Blotto, and then the secondary horseradish peroxidase-conjugated donkey anti-rabbit immunoglobulin was added for 1 h at 1/10,000 dilution. The blot was washed with Blotto, and enhanced chemiluminescence was used to detect bound antibody (Amersham Life Science). The NHE3 antibody was a polyclonal rabbit anti-rat antifusion protein antiserum against a fusion protein of maltose-binding protein and rat NHE3 amino acids 405–831 (23). NHE3 protein abundance was quantitated using densitometry.
RNA isolation and analysis.
The kidneys were decapsulated, and renal cortex was homogenized in RNAzol [1:1, phenol-RNAzol stock (4 M guanidinium thiocyanate, 25 mM disodium citrate, pH 7.0), 0.5% sarcosyl] containing 3.6 μL/mL β-mercaptoethanol. RNA was extracted using 3 M NaOAc (pH 4.0) and chloroform, purified using isopropanol precipitation, and washed twice with 75% ethanol (24). Poly(A)+ RNA was purified using oligo (dT) column chromatography. Five micrograms of poly(A)+ RNA was fractionated by agarose-formaldehyde gel electrophoresis and transferred to a nylon filter (GeneScreen Plus; New England Nuclear, Boston, MA, U.S.A.). The filter was prehybridized at 42°C for 4 h with 5× standard saline citrate (SSC), 5× Denhardt's (Ficoll, BSA, and polyvinylpyrrolidone, each at 1 mg/mL), 0.5% SDS, and 0.5 mg/mL of sheared salmon sperm DNA, then hybridized to double-stranded uniformly 32P-labeled cDNA probes (>106 counts·min−1·mL−1) in the above hybridization solution at 42°C for 16 h. The probes were synthesized by the random hexamer method using 50–100 ng of DNA. NHE3 cDNA was the rat 1.2-kb Pst I fragment (25). The filter was then washed twice with 2× SSC and 0.1% SDS for 5 min at room temperature and then with 0.1× SSC and 1% SDS at 55°C for 40 min one or two additional times. Message abundance was quantitated by autoradiography and densitometry.
Institutional approval.
These studies were approved by the Animal Resources Center of the University of Texas Southwestern Medical Center.
Statistical analysis.
Statistical significance was determined by t test and ANOVA.
RESULTS
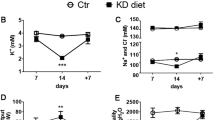
The steady-state pHi in neonatal and adult PCTs in the presence and absence of luminal sodium is shown in Table 2. The cell pHi was lower in the absence of luminal sodium than that in the control and recovery groups (p < 0.001). The rate of change in pHi with luminal sodium removal is shown in Figure 1. As can be seen, the d pHi/ dt was 1.5-fold and JH was 4-fold greater in adult compared with neonatal rat PCTs. As is shown in Figure 1, neonatal and adult Na+/H+ antiporter activity was inhibited by 100 μM EIPA.

Na+/H+ antiporter activity in rat PCTs perfused in vitro. Top, d pHi/ dt and Bottom, JH measured after removal of luminal sodium. Na+/H+ antiporter activity in neonatal and adult PCT was inhibited by luminal EIPA.
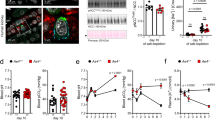
NHE3 is the predominant or only isoform of the Na+/H+ antiporter responsible for luminal proton secretion in the PCT (3–8). We next examined the ontogeny of rat renal cortical NHE3 mRNA. There was a significant increase in NHE3 mRNA abundance with postnatal maturation (p < 0.001). A typical Northern blot is shown in Figure 2. Renal cortical NHE3 mRNA abundance increased 10-fold during postnatal development. NHE3 mRNA abundance increased approximately 2.5-fold from d 14 to 24 (p < 0.05). There was no significant difference in renal cortical NHE3 mRNA abundance between 24-d-old and adult rats.

