
Abstract
Effective immunity is dependent on long-surviving memory T cells. Various memory subsets make distinct contributions to immune protection, especially in peripheral infection. It has been suggested that T cells in nonlymphoid tissues are important during local infection, although their relationship with populations in the circulation remains poorly defined. Here we describe a unique memory T cell subset present after acute infection with herpes simplex virus that remained resident in the skin and in latently infected sensory ganglia. These T cells were in disequilibrium with the circulating lymphocyte pool and controlled new infection with this virus. Thus, these cells represent an example of tissue-resident memory T cells that can provide protective immunity at points of pathogen entry.
Similar content being viewed by others
Main
Immune memory results from the action of various cellular components that combine to control infectious pathogens1. T cells are key mediators of the memory response, although uncertainty remains as to the exact function of individual memory T cell subsets2,3,4,5,6. Memory cells have been categorized as effector memory and central memory T cells, depending on their respective ability to recirculate between secondary lymphoid organs or to enter peripheral tissues5,7,8. Localized infection results in rapid population expansion of effector T cells in draining lymph nodes, and although most are lost, a substantial population of circulating memory T cells survives this contraction9. In addition, many T cells are also present in nonlymphoid tissues7,8, where their numbers may actually exceed those in the circulation10.
The relationship between peripheral and recirculating memory cells remains mostly undefined. Peripheral T cells can be replaced from the circulation in the steady state and can be further supplemented by effector memory T cells newly recruited during infection11. However, an active mechanism of T cell retention may exist in nonlymphoid tissues12,13,14, and this could explain the accumulation noted at sites that have cleared an infectious virus14,15,16. Thus, once in the periphery, at least a subset of memory cells may be separated from the circulating T cell pool.
Herpes simplex virus (HSV) can infect the skin of mice, causing a primary infection that rapidly moves to the innervating sensory ganglia17,18. There replication is controlled in about a week and then persists as a tightly controlled latent infection19. The acute infection induces a robust CD8+ T cell effector and memory response that is able to control virus in the ganglia20. The requirement for memory T cell responses in peripheral tissues has been examined with latently infected ganglia containing a long-lived population of virus-specific memory T cells21,22. It was found that these memory cells were not terminally differentiated but could instead mount a secondary proliferative response after virus reactivation22. Notably, this occurred in the ganglia itself, in the absence of any involvement of lymphoid tissue.
One issue that arose from those studies was whether the T cells residing in the ganglia were circulating memory cells or, alternatively, represented an autonomous memory population separate from the circulating pool. Although in the case of HSV, such local memory cells act to control a preexisting latent infection21,23, this type of T cell sequestration could conceivably result in more efficient protection against reinfection by an environmentally derived pathogenic agent. Here we provide evidence of a population of peripheral memory T cells that seemed to be in disequilibrium with the circulation. Notably, we show that such tissue-resident memory cells contributed to peripheral HSV control, thus effectively limiting the extent of renewed infection.
Note: Supplementary information is available on the Nature Immunology website.
Results
Nonmigrating peripheral tissue–resident memory T cells
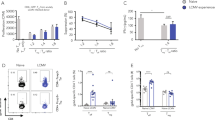
Stimulation of T cells occurs after transplantation of sensory ganglia containing both a persisting latent form of HSV and a population of CD8+ T cells specific for this pathogen22. In this model, the local T cells undergo secondary restimulation exclusively in the grafts during virus reactivation, with no apparent involvement of lymph nodes. This suggests that either the ganglionic cells form part of a true nonmigrating memory population separate from the recirculating memory pool, or the excision and transplantation of the ganglia under the kidney capsule of recipient mice affects the normal egress of T cells from this tissue. To show that recirculating memory T cells can leave the transplanted tissues, we first recruited such a migrating population into the grafts by transplanting latent ganglia into mice that had circulating HSV-specific CD8+ memory cells. We generated these primary recipients by infecting them with a recombinant influenza virus carrying the immunodominant major histocompatibility complex class I–restricted determinant from HSV glycoprotein B (gB; Supplementary Fig. 1 online). In these experiments, we 'genetically marked' the graft-resident T cells with green fluorescent protein (GFP) by using cells from gBT-I.GFP-transgenic mice, whose T cells are all specific for the immunodominant HSV gB determinant24. T cells recruited from the circulation were from gBT-I.CD45.1-transgenic mice, which meant that we could use differences in the expression of CD45 and GFP to track the respective recruited and resident memory T cell populations. It should be noted that ganglia transplantation resulted in recruitment of the circulating gBT-I cells as well as reactivation of HSV from latency caused by the trauma of neurectomy22. As a consequence, both the resident (GFP+) and the recruited (CD45.1+) HSV-specific T cells were subjected to antigen-specific stimulation, as shown by incorporation of the thymidine analog BrdU (Fig. 1a). To show that only the recruited T cells could successfully enter the circulation, we extracted ganglia from under the kidney capsule and retransplanted them into a naive, secondary host. We then challenged the new recipients with a systemic HSV infection 20 d after secondary transplantation to expand populations of cells that had left the graft, thus allowing their detection (Supplementary Fig. 1). Almost all gB-specific T cells responding to this virus challenge originated from the T cells newly recruited from the circulation (Fig. 1b,c), even though the ganglia contained roughly equal numbers of both resident (primary transplant donor) and recruited (primary transplant recipient) T cell populations (Fig. 1d). The lack of retention of recruited T cells was not due simply to their priming by recombinant influenza rather than HSV, because a repeat experiment with HSV-infected recipients of primary grafts confirmed that only the recruited T cells underwent considerable population expansion after systemic challenge (Supplementary Fig. 2 online).

