
Abstract
Thousands of putative biosynthetic genes in Arabidopsis thaliana have no known function, which suggests that there are numerous molecules contributing to plant fitness that have not yet been discovered1,2. Prime among these uncharacterized genes are cytochromes P450 upregulated in response to pathogens3,4. Here we start with a single pathogen-induced P450 (ref. 5), CYP82C2, and use a combination of untargeted metabolomics and coexpression analysis to uncover the complete biosynthetic pathway to 4-hydroxyindole-3-carbonyl nitrile (4-OH-ICN), a previously unknown Arabidopsis metabolite. This metabolite harbours cyanogenic functionality that is unprecedented in plants and exceedingly rare in nature6,7; furthermore, the aryl cyanohydrin intermediate in the 4-OH-ICN pathway reveals a latent capacity for cyanogenic glucoside biosynthesis8,9 in Arabidopsis. By expressing 4-OH-ICN biosynthetic enzymes in Saccharomyces cerevisiae and Nicotiana benthamiana, we reconstitute the complete pathway in vitro and in vivo and validate the functions of its enzymes. Arabidopsis 4-OH-ICN pathway mutants show increased susceptibility to the bacterial pathogen Pseudomonas syringae, consistent with a role in inducible pathogen defence. Arabidopsis has been the pre-eminent model system10,11 for studying the role of small molecules in plant innate immunity12; our results uncover a new branch of indole metabolism distinct from the canonical camalexin pathway, and support a role for this pathway in the Arabidopsis defence response13. These results establish a more complete framework for understanding how the model plant Arabidopsis uses small molecules in pathogen defence.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


References
Chae, L., Kim, T., Nilo-Poyanco, R. & Rhee, S. Y. Genomic signatures of specialized metabolism in plants. Science 344, 510–513 (2014)
D'Auria, J. C. & Gershenzon, J. The secondary metabolism of Arabidopsis thaliana: growing like a weed. Curr. Opin. Plant Biol. 8, 308–316 (2005)
Bednarek, P. & Osbourn, A. Plant-microbe interactions: chemical diversity in plant defense. Science 324, 746–748 (2009)
Denoux, C. et al. Activation of defense response pathways by OGs and Flg22 elicitors in Arabidopsis seedlings. Mol. Plant 1, 423–445 (2008)
Bak, S. et al. Cytochromes P450. Arabidopsis Book 9, e0144 (2011)
Jones, T. H., Conner, W. E., Meinwald, J., Eisner, H. E. & Eisner, T. Benzoyl cyanide and mandelonitrile in the cyanogenetic secretion of a centipede. J. Chem. Ecol. 2, 421–429 (1976)
Zagrobelny, M., Bak, S. & Møller, B. L. Cyanogenesis in plants and arthropods. Phytochemistry 69, 1457–1468 (2008)
Gleadow, R. M. & Møller, B. L. Cyanogenic glycosides: synthesis, physiology, and phenotypic plasticity. Annu. Rev. Plant Biol. 65, 155–185 (2014)
Tattersall, D. B. et al. Resistance to an herbivore through engineered cyanogenic glucoside synthesis. Science 293, 1826–1828 (2001)
Bednarek, P. et al. A glucosinolate metabolism pathway in living plant cells mediates broad-spectrum antifungal defense. Science 323, 101–106 (2009)
Clay, N. K., Adio, A. M., Denoux, C., Jander, G. & Ausubel, F. M. Glucosinolate metabolites required for an Arabidopsis innate immune response. Science 323, 95–101 (2009)
