
Abstract
Many arthropod species are infected by maternally inherited bacteria that induce cytoplasmic incompatibility (CI). CI causes embryonic mortality in offspring when infected males mate with either uninfected females or with females that are infected with a different strain of bacteria. Here, we review theoretical and empirical studies concerning the infection dynamics of CI-inducing bacteria, focusing in particular on the impact of the host population structure on the spread of CI. As different theoretical models have often produced divergent predictions with regard to issues such as the speed of CI spread and the stability of infection polymorphisms, we specifically aim to clarify how the various assumptions concerning population structure that underlie these models affect these predictions. We also discuss several implications of population structure, including the impact of CI on host gene flow reduction and speciation, the evolutionary dynamics of CI and strategies to control insect pest populations by means of CI-inducing microbes.
Similar content being viewed by others
Introduction
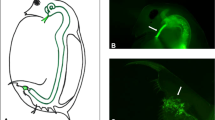
Marshall (1938) was apparently the first to note that crosses between certain strains of the mosquito Culex pipiens were incompatible in one direction (that is, males from strain A and females from strain B), whereas the other direction (B males × A females) produced viable progeny. This phenomenon was further investigated in the 1940s and 1950s by Roubaud (1941), Ghelelovitch (1952) and, in particular, Laven (1951, 1956, 1957, 1959). These early investigations showed that the incompatibility trait was inherited maternally (Laven, 1956), suggesting an extranuclear causative agent and giving rise to the term ‘cytoplasmic incompatibility’ (CI). In addition to unidirectional CI, bidirectional CI was also observed between strains of C. pipiens (Laven, 1951, 1959; Ghelelovitch, 1952). In the early 1970s, a connection was drawn by Yen and Barr (1971, 1973) between CI and the intracellular α-proteobacterium Wolbachia that had long been known to infect C. pipiens (Hertig and Wolbach, 1924): unidirectional CI usually occurs when infected males mate with uninfected females, but can also be found in crosses between individuals infected with different strains of bacteria (for example, Duron et al., 2006). In bidirectional CI, both directions of a cross are incompatible because of infection with different strains of CI-inducing microorganisms (Figure 1). In both cases, offspring from incompatible crosses suffer mortality at early stages of their development.

Illustration of uni- and bidirectional cytoplasmic incompatibility (CI). The two tables show success (green tick marks) or failure (red crosses) of offspring production of crosses between parents with different infection states. Empty symbols in the parent generation denote that these parents are uninfected, whereas the two shades of blue denote infection with two different strains of bacteria. With unidirectional CI, only crosses between infected males and uninfected females are incompatible. With bidirectional CI, crosses between males and females infected with different strains of CI-inducing bacteria are incompatible. Note that sometimes, unidirectional CI is also observed between hosts infected with different strains of bacteria.
Today, CI has been recorded in all major insect orders as well as in mites and woodlice (Table 1). CI is induced not only by Wolbachia but also by the unrelated bacterium Cardinium hertigii (Hunter et al., 2003; Gotoh et al., 2007a). Both bacteria are transmitted predominantly maternally through the egg cytoplasm, although paternal or horizontal transmission may also occur at a low rate (Turelli et al., 1992). In terms of evolutionary theory, CI can be understood in the light of intragenomic conflicts that arise from the maternal transmission of the bacteria (Cosmides and Tooby, 1981). As males are reproductive dead ends for cytoplasmic elements, it is advantageous to ‘exploit’ them to increase transmission through female hosts. CI is one of four strategies of such ‘reproductive parasitism’: males are exploited by the bacteria to kill the offspring of uninfected females so that infected females have a selective advantage (Werren, 1997). The other three phenotypes of reproductive parasitism (male killing, feminization and parthenogenesis induction) all involve sex-ratio distortion and are not discussed in this review.