(a) Flow cytometry of short-term BrdU uptake 7 d after transplantation of latently infected dorsal root ganglia containing gBT-I.GFP memory CD8+ T cells (resident) into recipient mice containing circulating OT-I.CD45.1 (Control) and gBT-I.CD45.1 (recruited) CD8+ memory T cells. Numbers above lines indicate percent BrdU+ cells (mean ± s.e.m.). Data are from two independent experiments with three to four mice. (b–d) Analysis of gBT-I populations in grafts and spleens after transplantation of latently infected ganglia containing gBT-I.GFP memory CD8+ T cells (resident (res)) into mice containing gBT-I.CD45.1 memory CD8+ T cells (recruited (rec)), followed 6 d later by retransplantation into naive recipients and, 20 d later, intravenous challenge of these recipient mice with HSV. (b) Numbers above outlined areas indicate percent recruited cells (left) and resident cells (right) among all CD8+ cells. (c) Proportion of gBT-I populations among all CD8+ cells in the spleen. (d) Recovery of gBT-I cells from grafts. Statistical analyses, paired Student's t-test. Data are from two independent experiments with five to six mice per group (mean and s.e.m. in c,d). (e) Recruited and tissue-resident gBT-I memory CD8+ T cells 9 d after retransplantation of latent ganglia, manipulated as described in b–d (original), together with a fresh latently infected ganglia graft containing no gBT-I cells (fresh). Statistical analyses, paired Student's t-test. Data are from two independent experiments with six to eight mice per group (mean and s.e.m.).
To demonstrate that the resident cells had a migration defect and did not simply lack the ability to mount a systemic response, we again recruited memory T cells into reactivating ganglia and, on day 6 after transplantation, collected the graft and retransplanted it into a naive recipient together with 'fresh' latent ganglia in a separate site under the same kidney capsule (Supplementary Fig. 3 online). On day 9 after retransplantation, we collected each graft and assessed if the infiltrating memory T cells (CD45.1+) and tissue-residing T cells (GFP+) were able to enter the neighboring ganglia, which represented a fresh site of HSV infection. We detected a substantial population of recruited memory CD8+ T cells in the fresh graft, in contrast to those resident memory T cells carried by the original graft (Fig. 1e). Thus, the resident memory T cells did not migrate to a nearby yet anatomically distinct site, even though it was equivalent to the tissue of origin for these cells.
Enrichment for 'biased' T cells after skin infection
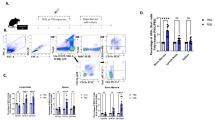
The data reported above suggested that HSV-specific T cells resident in the ganglia seemed functionally different from those found in the circulation. We reasoned that an analogous population of tissue-resident memory T cells would also be found in other peripheral sites of infection, notably the skin. Evidence for this took the form of 'preferential' retention of virus-specific T cells in flank skin involved in overt HSV disease relative to that in the uninvolved contralateral flanks. There was 'preferential' enrichment for gBT-I T cells in flank skin ipsilateral to the origin of infection at all times assessed (Fig. 2a,b). In contrast, there was no significant difference in the number of endogenous CD8+ T cells in opposing flanks after clearance of replicating virus, which occurs by day 8 after infection18 (Fig. 2c). It should be noted that gBT-I cells represented a relatively small proportion of all CD8+ T cells in the skin, especially at these later times (day 30 or beyond; Fig. 2d). The accumulation of HSV-specific T cells was not restricted to transgenic cells, because repeat experiments with tetramer staining to detect HSV-specific T cells showed similar biases (Supplementary Fig. 4 online). Retention of T cells in the skin had a nonspecific component, as demonstrated by the use of attenuated recombinant viruses (HSV strains rgB and rgB-L8A)25 (Supplementary Fig. 5 online). The number of long-term gB-specific T cells was the same in flanks infected with gB-sufficient rgB virus and control gB-deficient rgB-L8A in dual-inoculated mice. Even scarification alone led to equivalent infiltration into rgB-infected and control flank skin at day 28–30 after inoculation. Although this demonstrated a strong non-virus-specific contribution to T cell retention, the use of such attenuated viruses that do not cause skin disease could be a cause for concern. As an alternative way to address the contribution of specificity, but now using wild-type virus, we assessed the survival of virus-specific gBT-I T cells and ovalbumin-specific OT-I T cells in wild-type infected flank skin. We transferred in vitro–activated gBT-I and OT-I T cells 2–4 d after flank infection and analyzed the cells at various times thereafter. Both gBT-I and OT-I T cells survived long term in the skin, although there was a modest antigen-dependent bias (Fig. 2e), which indicated that the phenomenon is not due solely to the inflammation associated with infection or skin trauma.