Dangl, J. L., Horvath, D. M. & Staskawicz, B. J. Pivoting the plant immune system from dissection to deployment. Science 341, 746–751 (2013)
Ahuja, I., Kissen, R. & Bones, A. M. Phytoalexins in defense against pathogens. Trends Plant Sci. 17, 73–90 (2012)
Vinayavekhin, N. & Saghatelian, A. in Current Protocols in Molecular Biology (eds Ausubel, F. M. et al.) Ch. 30 (2010)
Böttcher, C. et al. The biosynthetic pathway of indole-3-carbaldehyde and indole-3-carboxylic acid derivatives in Arabidopsis. Plant Physiol. 165, 841–853 (2014)
Mikkelsen, M. D., Hansen, C. H., Wittstock, U. & Halkier, B. A. Cytochrome P450 CYP79B2 from Arabidopsis catalyzes the conversion of tryptophan to indole-3-acetaldoxime, a precursor of indole glucosinolates and indole-3-acetic acid. J. Biol. Chem. 275, 33712–33717 (2000)
Zhao, Y. et al. Trp-dependent auxin biosynthesis in Arabidopsis: involvement of cytochrome P450s CYP79B2 and CYP79B3. Genes Dev. 16, 3100–3112 (2002)
Millet, Y. A. et al. Innate immune responses activated in Arabidopsis roots by microbe-associated molecular patterns. Plant Cell 22, 973–990 (2010)
Boudsocq, M. et al. Differential innate immune signalling via Ca2+ sensor protein kinases. Nature 464, 418–422 (2010)
Peyret, H. & Lomonossoff, G. P. The pEAQ vector series: the easy and quick way to produce recombinant proteins in plants. Plant Mol. Biol. 83, 51–58 (2013)
Thomma, B. P., Nelissen, I., Eggermont, K. & Broekaert, W. F. Deficiency in phytoalexin production causes enhanced susceptibility of Arabidopsis thaliana to the fungus Alternaria brassicicola. Plant J. 19, 163–171 (1999)
Gómez-Gómez, L. & Boller, T. Flagellin perception: a paradigm for innate immunity. Trends Plant Sci. 7, 251–256 (2002)
Zipfel, C. et al. Bacterial disease resistance in Arabidopsis through flagellin perception. Nature 428, 764–767 (2004)
Liu, F. et al. The Arabidopsis P450 protein CYP82C2 modulates jasmonate-induced root growth inhibition, defense gene expression and indole glucosinolate biosynthesis. Cell Res. 20, 539–552 (2010)
Nafisi, M. et al. Arabidopsis cytochrome P450 monooxygenase 71A13 catalyzes the conversion of indole-3-acetaldoxime in camalexin synthesis. Plant Cell 19, 2039–2052 (2007)
Nelson, D. & Werck-Reichhart, D. A P450-centric view of plant evolution. Plant J. 66, 194–211 (2011)
Møller, B. L. Functional diversifications of cyanogenic glucosides. Curr. Opin. Plant Biol. 13, 338–347 (2010)
Rauhut, T. & Glawischnig, E. Evolution of camalexin and structurally related indolic compounds. Phytochemistry 70, 1638–1644 (2009)
Acknowledgements
We thank F. Ausubel, M. B. Mudgett, C. Khosla, A. Saghatelian, Y. Millet, C. Danna, S. Galanie, and members of the Sattely and Clay laboratories for advice on experiments and comments on the manuscript. We thank the Salk Institute Genomic Analysis Laboratory for providing the sequence-indexed Arabidopsis transfer-DNA insertion mutants. We thank G. Lomonossoff (John Innes Centre) for providing plasmid pEAQ. This work was supported by R00 GM089985 and DP2 AT008321 (to E.S.S.), T32 GM008412-20 (to J.R.), and T32 GM007499-38 (to B.B.). The early stages of this work were supported by National Science Foundation grant MCB-0519898 and National Institutes of Health grant R37 GM 48707 (awarded to Fred Ausubel, Massachusetts General Hospital, Boston, Massachusetts, USA).