Maturational changes in NHE3 mRNA abundance. Northern blot with 5 μg of poly(A)+ RNA from renal cortex of rats of ages shown below lanes. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA is shown for comparison. There was a significant increase in NHE3 mRNA abundance as a function of age (p < 0.001).
We next examined the developmental increase in BBMV NHE3 protein abundance. As seen in Figure 3, there was a maturational increase in NHE3 protein abundance (p < 0.001). NHE3 protein was almost undetectable for the first week of life. There was a significant difference in BBMV NHE3 protein abundance between 1-, 7-, and 14-d-old rats and 24-d-old and adult rats. BBMV NHE3 protein abundance increased 7-fold between 14 and 24 d of age. BBMV NHE3 protein abundance was the same in 24-d-old and adult rats. These findings are consistent with our transport data indicating that there is a maturational increase in Na+/H+ antiporter activity in the rat.

Maturational changes in NHE3 protein abundance compared with β-actin. Renal BBMV were harvested from animals of ages shown below lanes. There was a maturational increase in NHE3 mRNA abundance (p < 0.001).
DISCUSSION
Na+/H+ antiporter activity was assayed as the rate of change in intracellular pH in response to a change in luminal sodium concentration using the same assay as has been used in adult rat proximal tubules in vivo (20). This was done so that a direct comparison could be made between the rates obtained in in vitro rat perfused tubules and those found in vivo. The d pHi/ dt was 3.70 ± 0.31 in vivo compared with 4.27 ± 0.36 pH units/min in the present study. These values were not different and confirm the viability of isolated perfused rat proximal tubules in vitro. The sodium-dependent proton secretion was inhibited by luminal EIPA consistent with a luminal Na+/H+ antiporter. Surprisingly, there was a significant increase in pHi in neonatal tubules perfused with EIPA and a tendency for a cell alkalinization in adult tubules as well. Inhibition of the Na+/H+ antiporter would be expected to result in cell acidification. The reason for the increase in pHi is unknown.
The serum bicarbonate level in neonates is lower than adults, which has been the topic of a review by our laboratory (11). Inasmuch as most of filtered bicarbonate is reabsorbed by the proximal tubule, the threshold for bicarbonate is in large part determined by the rate of bicarbonate reabsorption in this segment. The rate of bicarbonate reabsorption by the neonatal rabbit PCT is less than that in the adult segment (9, 10). Although proximal tubule bicarbonate reabsorptive rates have not been measured in the neonatal rat, one previous developmental study had examined the rate of volume absorption in rat superficial proximal tubules using free-flow micropuncture. The rate of volume absorption increased 2-fold from 22- to 24-d-old rats to 40- to 45-d-old adult rats (26). Although this study compared weanling rats to adults and did not measure bicarbonate reabsorption, this study is consistent with our findings and previous developmental studies in rabbits demonstrating that there is a maturational increase in proximal tubule transport (9).
BBMV have been used to study the maturation of the Na+/H+ antiporter in the rat (17). This study found that the rate of sodium uptake in the presence of a pH gradient (intravesicular pHi = 5.5 and extravesicular pHi = 7.5) was higher in neonates than adult rats. The difference between the findings in this study and the present one is likely related to the techniques involved in measuring Na+/H+ antiporter activity. Uptake of sodium into a BBMV may not be dependent only on the number of Na+/H+ exchangers. The binding of sodium to the vesicle membranes, the sodium and proton permeability of the vesicle, and the lipid and protein environment of neonatal and adult brush border membranes may be different between the neonate and the adult (27, 28). These factors may be affected by preparation of membrane vesicles, which may produce an apparent increase in sodium uptake.
It is now apparent that NHE3 mediates all or the vast majority of proximal tubule apical Na+/H+ antiporter activity (3–8). To confirm our results using in vitro microperfusion of rat proximal tubules, we measured NHE3 mRNA and protein abundance. There was a maturational increase in both renal cortical NHE3 mRNA and BBMV protein abundance. These studies confirm our previous observations in the rabbit (22).