(a–d) Analysis of naive gBT-I.CD45.1 CD8+ T cells transferred into C57BL/6 mice 1 d before HSV-1 infection of the left flank skin at various times after inoculation (above graphs). L, left flank (skin); R, right flank (skin); C, control skin from mock-infected mice. Statistical analyses, paired Student's t-test (b,c) or analysis of variance followed by Tukey's post-test comparison (d). Data are from at least three independent experiments with two to four mice per group (mean and s.e.m. in b,c). (a) Numbers adjacent to outlined areas indicate percent gBT-I.CD45.1 CD8+ T cells among all events. (b,c) Isolation of gBT-I.CD45.1 and endogenous CD8+ T cells. The number of gBT-I.CD45.1 T cells in skin from mock-infected mice was below the limit of detection. (d) Frequency of gBT-I.CD45.1 cells among all CD8+ T cells. (e) Proportion of gBT-I.CD45.1 T cells among all T cell antigen receptor–transgenic T cells (TCRtg; gBT-I plus OT-I) after transfer of in vitro–activated gBT-I.CD45.1 and OT-I.CD45.1 CD8+ T cells together into C57BL/6 mice 2–4 d after HSV skin infection. *, P < 0.01; **, P < 0.001 (paired Student's t-test). Data are from two to three independent experiments with three mice per group (mean and s.e.m.).
Phenotype of skin-resident memory T cells
Histological examination showed an accumulation of T cells in the epithelial layer of the skin, especially clustered around the site of cleared infection (Fig. 3a,b). Consistent with that tissue localization, long-term skin-resident T cells were CD103+ (Fig. 3c), an E-cadherin-binding molecule widely expressed by intraepithelial lymphocytes in gut and skin26,27. In contrast, their splenic counterparts were CD103−. Virus-specific skin T cells seemed to be phenotypically distinct from lymphoid memory T cells, with low expression of CD62L and CD122 but high expression of CD69 (Fig. 4a and Supplementary Fig. 6 online). Furthermore, skin T cells had higher expression of VLA-1 (Fig. 4b), an integrin that binds extracellular matrix proteins in peripheral tissues28,29. Skin-resident cells underwent slower homeostatic proliferation than did the circulating population (Fig. 4c), consistent with a dissociation of the circulating and resident populations. This contrasts with the identical homeostatic proliferation of lymphoid and lung-resident T cells after influenza infection, in which the latter is replaced by cells from from the circulating memory T cell population30,31,32.

(a,b) Microscopy of skin after transfer of naive gBT-I.GFP CD8+ T cells into C57BL/6 mice 1 d before skin infection with HSV-1. (a) Immunohistochemistry of the distribution of virus-specific gBT-I.GFP CD8+ T cells in skin obtained from previously infected areas 13 d after infection, then stained with DAPI and antibody to keratin 1 expressed in keratinocytes in the upper layer of the epidermis. Control, tissue from contralateral flanks. Arrowheads indicate gBT-I.GFP CD8+ T cells in the dermal-epidermal region. Scale bars, 100 μm. (b) Epidermal sheets from previously infected flank skin 25 d after infection, when the epithelium had reformed. The density of gBT-I.GFP CD8+ T cells is shown in the central area of lesion resolution (lesion center), on the edge of the lesion (lesion periphery) and in an unaffected area (control). Virus-specific T cells could not be detected by this method in unaffected areas in infected mice (control). Scale bars, 50 μm. (c) CD103 expression by memory gBT-I.CD45.1 CD8+ T cells isolated from spleen and epidermal sheets 13 weeks after infection of the skin with HSV. Numbers above lines (right) indicate percent CD103+ cells. Data are from one experiment representative of three.

Analysis of naive gBT-I.CD45.1 CD8+ T cells transferred into C57BL/6 mice 1 d before infection of the skin with HSV-1, followed by a period of rest for the establishment of immunological memory. (a) Flow cytometry of memory gBT-I.CD45.1 CD8+ T cells isolated from brachial lymph nodes (bLN), spleen and skin 30–386 d after infection (median, 51 d). Data represent four independent experiments with pooled cell preparations from six to seventeen mice per group (mean and s.e.m.). (b) Expression of CD49a (VLA-1) on gBT-I.CD45.1 CD8+ T cells isolated 50 d after infection. Numbers above outlined areas indicate percent CD49a+ cells. Isotype, isotype-matched control antibody. Data are from three independent experiments with two to four mice per group (mean ± s.e.m.). (c) Flow cytometry of the homeostatic proliferation of cells from 'memory mice' (43–428 d after infection; median, 72 d) treated for 7 d with BrdU. Data are from six individual experiments with three to eight mice per group (mean and s.e.m.). Statistical analyses, repeated measures analysis of variance followed by Tukey's post-test comparison.
Finally, to show that gB-specific T cells persisted in the skin in the absence of detectable virus-specific T cells in the circulation, we transplanted skin from infected flanks onto naive recipients (Supplementary Fig. 7a online). Comparison of recovered grafts and control patches of skin in the same recipients early (days 14–18; Fig. 5) or at later times (day 35; Supplementary Fig. 7b,c) showed that T cells were present only in the transplanted tissue. This was consistent with the possibility that tissue-resident memory cells are an autonomous population. Moreover, severing of the nerves that link the skin to the infected ganglia also indicated that the persistence of memory T cells was not due to continuous feeding of virus or antigen to the skin from latently infected sensory nerve bodies undergoing spontaneous bouts of reactivation, which in any case is not a property of mouse infection with HSV19.