Author information
Authors and Affiliations
Contributions
J.R., B.B., N.K.C., and E.S.S. designed experiments. J.R. and B.B. performed experiments. J.R., B.B., N.K.C., and E.S.S. analysed data and wrote the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
Extended Data Figure 1 Elicitation of compounds identified in metabolomics screen by flg22 peptide and origin of ICA methyl ester as artefact of the methanol extraction method.
a, Levels of compounds in Flg22-elicited Arabidopsis Col-0 seedling tissue, quantified as mean [M + H]+ ion (m/z ± 10 ppm) abundances extracted from raw data; error bars, s.d. based on three biological replicates. Production of these compounds in axenic plant culture demonstrates that they are plant-derived. b, c, Structure and mass peaks of ICA methyl ester (compound A1) seen in LC–MS analysis (b), and EICs for the expected m/z using a standard extraction with 80:20 CH3OH/H2O or with 80:20 CD3OD/D2O (c). d, e, Structure and mass spectrum peaks seen for the triply deuterated A1 analogue (d), and EICs for the expected m/z using extraction with 80:20 CH3OH/H2O, or with 80:20 CD3OD/D2O (all EICs are to scale) (e). The presence of the deuterated analogue of ICA methyl ester and the complete absence of the non-deuterated compound in plant extracts when CD3OD is substituted for CH3OH show that the methyl ester is not a product of Arabidopsis metabolism, but arises because of the extraction method as a degradation product of ICN.
Extended Data Figure 2 Comparison of spectra for plant-extracted and synthetic compound establishes identity of ICN as new indolic metabolite produced by A. thaliana.
a, Full-range (δ = 10.5 to −0.5) and, b, downfield region partial (δ = 8.5–7.0) 1H NMR spectra in CD3CN. Upfield contaminants in the full-range spectra are presumed to be residual solvent. c, Ultraviolet–visible absorbance spectra obtained via a diode array detector during LC analysis. Note that the prominent peak at 230 nm is due to acetonitrile in the LC mobile phase. d, Targeted MS/MS spectra for the parent ICN [M + H]+ ion (m/z = 171.0550) at a collision energy of 20 V. See Supplementary Table 1 for relative peak intensities at other collision energies. e, Aligned EICs for the ICN [M + H]+ ion for a Col-0 + Psta tissue sample extracted with DMSO and synthetic compound, showing identical retention times.
Extended Data Figure 3 Comparison of plant-extracted ICN derivatives, 4-OH-ICN derivatives, and synthetic standards shows identical column elution times for all compounds.
Col-0 + Psta combined EICs were extracted for the relevant compound [M + H]+ m/z values for a DMSO-extracted medium sample (4-OH-ICN trace), or a MeOH-extracted seedling tissue sample (all other traces), while synthetic EICs were extracted for a mixed standard in DMSO. Note that chromatograms are not to scale, and the synthetic standard is not equimolar with respect to all compounds because of partial degradation.
Extended Data Figure 4 CYP82C2 is an ICN 4-hydroxylase.
a, b, 1H NMR spectra in CD3OD of synthetic ICA (a) and 4-OH-ICA (b). c, Spectrum for large-scale enzymatic reaction extract of ICN incubated with CYP82C2. In addition to ICA, resulting from hydrolysis of ICN, peaks for a singly hydroxylated analogue of ICA are seen; these are qualitatively consistent with, but shifted slightly upfield (∼30–60 Hz) from the 4-OH-ICA spectrum, possibly because of impurities or a pH effect in the enzymatic reaction sample. d, To confirm the identity conclusively, 80 μg of 4-OH-ICA dissolved in CD3OD was added to the enzymatic reaction NMR sample before acquiring another spectrum: no new peaks are seen, while the prior hydroxylated ICA peaks grow in intensity, establishing the product of the enzymatic reaction as 4-OH-ICA. e, EICs for enzymatic reactions of CYP82C2 on ICN or ICA, or empty vector control incubation with ICN. Only trace amounts of the expected 4-OH-ICN product but significant amounts of 4-OH-ICA are seen for the CYP82C2/ICN reaction. No hydroxylated products are seen for the CYP82C2/ICA or empty vector/ICN reactions, indicating that CYP82C2 catalyses only the hydroxylation of ICN to 4-OH-ICN, but 4-OH-ICA is seen as the predominant end product due to rapid hydrolysis of 4-OH-ICN (f). Chromatograms in this figure were obtained using the 20 min LC–MS gradient (see Supplementary Information, Methods section 1.9 LC–MS analysis).