The factors that are responsible for the postnatal maturation of Na+/H+ antiporter activity are unclear at present. We have provided evidence that the postnatal rise in glucocorticoids and thyroid hormone may increase Na+/H+ antiporter activity and NHE3 mRNA and protein abundance (10, 21, 22). The present study demonstrates that there is postnatal maturation of rat proximal tubular acidification, which leads to general applicability of our previous findings in the rabbit (15). In addition, this study demonstrates that in vitro microperfusion of the rat PCT is possible and will likely be a useful tool to study rodents with targeted gene disruptions potentially affecting proximal tubule acidification.
The lower rate of bicarbonate reabsorption by the proximal tubule is likely the predominant factor responsible for the lower serum bicarbonate concentration in neonates. This study shows that there is a maturational increase in rat Na+/H+ antiporter activity as well as in NHE3 mRNA and protein abundance. These data are consistent with the low rates of proximal tubule Na+/H+ antiporter activity being an important factor that results in lower serum bicarbonate levels in neonates compared with adults.
Abbreviations
- NHE:
-
sodium hydrogen exchanger
- pHi:
-
intracellular pH
- BBMV:
-
brush border membrane vesicles
- EIPA:
-
5-(N-ethyl-N-isopropyl)-amiloride
- BCECF:
-
2′,7′-bis(carboxyethyl)-5(6)-carboxyfluorescein
- JH:
-
proton secretory rate
- PCT:
-
proximal convoluted tubule
- NaOAc:
-
sodium acetate
References
Preisig PA, Ives HE, Cragge EJ Jr, Alpern RJ, Rector FC Jr, 1987 Role of the Na+/H+ antiporter in rat proximal tubule bicarbonate absorption. J Clin Invest 80: 970–978.
Baum M 1992 Developmental changes in rabbit juxtamedullary proximal convoluted tubule acidification. Pediatr Res 31: 411–414.
Schultheis PJ, Clarke LL, Meneton P, Miller ML, Soleimani M, Gawenis LR, Riddle TM, Duffy JJ, Doetschman T, Wang T, Giebisch G, Aronson PS, Lorenz JN, Shull GE 1998 Renal and intestinal absorptive defects in mice lacking the NHE3 Na+/H+ exchanger. Nat Genet 19: 282–285.
Wang T, Yang C-L, Abbiat T, Schultheis PJ, Shull GE, Giebisch G, Aronson PS 1998 Mechanism of proximal tubule bicarbonate absorption in NHE3 knockout mice. J Am Soc Nephrol 9: 14Aabstr
Schultheis PJ, Clarke LL, Meneton P, Harline M, Boivin GP, Stemmermann G, Duffy JJ, Doetschman T, Miller ML, Shull GE 1998 Targeted disruption of the murine Na+/H+ exchanger isoform 2 gene causes reduced viability of gastric parietal cells and loss of net acid secretion. J Clin Invest 101: 1243–1253.
Wu M-S, Biemesderfer D 1996 Role of NHE3 in mediating renal brush border Na+-H+ exchange. J Biol Chem 271: 32749–32752.
Biemesderfer D, Pizzonia JH, Exner M, Reilly RF, Igarashi P, Aronson PS 1993 NHE3: a Na+/H+ exchanger isoform of the renal brush border. Am J Physiol 265: F736–F742.
Loffing J, Lotscher M, Kaissling B, Biber J, Murer H, Seikaly M, Alpern RJ, Levi M, Baum M, Moe OW 1998 Renal Na/H exchanger NHE-3 and Na-PO4 cotransporter NaPi-2 protein expression in glucocorticoid excess and deficient states. J Am Soc Nephrol 9: 1560–1567.
Schwartz GJ, Evan AP 1983 Development of solute transport in rabbit proximal tubule. I. HCO-3 and glucose absorption. Am J Physiol 245:F382–F390.
Baum M, Quigley R 1991 Prenatal glucocorticoids stimulate neonatal juxtamedullary proximal convoluted tubule acidification. Am J Physiol 261:F746–F752.
Baum M, Quigley R 1995 Ontogeny of proximal tubule acidification. Kidney Int 48: 1697–1704.