CI is usually understood within a modification-rescue (or poison-antidote) framework (Werren, 1997): sperm is modified by the bacteria in males, and the same or a similar strain must be present in the eggs to ‘rescue’ the modification, enabling the progeny to develop normally. The precise nature of this modification-rescue mechanism is currently not understood. Cytological studies have shown that in incompatible crosses, the paternal chromosomes do not condense, are damaged and eventually lost during the first mitotic divisions (Breeuwer and Werren, 1990; Callaini et al., 1997; Tram and Sullivan, 2002). In diplodiploid and most haplodiploid species, the loss of paternal chromosomes leads to disrupted development and death of the embryos (Breeuwer, 1997; Callaini et al., 1997; Vavre et al., 2000). By contrast, in the haplodiploid wasp Nasonia vitripennis the haploid embryos survive and develop into males instead of females (Breeuwer and Werren, 1990). For excellent reviews on the mechanistic basis of CI, we refer to Poinsot et al. (2003) and Tram et al. (2003).
It is well known in the fields of evolutionary biology and epidemiology that the host population structure strongly affects the infection dynamics of diseases or symbionts as well as their evolutionary trajectories (for example, Herre, 1993; Keeling, 1999), and some sort of population structure certainly is a pervasive feature of most natural populations. However, models with population structure are often neglected. This is partly because of the mathematical difficulties that are involved in the corresponding models (that is, partial differential equations (PDEs) are usually more difficult to analyse than ordinary differential equations), owing in part to the lack of empirical data that would justify a model with a particular population structure.
In the case of CI-inducing microbes, there is an increasing number of field studies examining their spatial distribution. Maybe most impressively, the rapid spread of CI-inducing Wolbachia was observed in Californian populations of Drosophila simulans (Turelli and Hoffmann, 1991), and other studies also clearly show such spatial spread of CI-inducing microbes (Riegler and Stauffer, 2002; Hiroki et al., 2005). However, some surveys also suggest rather stable infection polymorphisms, with some populations being infected and others being uninfected or infected with a different bacterial strain (Merçot et al., 1995; Clancy and Hoffmann, 1996; Riegler and Stauffer, 2002; Baudry et al., 2003; Keller et al., 2004; Jaenike et al., 2006). Table 1 gives examples of species infected with CI-inducing microbes, indicating results concerning infection polymorphisms and population structure.
Here, we review the infection dynamics and evolution of CI-inducing microbes with a focus on host population structure. The various empirical observations have motivated several theoretical studies, and the main aim of this review is to clarify the assumptions that underlie these models and explain how they may sometimes lead to conflicting predictions. We start out by explaining some basic features of CI infection dynamics that emerge in simple models assuming single, panmictic populations of infinite size. This ‘null model’ of CI then forms the basis for a detailed discussion of more elaborate models that incorporate various forms of host population structure. In addition to infection dynamics, population structure also has an impact on evolutionary aspects of CI. We discuss recent theoretical advances concerning both the evolutionary consequences of CI-inducing microbes on their hosts (including gene-flow modification and potential facilitation of speciation) as well as the evolutionary dynamics of the microbes themselves. Finally, we review several CI-based strategies for insect pest control and evaluate their applicability in the light of the effects of host population structure.
The dynamics of CI in unstructured host populations
The null model of CI
Induction of CI can enable maternally inherited bacteria to spread in a host population. This is because if there are infected males in a population, infected females (which are compatible with all males) may on the average produce more daughters than uninfected females (which are compatible only with uninfected males). Thus, by decreasing the number of daughters that uninfected females produce, CI-inducing bacteria increase the relative daughter production of infected females and boost their own spread. This reasoning also shows a fundamental property of CI dynamics, the positively frequency-dependent selection for CI: the higher the infection frequency of the bacteria in the population, the stronger the selection for the bacteria. In what follows, we will describe a basic ‘null model’ for the infection dynamics of CI-inducing bacteria that serves as a starting point for models incorporating more complicated features of host biology, including population structure.