Flow cytometry of cells from spleen and skin 2–3 weeks after transplantation of previously HSV-1-infected skin containing gBT-I.CD45.1 CD8+ memory T cells onto naive recipients (at 3–4 weeks after infection). G, skin graft; C, control (unrelated skin from the same recipient as the graft); L, left ipsilateral flank (skin). (a) Gating on propidium iodide–negative, CD45.2+ events derived from pooled grafts, pooled control skin and a recipient spleen, as well as donor tissues (HSV-infected skin and spleen). Numbers above outlined areas indicate percent gBT-I.CD45.1+ cells among all events. (b) Isolation of gBT-I.CD45.1 CD8+ T cells from graft and control skin. Each symbol represents an individual mouse; small horizontal lines indicate the mean. Statistical analysis, paired Student's t-test. Data are from two independent experiments with six mice per group.
Immune protection by tissue-resident memory T cells
As well having as 'preferential' accumulation of memory T cells, ipsilateral flanks had enhanced protection against subsequent infection. To demonstrate this without antibody interference, we infected B cell–deficient μMT mice in the flank with HSV and challenged them at various times afterward. The ipsilateral flank showed 100-fold better control of infection than that of the control contralateral flank up to 50 d after the first infection (Fig. 6a). We noted a bias toward ipsilateral protection even at more than 100 d after infection, although this was less than that present at the earlier times. The greater protection of ipsilateral flanks was virus specific, as it did not extend to heterologous infection with vaccinia virus (Supplementary Fig. 8 online).

(a) Viral titers in μMT mice infected in the left flank with HSV-1 (1° HSV) and then reinfected (time, above graphs) with HSV-1 KOS.CreTK− on both the left and right flanks and assessed 2 d later. C, control (initially mock-infected and naive mice). Data are representative of five (30 d) or two (50 d and >100 d) pooled experiments. (b) Viral titers in mice treated with depleting anti-CD4 (α-CD4) and/or anti-CD8 (α-CD8) before reinfection with HSV-1 as described in a. Data are pooled from four independent experiments with two to four mice per group. (c–e) Analysis of gBT-I.CD45.1 CD8+ T cells (c) and viral titers (d,e) in Rag1−/− mice mock-infected (control (C) in d,e) or infected with HSV-1 in the left flank and given transfer of in vitro–activated gBT-I.CD45.1 CD8+ T cells 2 d or 4 d later. (c) Isolation of gBT-I.CD45.1 CD8+ T cells 30 d after infection. Data represent four independent experiments with one to four mice per group (mean and s.e.m.). (d) Viral titers 2 d after reinfection of both flanks with HSV-1 KOS.CreTK−. Data are pooled from three independent experiments with two to five mice per group. (e) Viral titers in mice treated as described in d or also with depleting anti-CD8 before reinfection with HSV. Data are pooled from three independent experiments with two to five mice per group. Each symbol represents an individual mouse; small horizontal lines indicate the mean (a,b,d,e). Statistical analyses, one-way analysis of variance followed by Tukey's post-test comparison.
Flank immunity was T cell dependent, because elimination of CD4+ and CD8+ cells by in vivo antibody depletion diminished antiviral protection (Fig. 6b). Antibody to CD4 (anti-CD4) had a much greater effect than did depletion with anti-CD8, which suggested that CD4+ T cells dominated control of skin infection33 or that help was critical to the contribution of CD8+ T cells to virus elimination34,35. To show that local CD8+ T cells could provide antiviral protection, we injected mice deficient in recombination-activating gene 1 (Rag1−/− mice) with in vitro–activated gBT-I CD8+ T cells 2–4 d after flank infection with HSV. Previously infected skin retained more T cells than did the contralateral flank (Fig. 6c), and this was associated with 'preferential' protection against subsequent infection with HSV (Fig. 6d). Such biased protection was eliminated by treatment with anti-CD8 (Fig. 6e), which showed that it was indeed mediated by the transferred T cells. In the Rag1−/− mice, CD8+ T cells act in the complete absence of CD4+ T cell help, most likely because their numbers are in excess of those that normally persist long term after infection of wild-type hosts36. Nevertheless, these data collectively show enhanced local protection by T cells that accumulate in tissues after resolution of a previous infection.
Discussion
There are obvious advantages provided by the existence of a sequestered T cell population for persisting, reactivating infections such as the one we used here. CD8+ T cells are known to maintain HSV in a latent state in the ganglia by constant expression of effector molecules such as granzyme B and interferon-γ37,38. Whether such populations exist in acute infection systems, especially after the virus has been completely cleared, remains to be determined. There are reports of enrichment for memory T cells7,8 at sites of initial pathogen infection well after clearance has been achieved14,15,16. That is similar to what we found after infection of the skin with HSV, with a notable imbalance of antigen-specific memory T cells in regions ipsilateral or contralateral to the original site of inoculation. Published studies using rapidly cleared viruses have excluded the possibility of prolonged infection as the mechanism of persistent accumulation13,14,16,30. An argument could conceivably be mounted for the possibility that ongoing gB-specific stimulation impedes the release of circulating memory T cells, given the nature of HSV latency. However, experiments with bone marrow–chimeric mice have shown that even in the sensory ganglia, T cells persist in the absence of specific recognition of infected neurons39. Retention in the skin by ongoing virus reactivation seems even less likely, given our demonstration that resident T cells survived transplantation and thus separation from the neuronal source of infection. Unlike humans, mice do not show spontaneous reactivation of HSV19, and skin-resident T cells show no signs of continuous antigen stimulation, such as the upregulation of granzymes, which is evident in T cells persisting in the ganglia40. Thus, although ongoing antigen presentation remains a formal possibility, it is unlikely to fully explain the retention of HSV-specific peripheral T cells, especially in the skin.