Extended Data Figure 5 Levels of numerous Arabidopsis indolic metabolites are altered in ICN pathway gene insertion lines compared with WT plants.
a–e, Relative compound levels for mock treatment condition and indicated pathway insertion line mutants, and, f, absolute levels in Psta-treated WT (Col-0) seedlings. For a–e, data bars represent a logarithmically scaled ratio of mean metabolite levels in the indicated line or treatment condition, quantified as [M + H]+ ion abundances by LC–MS analysis with XCMS processing, to levels in Psta-treated WT Arabidopsis seedlings. In f, absolute levels for all compounds except RA were quantified by measuring [M + H]+ ion abundances and comparing to standard curves. Error bars, s.d., based on six biological replicates. Cam, camalexin; RA, raphanusamic acid; other abbreviations as detailed previously.
Extended Data Figure 6 Putative ICGs observed in Arabidopsis and in N. benthamiana expressing ICN pathway enzymes.
a, EICs for putative ICGs in WT Arabidopsis and fox mutant elicited with Psta. The m/z values shown are median values calculated by XCMS. b, Hypothesized structures and theoretical m/z values for the two ICGs identified. c, MS/MS spectrum for ICG1; m/z values and relative abundances are shown above each peak. The ion analysed here (m/z = 691.2210) represents a [2M + Na]+ dimer that is significantly more abundant than the [M + Na]+ ion. Direct analysis of the [M + Na]+ ion (m/z = 357.1057) yielded low abundance spectra that could not be easily analysed. At lower collision energies, the [2M + Na]+ ion fragments to [M + Na]+, but yields a rich spectrum at 40 V, which is shown. Predicted peak assignments for the ICG1 MS/MS spectrum are shown in the accompanying table. For peaks in bold, exact counterparts could be identified in the dhurrin [M + Na]+ 20 V MS/MS spectrum in the METLIN metabolite database. d, MS/MS spectrum obtained for the ICG2 [M + Na]+ ion and predicted peak assignments. While the [2M + Na]+ peak (m/z = 864.2225) is also seen for this compound (not shown), [M + Na]+ is more abundant in this case, and was analysed directly. e, f, Levels of ICG1 and ICG2 in ICN pathway mutants (e) and in WT plants elicited with Psta and N. benthamiana expressing ICN pathway enzymes (f). For e and f, levels are quantified as mean [M + Na]+ ion (m/z ± 10 ppm) abundances extracted from raw data; error bars, s.d., based on six biological replicates.
Extended Data Figure 7 ICN pathway metabolites are produced in N. benthamiana transiently expressing pathway genes.
Levels of ICN and 4-OH-ICN derivatives (left axis) and other relevant indolic compounds (right axis), quantified as mean [M + H]+ ion (m/z ± 10 ppm) abundances extracted from raw data; error bars, s.d., based on six biological replicates. The set of transiently expressed genes is indicated for each panel. Background levels of ICA and IAL detected when only the early pathway genes CYP71A12 and/or CYP79B2 are expressed indicate potential involvement of endogenous N. benthamiana enzymes.
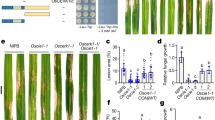
Extended Data Figure 8 ICN pathway metabolites contribute to disease resistance towards B. cinerea but not towards G. orontii.