Three parameters have been established to be important in quantitatively predicting the spread of CI-inducing endosymbionts in a host population. First, the transmission rate gives the proportion of offspring produced by an infected mother that is also infected by the bacteria. Transmission rates are often close to one (for example, Rasgon and Scott, 2003; Narita et al., 2007; Perlman et al., 2008), but they may also be <95% (Poinsot et al., 2000). Second, the CI mortality (or CI level) is the proportion of offspring that dies in incompatible crosses. CI mortality varies widely (from zero to almost one) between different strains of bacteria (Drosophila strains reviewed in Merçot and Charlat, 2004) and it may also decline with the age of males (Clark et al., 2002; Reynolds and Hoffmann, 2002). Finally, CI-inducing bacteria may have direct effects on female fitness, necessitating a third parameter. For example, female fecundity can be decreased (for example, Hoffmann and Turelli, 1988; Perlman et al., 2008) or increased (Dobson et al., 2004) by the bacterial infection, but often, no effect on fecundity is seen (for example, Giordano et al., 1995; Bordenstein and Werren, 2000). It is noted that selection acting on the bacteria may also increase the fecundity of infected females over relatively short time spans and to the extent where infected females eventually have a higher fecundity than uninfected females (Weeks et al., 2007). As fecundity is the best-studied fitness component directly influenced by CI-inducing microbes, most theoretical models incorporate a parameter for fecundity; however, other fitness components may also be important and may require separate treatment in the models.
Several models for the infection dynamics of CI have been developed that incorporate some or all of these parameters (Caspari and Watson, 1959; Fine, 1978; Hoffmann et al., 1990; Hurst, 1991; Turelli, 1994). Discrete generations and random mating between hosts are generally assumed, and bacteria infecting a host are treated as a single genetic entity that can either be present or absent. In Box 1, we present a mathematical formulation and solutions of these models. The most important result is that because of the frequency-dependent selection, there may be an invasion threshold for CI, that is, an infection frequency below which the bacteria become extinct and above which they spread. This invasion threshold exists whenever infected females have (1) a reduced fecundity and/or (2) when transmission is imperfect (and this is not sufficiently compensated for by increased fecundity). Provided the invasion threshold is reached, the infection is expected to spread to a high polymorphic equilibrium frequency or—with perfect transmission or CI mortality—to fixation.
Population cage experiments with flies and mosquitoes have confirmed the qualitative predictions of the above models. Both Nigro and Prout (1990) and Dobson et al. (2002b) consistently observed an increasing infection frequency of CI-inducing Wolbachia over a few generations. In addition, Xi et al. (2005), using an artificially established Wolbachia infection in Aedes aegypti that led to a strong fecundity reduction of infected females, also reported the extinction of Wolbachia when it was introduced at a low initial infection frequency. This observation is in line with the invasion threshold of CI-inducing bacteria that have deleterious effects on their female hosts.
Extensions of the null model
Although the main focus of this review is the impact of population structure on the infection dynamics of CI, we will also give a short overview in this section of other factors that are expected to influence these dynamics.
It is currently not understood how the invasion threshold is overcome in natural populations, but stochastic effects (that is, random genetic drift) are likely to play an important role (Rousset and Raymond, 1991). Analytical approximations (Rigaud and Rousset, 1996; Jansen et al., 2008) and simulations (Egas et al., 2002; Jansen et al., 2008) indicate that the probability of CI spread starting from a singly infected female is generally rather low, particularly with large populations and high invasion thresholds. However, even in the face of an invasion threshold, the spread of CI was often found to be more likely than that of a comparable neutral genetic element, suggesting that repeated exposure of a population to CI-inducing bacteria through horizontal transmission may produce a reasonable likelihood of eventual invasion (Jansen et al., 2008).
When a single panmictic population is infected with more than one strain of bacteria that induce bidirectional CI, it is crucial to distinguish whether multiple infections occur at the level of individual hosts or not. In the latter case, modelling predicts that one of the strains will eventually drive the other(s) extinct, and which of the strains prevails will depend both on the parameters and on the initial frequencies of each strain (Rousset et al., 1991). When there are multiply infected females in the population, the population may remain polymorphic (Frank, 1998). More precisely, a stable polymorphism of several CI-inducing strains can only be maintained in a panmictic population if a subset of the population is infected with all of these strains. This is because only females infected with all strains of bacteria are compatible with all males and therefore enjoy the highest fitness in the population. Female hosts carrying only a subset of bacterial strains have lower fitness but may be maintained in the population through imperfect maternal transmission.