Although localized infection can 'imprint' 'preferential' T cell migration41,42, and this could explain other cases of local accumulation of T cells, it is difficult to see how such tissue-tropic infiltration would contribute to the 'side-biased' T cell accumulation in the flank skin shown here. Moreover, the lack of T cell migration between the two ganglia transplanted under the same kidney capsule indicates that these tissue-resident T cells simply did not migrate between anatomically distinct but otherwise equivalent tissues. As a consequence, we propose these HSV-specific T cells represent a sessile or nonmigratory memory subset that we define as 'tissue-resident memory T cells'. These cells are either separate from the recirculating pool or retained long enough in peripheral tissues to infrequently exchange with this population. Therefore, they are different from other memory T cells, including the central and effector memory T cells found in the wider circulation4,5.
CD8+ T cells in the ganglia can mount a stand-alone new immune response that includes helper T cell–dependent proliferation and stimulation by dendritic cells recruited from the circulation22. Together with our data here, this suggests that tissue-resident memory T cells are not terminally differentiated but can self-renew either through homeostatic or antigen-stimulatory mechanisms. Furthermore, they suggest that studies confined to circulating effector memory and central memory T cell populations may underestimate the totality of immunological memory in a given person.
Persisting T cells have been found in human skin that has cleared HSV infection43, and embedded T cells are associated with recurring allergic responses in humans, known as 'fixed drug eruptions', driven by CD8+ T cells in the epidermis that do not disseminate throughout the skin44,45. In addition, pathogenic T cells persist in pre-psoriatic human skin, and these cells can be transferred during mouse xenotransplantation46. In those last two examples, the residing T cells express markers common to the tissue-resident memory T cells in our study, such as VLA-1 and CD103; the latter are important for epithelial localization26. Finally, we have shown that these memory T cells can efficiently inhibit HSV replication in peripheral nonlymphoid tissues, including those at the body surface. Thus, our study raises the possibility that resident memory cells can be exploited as a means of controlling infection at body surfaces that act as portals of entry for invading pathogens.
Methods
Mice.
C57BL/6, B6.SJL-PtprcaPep3b/BoyJ (B6.CD45.1), gBT-I, gBT-I × B6.CD45.1 (gBT-I.CD45.1), gBT-I.GFP, OT-I × B6.CD45.1 (OT-I.CD45.1), μMT and Rag1−/−/Je (Rag1−/−) mice were bred in the Department of Microbiology and Immunology of The University of Melbourne in Parkville, Australia. The gBT-I and OT-I mice are CD8+ T cell receptor-transgenic mice that recognize the H-2Kb-restricted HSV-1 gB epitope of amino acids 498–505 (gB(498–505)) and the ovalbumin-derived epitope of amino acids 257–264 (OVA(257–264)), respectively. B6.CD45.1 mice express the congenic marker CD45.1, in contrast to C57BL/6, μMT and Rag1−/− mice, which express CD45.2. The gBT-I.CD45.1 and OT-I.CD45.1 mice were F1 offspring expressing both CD45.1 and CD45.2. Animal experiments were approved by The University of Melbourne Animal Ethics Committee.
Virus infection.
Viruses used were HSV-1 KOS, KOS.CreTK−, KOS.rgB25 (rgB) and KOS.rgB-L8A (rgB-L8A)25, as well as WSN/NA/gB (flu-gB; from S. Tevethia), WSN/NA/OVA (flu-OVA; from P. Doherty) and vaccinia-NP (from J. Yewdell). The recombinant influenza viruses express gB(498–505) (flu-gB) or OVA(257–264) (flu-OVA) in their neuraminidase stalk. Routes of infections were epidermal (skin; 1 × 106 plaque-forming units (PFU)), subcutaneous (5 × 102 PFU influenza virus and 1 × 106 PFU HSV) and intravenous (1 × 105 PFU). Skin was infected with HSV-1 KOS and KOS.CreTK− by scarification as described18. The HSV-1 KOS strain caused typical zosteriform lesions spreading from the primary inoculation site along the affected dermatomes on the same flank. The other HSV strains used had attenuated skin replication and did not cause any visible zosteriform lesions. Skin was infected with vaccinia-NP (5 × 107 PFU) by inoculation of flank skin as for HSV infection. For subcutaneous infection, 25 μl of a virus suspension was injected into foot hocks.
In vitro activation and adoptive transfer of transgenic CD8+ T cells.
All adoptive transfers of gBT-I and OT-I cells were done intravenously with lymph node suspensions or in vitro–generated effector splenocytes activated by peptide-pulsed targets as described18. First, gBT-I or OT-I splenocytes were cultured for 4 d together with gB(498–505)- or OVA(257–264)-pulsed (1 μg/ml) C57BL/6 splenocytes, respectively. On days 2 and 3, cultures were split 1: 2 and interleukin 2 (10 U/ml; PeproTech) was added. Then, 1 × 106 to 1.5 × 106 effector cells derived from those cultures were transferred intravenously into mice whose skin was infected with HSV-1. Adoptive transfer of naive gBT-I.CD45.1 cells (5 × 104) resulted in a frequency of gBT-I cells roughly ten times higher than that of their endogenous HSV-1-specific counterparts47.
Flow cytometry of tissue cells.