Aronson PS, Giebisch G 1997 Mechanisms of chloride transport in the proximal tubule. Am J Physiol 273:F179–F192.
Shah M, Quigley R, Baum M 1998 Maturation of rabbit proximal straight tubule chloride/base exchange. Am J Physiol 274:F883–F888.
Fukuda Y, Aperia A 1988 Differentiation of Na+-K+ pump in rat proximal tubule is modulated by Na+-H+ exchanger. Am J Physiol 255:F552–F557.
Baum M 1990 Neonatal rabbit juxtamedullary proximal convoluted tubule acidification. J Clin Invest 85: 499–506.
Beck JC, Lipkowitz MS, Abramson RG 1991 Ontogeny of Na/H antiporter activity in rabbit renal brush border membrane vesicles. J Clin Invest 87: 2067–2076.
Zelikovic I, Stejskal E, Lohstroh P, Budreau A, Chesney RW 1991 Developmental maturation of Na+-H+ exchange in rat renal tubular brush-border membrane. Am J Physiol 261:F1017–F1025.
Alpern RJ, Chambers M 1986 Cell pH in the rat proximal convoluted tubule: regulation by luminal and peritubular pH and sodium concentration. J Clin Invest 78: 502–510.
Roos A, Boron WF 1981 Intracellular pH. Physiol Rev 61: 297–434.
Preisig PA, Alpern RJ 1988 Chronic metabolic acidosis causes an adaptation in the apical membrane Na/H antiporter and basolateral membrane Na(HCO3)3 symporter in the rat proximal convoluted tubule. J Clin Invest 82: 1445–1453.
Baum M, Dwarakanath V, Alpern RJ, Moe OW 1998 Effects of thyroid hormone on the neonatal renal cortical Na+/H+ antiporter. Kidney Int 53: 1254–1258.
Baum M, Biemesderfer D, Gentry D, Aronson PS 1995 Ontogeny of rabbit renal cortical NHE3 and NHE1: effect of glucocorticoids. Am J Physiol 268:F815–F820.
Amemiya M, Loffing J, Lotscher M, Kaissling B, Alpern RJ, Moe OW 1995 Expression of NHE-3 in the apical membrane of rat renal proximal tubule and thick ascending limb. Kidney Int 48: 1206–1215.
Chomcznmski P, Sacchi N 1987 Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 162: 156–159.
Orlowski J, Kandasamy RA, Shull GE 1992 Molecular cloning of putative members of the Na/H exchanger gene family. J Biol Chem 267: 9331–9339.
Aperia A, Larsson L 1979 Correlation between fluid reabsorption and proximal tubule ultrastructure during development of the rat kidney. Acta Physiol Scand 105: 11–22.
Arar M, Levi M, Baum M 1994 Maturational effects of glucocorticoids on neonatal brush border membrane phosphate transport. Pediatr Res 35: 474–478.
Schwarz SM, Hostetler B, Ling S, Mone M, Watkins JB 1985 Intestinal membrane lipid composition and fluidity during development in the rat. Am J Physiol 248:G200–G207.
Acknowledgements
The authors thank Janell McQuinn for her secretarial assistance.
Author information
Authors and Affiliations
Additional information
Supported by National Institutes of Health grants DK41612 (M.B.) and DK48482 and the Department of Veteran Affairs Research Service (O.W.M.).
Rights and permissions
About this article
Cite this article
Shah, M., Gupta, N., Dwarakanath, V. et al. Ontogeny of Na+/H+ Antiporter Activity in Rat Proximal Convoluted Tubules. Pediatr Res 48, 206–210 (2000). https://doi.org/10.1203/00006450-200008000-00014
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-200008000-00014
This article is cited by
-
Developmental changes in renal tubular transport—an overview
Pediatric Nephrology (2015)
-
Proximal tubule specific knockout of the Na+/H+ exchanger NHE3: effects on bicarbonate absorption and ammonium excretion
Journal of Molecular Medicine (2013)