Other complications of CI concern haplodiploid species. First, males develop from unfertilized eggs in haplodiploid species, and so that their production is not affected by incompatible matings. Second, as CI involves a loss of the paternal set of chromosomes, diploid zygotes arising from incompatible matings may survive as haploid males. This type of CI has been observed in the wasp N. vitripennis (Breeuwer and Werren, 1990) and is termed the male development (MD) type of CI. Most haplodiploid species, however, seem to show the female mortality (FM) type of CI, in which fertilized eggs from incompatible crosses fail to hatch as in diplodiploid species (for example, Breeuwer, 1997; Vavre et al., 2000). It has been shown theoretically that all else being equal, the invasion threshold for CI is highest for the MD type of CI, followed by the FM type and the diplodiploid type of CI (Vavre et al., 2000). Conversely, the stable equilibrium frequency is highest in diplodiploid species and lowest with the MD type of CI in haplodiploid species.
Finally, infection dynamics may be influenced by a number of other factors that have been studied theoretically, but which we can only briefly mention here. These factors include paternal transmission of the bacteria (Nigro and Prout, 1990), host nuclear genes suppressing CI action (Rousset et al., 1991; Turelli, 1994; Koehnke et al., 2009), competition within a brood of siblings (Freeland and McCabe, 1997), the presence of other maternally inherited bacteria that induce a sex-ratio distortion (Freeland and McCabe, 1997; Engelstädter et al., 2004), age-structured host populations (Rasgon and Scott, 2004) and sperm competition (Hoffmann and Turelli, 1997; Champion de Crespigny et al., 2008).
The dynamics of CI in structured host populations
Roughly speaking, three different types of models with host population structure have been constructed, which are illustrated in Figure 2. These types of models are (1) deterministic models of continuously structured (‘viscous’) populations, (2) deterministic models with discrete subpopulations and (3) stochastic models with discrete subpopulations (‘deme-structured’ populations). We will discuss these different classes of models in turn and aim at clarifying how they produce different outcomes and sometimes even opposing predictions.

Illustration of three types of approaches to model population structure. In each model, red and black colours represent different infection states of individuals, for example, infected and uninfected. (a) Models with continuous spatial distribution of individuals, employing partial differential equations. (b) Models with migration between two infinitely large subpopulations. Migration may occur in both directions (two-islands model) or in one direction only (mainland–island model). (c) Models with a number of demes of small to intermediate size. Migration may occur between all demes with equal probability (Wright's island model) or in a geographically restricted way (for example, stepping-stone model).
Before entering this discussion, however, we would like to briefly examine how inbreeding and outbreeding influence the spread of CI. Inbreeding and outbreeding are deviations from panmixis that may result from spatial subdivision of populations, but may also represent a form of population structure that only extends to mating (for example, through behavioural inbreeding avoidance) without actual spatial subdivision. To date, a systematic investigation of the impact of inbreeding and outbreeding on CI dynamics is still lacking. Numerical analyses of a model with sibmating or sibmating avoidance indicate that substantial inbreeding can increase the invasion threshold and decrease the stable equilibrium frequency of the CI-inducing microbes (Engelstädter et al., 2006). This is because with increasing levels of inbreeding, incompatible matings become increasingly unlikely, thus diminishing selection for the CI microbes. When the level of inbreeding is above a certain threshold, CI invasion and persistence can even become impossible. Sibmating avoidance (outbreeding), on the other hand, did not seem to have an effect on the infection dynamics (Engelstädter et al., 2006). This is because even when females never mate with their brothers, this hardly affects the probability of mating with an uninfected or infected male when the population is reasonably large.