Mice were killed by carbon dioxide administration and were perfused with 10 ml Hank's buffered-salt solution (Media Preparation Unit, The University of Melbourne). Lymph nodes and spleens were collected and disrupted by passage through a wire mesh. Skin tissue (1–2 cm2) and ganglia were removed, were chopped into small fragments and were incubated for 90 min at 37 °C in Eagle's minimum essential medium containing 2% (vol/vol) FCS (Gibco BRL), collagenase (3 mg/ml; Worthington) and DNase (5 μg/ml; Sigma). For minimization of the digestion of certain surface receptors, this incubation period was decreased to 30 min in some phenotyping experiments. Thereafter, cell suspensions were filtered twice through nylon meshes (pore size, 70 μm and 20 μm) and were stained for 30 min with antibodies for flow cytometry. The following antibodies were from BD Pharmingen: allophycocyanin- and phycoerythrin-indotricarbocyanine–conjugated anti-CD8α (53-6.7), fluorescein isothiocyanate– and phycoerythrin-conjugated anti-CD45.1 (A20), fluorescein isothiocyanate–conjugated anti-CD45.2 (104), phycoerythrin-conjugated anti-CD4 (GK1.5), anti-CD49a (Ha31/8), anti–keyhole limpet hemocyanin hamster IgG2 553962, fluorescein isothiocyanate–conjugated anti-CD44 (IM7), phycoerythrin-conjugated anti-Vα2 (B20.1), phycoerythrin-conjugated anti-CD69 (H1.2F3), phycoerythrin-conjugated anti-CD122 (TM-β1), fluorescein isothiocyanate–conjugated anti-CD62L (Mel-14), fluorescein isothiocyanate–conjugated antibody to Armenian and Syrian hamster IgG (G192-1), and anti-CD16/32 (2.4G2). Allophycocyanin-conjugated anti-CD45.1 (A20), phycoerythrin- and allophycocyanin–Alexa Fluor 750–conjugated anti-CD45.2 (104) and fluorescein isothiocyanate–conjugated anti-CD103 (2E7) were from eBiosciences. H-2Kb gB(498–505)–phycoerythrin tetramer was generated at the Department of Microbiology and Immunology of The University of Melbourne. Dead cells were excluded by propidium iodide staining. Sphero calibration particles (BD Pharmingen) were added to samples to allow calculation of cell numbers. For analysis of the homeostatic turnover of memory cells, 1 mg sterile BrdU (5-bromodeoxyuridine) was injected intraperitoneally on 7 consecutive days. For short-pulse experiments, 1.25 mg BrdU was injected and mice were analyzed 1 h later. Uptake was detected with a BrdU Flow kit according to the manufacturer's instructions (BD Pharmingen). A FACSCalibur or FACSCanto II, plus Cell QuestPro software (BD Biosciences) and FlowJo software (TreeStar), were used for analysis.
Transplantation experiments.
Latently infected dorsal root ganglia were transplanted under the kidney capsules of syngeneic recipients as described22. For transplantation of previously HSV-infected skin tissue, donor mice were killed by carbon dioxide administration 2–4 weeks after HSV infection of the left flank skin. After flank skin was clipped and then treated with Veet depilation cream (Reckitt Benckiser), mice were perfused through the left ventricle with 10 ml Hank's balanced-salt solution. Donor skin tissue (previously infected skin with an area of 1–1.5 cm × 1 cm) was removed in aseptic conditions for storage in ice-cold buffer. Naive recipient mice were anesthetized and the skin of their flanks and backs was clipped, was treated with Veet cream and was disinfected with 70% (vol/vol) ethanol. Graft beds were prepared by carefully snipping away of the skin layer with sterile curved scissors. Finally, graft tissue was placed onto the graft bed and was secured with Jelonet gauze dressing (Smith & Nephew) as well as Micropore and Transpore tape (both from 3M Health Care). For the next 4 d, recipient mice were given analgesics in their drinking water (paracetamol; 1.3 mg/ml). After 8–10 d, bandages were removed and engraftment of transplanted skin was monitored daily.
Immunohistochemistry.
Skin tissues were fixed for 2 h on ice in 4% (vol/vol) paraformaldehyde and 10% (wt/vol) sucrose in PBS and were frozen in Tissue Tek (Sakura Finetek). Sections were cut on a cryomicrotome (CM3050S; Leica), were air-dried and were fixed in ice-cold acetone. Epidermal sheets were prepared by incubation of full-thickness skin with Dispase II (2.5 mg/ml; Roche), followed by manual separation of the dermal and epidermal layers and fixation in 4% (vol/vol) paraformaldehyde and 10% (wt/vol) sucrose in PBS. Keratin expression was visualized by incubation with polyclonal rabbit-anti–mouse keratin 1 (AF 109; Covance) followed by Alexa Fluor 594–conjugated donkey anti-rabbit (A21207; Molecular Probes). Slides were mounted with Vectashield containing DAPI (4,6-diamidino-2-phenylindole; Vector Laboratories). Images were acquired with a fluorescence microscope (DMI 4000B) and digital camera (DFC 490) with IM50 software (all from Leica) and the Adobe Photoshop program.
References
Welsh, R.M., Selin, L.K. & Szomolanyi-Tsuda, E. Immunological memory to viral infections. Annu. Rev. Immunol. 22, 711–743 (2004).
Bachmann, M.F., Kundig, T.M., Hengartner, H. & Zinkernagel, R.M. Protection against immunopathological consequences of a viral infection by activated but not resting cytotoxic T cells: T cell memory without “memory T cells”? Proc. Natl. Acad. Sci. USA 94, 640–645 (1997).