a, Top: typical lactophenol trypan blue staining of leaves drop-inoculated with spores from the virulent fungal necrotroph B. cinerea to visualize the extent of host cell death (darkly stained areas within and beyond the fungal spore droplet region). Middle: microscopic analysis of stained leaves to visualize the extent of fungal colonization (stained filamentous fungal hyphae within and beyond the fungal spore droplet region). Images were taken at the same magnification (×25) and are representative of five biological replicates. Bottom: close-up images of the fungal hyphae beyond the fungal spore droplet region for cyp82C2 and cyp71A13 mutants. Images were taken at the same magnification (×100). b, Measurement of the disease lesion diameters in infected leaves. Data represent the median ± s.e.m. for five biological replicates. Asterisks denote statistical significance relative to WT (P < 0.05, two-tailed t test). c, Typical lactophenol trypan blue staining of fungal conidiophores (spore-bearing structures) formed in leaves infected with the adapted powdery mildew G. orontii. The pad4-1 mutant is more susceptible to fungal growth by G. orontii and thus produces significantly more conidiophores. Images were taken at the same magnification (×100) and are representative of three biological replicates. d, Measurement of the number of conidiophores in infected leaves. Data represent the mean ± s.d. for three biological replicates. e, Top: typical disease symptoms 3 days after drop inoculation of leaves with spores from the avirulent fungal necrotroph A. brassicicola. Bottom: microscopic analysis of infected leaves after lactophenol trypan blue staining confirming that disease symptoms are consistent with extent of fungal colonization (lightly stained fungal hyphae extending from the fungal spore droplet region) and host cell death (darkly stained areas along and beyond the border of the spore droplet region). Images were taken at the same magnification (×25) and are representative of ten biological replicates. f, Measurement of the disease lesion diameters in infected leaves. Data represent the median ± s.e.m. of eight (top graph) or ten biological replicates (bottom graph). Different letters denote statistically significant differences (P < 0.05, two-tailed t test).
Extended Data Figure 9 ICN and 4-OH-ICN but not their degradation products inhibit fungal growth in vitro.
a, b, Fungal growth inhibition assays on B. cinerea SF1 (a) or A. brassicicola FSU218 (b) with the tested compound (or compound combination) indicated. For compound combinations, the concentration indicated is for each compound; the given combinations approximate the hydrolysis products of ICN or 4-OH-ICN. Growth of fungi in potato dextrose broth on a microplate was quantified by measuring absorbance at 600 nm (OD600 nm) 72 h after spore inoculation and subtracting the absorbance at 0 h; see Methods for further details. Error bars, s.d. based on three biological replicates. Note that the half-maximum inhibitory concentrations (IC50) for both camalexin and ICN are approximately 25 μM against B. cinerea and 50 μM against A. brassicicola. For 4-OH-ICN, the inhibitory effect is not as pronounced, possibly because of rapid degradation of 4-OH-ICN in potato dextrose broth (see Supplementary Table 2).
Extended Data Figure 10 Levels of indolic compounds in leaves of mature plants after mock treatment or fungal infection.
Tissue extracts were analysed by LC–MS 7 days post-infection for A. brassicicola FSU218 and 5 days post-infection for B. cinerea SF1. a–e, Levels of indicated compound, quantified as EIC integral for the [M + H]+ ion (m/z ± 10 ppm) and converted to absolute amounts by comparison with a standard curve. f, Ion count integrals for indole glucosinolates ([M-H]− ion, m/z ± 10 ppm). Error bars in all panels, s.d. based on six biological replicates.
Supplementary information
Supplementary Information
This file contains Supplementary Text and Data and Supplementary References. (PDF 832 kb)
Supplementary Data
This file contains Supplementary Table 1. (XLSX 13 kb)
Supplementary Data
This file contains Supplementary Table 2. (XLSX 10 kb)
Supplementary Data
This file contains Supplementary Table 3. (XLSX 13 kb)
Rights and permissions
About this article

Cite this article
Rajniak, J., Barco, B., Clay, N. et al. A new cyanogenic metabolite in Arabidopsis required for inducible pathogen defence. Nature 525, 376–379 (2015). https://doi.org/10.1038/nature14907
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature14907
This article is cited by
-
The evolutionary trajectories of specialized metabolites towards antiviral defense system in plants
Molecular Horticulture (2024)
-
Xylobiose treatment triggers a defense-related response and alters cell wall composition
Plant Molecular Biology (2023)
-
Transcriptomic and genomic analysis provides new insights in molecular and genetic processes involved in zucchini ZYMV tolerance
BMC Genomics (2022)
-
Biosynthetic pathways of triterpenoids and strategies to improve their Biosynthetic Efficiency
Plant Growth Regulation (2022)
-
Early transcriptome changes induced by the Geminivirus C4 oncoprotein: setting the stage for oncogenesis
BMC Genomics (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