Bachmann, M.F., Wolint, P., Schwarz, K. & Oxenius, A. Recall proliferation potential of memory CD8+ T cells and antiviral protection. J. Immunol. 175, 4677–4685 (2005).
Sallusto, F., Geginat, J. & Lanzavecchia, A. Central memory and effector memory T cell subsets: function, generation, and maintenance. Annu. Rev. Immunol. 22, 745–763 (2004).
Sallusto, F., Lenig, D., Forster, R., Lipp, M. & Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 401, 708–712 (1999).
Wherry, E.J. et al. Lineage relationship and protective immunity of memory CD8 T cell subsets. Nat. Immunol. 4, 225–234 (2003).
Masopust, D., Vezys, V., Marzo, A.L. & Lefrancois, L. Preferential localization of effector memory cells in nonlymphoid tissue. Science 291, 2413–2417 (2001).
Reinhardt, R.L., Khoruts, A., Merica, R., Zell, T. & Jenkins, M.K. Visualizing the generation of memory CD4 T cells in the whole body. Nature 410, 101–105 (2001).
Kaech, S.M., Wherry, E.J. & Ahmed, R. Effector and memory T-cell differentiation: implications for vaccine development. Nat. Rev. Immunol. 2, 251–262 (2002).
Clark, R.A. et al. The vast majority of CLA+ T cells are resident in normal skin. J. Immunol. 176, 4431–4439 (2006).
Hikono, H. et al. T-cell memory and recall responses to respiratory virus infections. Immunol. Rev. 211, 119–132 (2006).
Harris, N.L., Watt, V., Ronchese, F. & Le Gros, G. Differential T cell function and fate in lymph node and nonlymphoid tissues. J. Exp. Med. 195, 317–326 (2002).
Klonowski, K.D. et al. Dynamics of blood-borne CD8 memory T cell migration in vivo. Immunity 20, 551–562 (2004).
Masopust, D. et al. Activated primary and memory CD8 T cells migrate to nonlymphoid tissues regardless of site of activation or tissue of origin. J. Immunol. 172, 4875–4882 (2004).
Hawke, S., Stevenson, P.G., Freeman, S. & Bangham, C.R. Long-term persistence of activated cytotoxic T lymphocytes after viral infection of the central nervous system. J. Exp. Med. 187, 1575–1582 (1998).
Hogan, R.J. et al. Activated antigen-specific CD8+ T cells persist in the lungs following recovery from respiratory virus infections. J. Immunol. 166, 1813–1822 (2001).
Simmons, A. & Nash, A.A. Zosteriform spread of herpes simplex virus as a model of recrudescence and its use to investigate the role of immune cells in prevention of recurrent disease. J. Virol. 52, 816–821 (1984).
van Lint, A. et al. Herpes simplex virus-specific CD8+ T cells can clear established lytic infections from skin and nerves and can partially limit the early spread of virus after cutaneous inoculation. J. Immunol. 172, 392–397 (2004).
Stanberry, L.R. Pathogenesis of herpes simplex virus infection and animal models for its study. Curr. Top. Microbiol. Immunol. 179, 15–30 (1992).
Simmons, A. & Tscharke, D.C. Anti-CD8 impairs clearance of herpes simplex virus from the nervous system: implications for the fate of virally infected neurons. J. Exp. Med. 175, 1337–1344 (1992).
Khanna, K.M., Bonneau, R.H., Kinchington, P.R. & Hendricks, R.L. Herpes simplex virus-specific memory CD8+ T cells are selectively activated and retained in latently infected sensory ganglia. Immunity 18, 593–603 (2003).
Wakim, L.M., Waithman, J., van Rooijen, N., Heath, W.R. & Carbone, F.R. Dendritic cell-induced memory T cell activation in nonlymphoid tissues. Science 319, 198–202 (2008).
Liu, T., Khanna, K.M., Chen, X., Fink, D.J. & Hendricks, R.L. CD8+ T cells can block herpes simplex virus type 1 (HSV-1) reactivation from latency in sensory neurons. J. Exp. Med. 191, 1459–1466 (2000).
Mueller, S.N., Heath, W., McLain, J.D., Carbone, F.R. & Jones, C.M. Characterization of two TCR transgenic mouse lines specific for herpes simplex virus. Immunol. Cell Biol. 80, 156–163 (2002).
Stock, A.T., Jones, C.M., Heath, W.R. & Carbone, F.R. CTL response compensation for the loss of an immunodominant class I-restricted HSV-1 determinant. Immunol. Cell Biol. 84, 543–550 (2006).
Cepek, K.L. et al. Adhesion between epithelial cells and T lymphocytes mediated by E-cadherin and the αEβ7 integrin. Nature 372, 190–193 (1994).
Pauls, K. et al. Role of integrin αE(CD103)β7 for tissue-specific epidermal localization of CD8+ T lymphocytes. J. Invest. Dermatol. 117, 569–575 (2001).
Hemler, M.E. VLA proteins in the integrin family: structures, functions, and their role on leukocytes. Annu. Rev. Immunol. 8, 365–400 (1990).
Ray, S.J. et al. The collagen binding α1β1 integrin VLA-1 regulates CD8 T cell-mediated immune protection against heterologous influenza infection. Immunity 20, 167–179 (2004).
Ely, K.H., Cookenham, T., Roberts, A.D. & Woodland, D.L. Memory T cell populations in the lung airways are maintained by continual recruitment. J. Immunol. 176, 537–543 (2006).
Hogan, R.J. et al. Long-term maintenance of virus-specific effector memory CD8+ T cells in the lung airways depends on proliferation. J. Immunol. 169, 4976–4981 (2002).
Zammit, D.J., Turner, D.L., Klonowski, K.D., Lefrancois, L. & Cauley, L.S. Residual antigen presentation after influenza virus infection affects CD8 T cell activation and migration. Immunity 24, 439–449 (2006).
Nash, A.A. et al. Different roles for L3T4+ and Lyt2+ T cell subsets in the control of an acute herpes simplex virus infection of the skin and nervous system. J. Gen. Virol. 68, 825–833 (1987).
Jennings, S.R., Bonneau, R.H., Smith, P.M., Wolcott, R.M. & Chervenak, R. CD4-positive T lymphocytes are required for the generation of the primary but not the secondary CD8-positive cytolytic T lymphocyte response to herpes simplex virus in C57BL/6 mice. Cell. Immunol. 133, 234–252 (1991).
Smith, C.M. et al. Cognate CD4+ T cell licensing of dendritic cells in CD8+ T cell immunity. Nat. Immunol. 5, 1143–1148 (2004).
Mintern, J.D., Davey, G.M., Belz, G.T., Carbone, F.R. & Heath, W.R. Cutting edge: precursor frequency affects the helper dependence of cytotoxic T cells. J. Immunol. 168, 977–980 (2002).
Knickelbein, J.E. et al. Noncytotoxic lytic granule-mediated CD8+ T cell inhibition of HSV-1 reactivation from neuronal latency. Science 322, 268–271 (2008).
Liu, T., Khanna, K.M., Carriere, B.N. & Hendricks, R.L. Gamma interferon can prevent herpes simplex virus type 1 reactivation from latency in sensory neurons. J. Virol. 75, 11178–11184 (2001).
van Lint, A.L. et al. Latent infection with herpes simplex virus is associated with ongoing CD8+ T-cell stimulation by parenchymal cells within sensory ganglia. J. Virol. 79, 14843–14851 (2005).
Mintern, J.D., Guillonneau, C., Carbone, F.R., Doherty, P.C. & Turner, S.J. Cutting edge: tissue-resident memory CTL down-regulate cytolytic molecule expression following virus clearance. J. Immunol. 179, 7220–7224 (2007).
Campbell, D.J. & Butcher, E.C. Rapid acquisition of tissue-specific homing phenotypes by CD4+ T cells activated in cutaneous or mucosal lymphoid tissues. J. Exp. Med. 195, 135–141 (2002).
Mora, J.R. & von Andrian, U.H. T-cell homing specificity and plasticity: new concepts and future challenges. Trends Immunol. 27, 235–243 (2006).
Zhu, J. et al. Virus-specific CD8+ T cells accumulate near sensory nerve endings in genital skin during subclinical HSV-2 reactivation. J. Exp. Med. 204, 595–603 (2007).
Mizukawa, Y. et al. Direct evidence for interferon-γ production by effector-memory-type intraepidermal T cells residing at an effector site of immunopathology in fixed drug eruption. Am. J. Pathol. 161, 1337–1347 (2002).
Shiohara, T. & Moriya, N. Epidermal T cells: their functional role and disease relevance for dermatologists. J. Invest. Dermatol. 109, 271–275 (1997).
Boyman, O. et al. Spontaneous development of psoriasis in a new animal model shows an essential role for resident T cells and tumor necrosis factor-α. J. Exp. Med. 199, 731–736 (2004).
Stock, A.T. et al. Optimization of TCR transgenic T cells for in vivo tracking of immune responses. Immunol. Cell Biol. 85, 394–396 (2007).
Acknowledgements
We thank S. Tevethia (Pennsylvania State University) for WSN/NA/gB; P. Doherty (The University of Melbourne) for WSN/NA/OVA; J. Yewdell (U.S. National Institutes of Health) for vaccinia-NP.; D. Masopust for discussions; and H. Kosaka for suggesting that fixed drug eruptions are caused by resident T cells. Supported by the Australian National Health and Medical Research Council, the Howard Hughes Medical Institute and the German Research Foundation (GE1666/1-1 to T.G.).
Author information
Authors and Affiliations
Corresponding authors
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–8 (PDF 4421 kb)
Rights and permissions
About this article
Cite this article
Gebhardt, T., Wakim, L., Eidsmo, L. et al. Memory T cells in nonlymphoid tissue that provide enhanced local immunity during infection with herpes simplex virus. Nat Immunol 10, 524–530 (2009). https://doi.org/10.1038/ni.1718
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ni.1718
This article is cited by
-
Assessing the generation of tissue resident memory T cells by vaccines
Nature Reviews Immunology (2023)
-
Localization, tissue biology and T cell state — implications for cancer immunotherapy
Nature Reviews Immunology (2023)
-
FGL2-targeting T cells exhibit antitumor effects on glioblastoma and recruit tumor-specific brain-resident memory T cells
Nature Communications (2023)
-
Unique properties of tissue-resident memory T cells in the lungs: implications for COVID-19 and other respiratory diseases
Nature Reviews Immunology (2023)
-
Human circulating and tissue-resident memory CD8+ T cells
Nature Immunology (2023)