Continuous ‘viscous’ populations
This class of models was originally motivated by the observation of an ongoing spread of CI-inducing Wolbachia through a Californian D. simulans population, documented in detail by Turelli, Hoffmann and colleagues (Hoffmann et al., 1986; Hoffmann and Turelli, 1988; Turelli and Hoffmann, 1991; Turelli et al., 1992). The spread was described mathematically using a PDE that was originally developed for the analysis of hybrid zones (Barton, 1979; Turelli and Hoffmann, 1991). To test the model, the authors measured the three main parameters for the CI dynamics (transmission rate, CI level, fecundity), and combined them in the model with rather phenomenological assumptions about the underlying insect dispersal mode (Turelli and Hoffmann, 1991). As a result, the model describes the spread of Wolbachia qualitatively well but fails to make accurate predictions about the invasion speed. As the authors acknowledge, this is most likely because of the lack of knowledge about the actual range of migration in D. simulans.
In a more recent theoretical study, Schofield (2002) investigated the role of insect dispersal on the spatiotemporal spread of unidirectional CI. The underlying PDEs are based on Turelli and Hoffmann (1991). Two types of PDE were analysed: reaction-diffusion and integro-difference equations. These models differ with respect to the mode of migration. In the former, dispersal is treated as a diffusion process and dispersal distance modelled by a normal distribution, whereas the latter uses a leptokurtic distribution that allows for rare long-range dispersal of hosts. Model analysis was carried out by computer simulations, and parameters were chosen to match the D. simulans system. The main result of this study was that long-range migration results in faster spread of CI and that including such dispersal provided a better description of CI spread in D. simulans. The merit of Schofield's study is to point out that the actual mode of insect dispersal plays a crucial role in the speed of spread as well as the shape of the infection wave. However, as with most theoretical studies based purely on computer simulations, the lack of analytical results and the limited range of parameters for which simulations were performed make it difficult to generalize the results.
Given the close relationship between models of hybrid zones and those of CI in continuous populations, Barton's classical study (Barton, 1979) may provide clues on factors that can be expected to affect the spatial spread of symbionts. The equations analysed by Barton (1979) model hybrid zones caused by underdominant chromosomal rearrangements. From a mathematical point of view, an underdominant locus is almost equivalent to symmetric bidirectional CI (that is, the degree of CI is equal in both directions). It might therefore be worth investigating to what extent previous predictions about hybrid zones, including results on spatial heterogeneity, hold true for bidirectional CI. In particular, the theoretical analysis of hybrid zones indicates that spatial heterogeneity in population density plays a crucial role in wave speed and cline stability; regions with low population density are obstacles that can prevent the spread and lead, under certain circumstances, to stable clines. An extension of Schofield's model could reveal a similar effect; the spread of a CI-inducing microbe might be impeded or prevented in regions with low population density. Stochastic effects, however, might lead to an opposite effect. This is because with low population density, random genetic drift becomes more important, which was shown to promote the spread of unidirectional CI (Reuter et al., 2008; also see discussion below).
We close this subsection with a general remark on spatial models. In PDEs, space is considered as a continuous variable, whereas in lattice or stepping-stone models, space consists of discrete units. Both types of models are often too complicated for analytical analysis, making the use of numerical methods necessary. When interpreting numerical results, however, it is important to recall that computers, in general, allow only discrete mathematical calculations; computational methods for solving PDEs do not generate solutions of the PDE, but only give approximations of such solutions. The results presented by Schofield (2002) are therefore, in a strict mathematical sense, the solutions of a stepping-stone model with a very high number of subpopulations. This should be kept in mind when comparing studies based on discrete models with those based on PDEs.
Deterministic models with discrete subpopulations
We next consider deterministic models with a discrete number of subpopulations that are connected by migration. The main question underlying these models is under which conditions coexistence of differentially infected subpopulations is possible. We discuss the following three models in detail: (1) unidirectional CI in a two-population model (Flor et al., 2007, 2) bidirectional CI in a two-population model (Telschow et al., 2005b) and (3) bidirectional CI in a lattice model (Keeling et al., 2003). The basic idea behind all three studies is the same (and goes back to Nigro and Prout (1990) and Rousset et al. (1991)): frequency-dependent selection on CI creates a situation where bistability does not allow stable coexistence in a panmictic population but does allow coexistence in different subpopulations if migration is below a critical value.
The dynamics of bacterial infections causing unidirectional CI in a two-population model was investigated by Flor et al. (2007). Infected and (mainly) uninfected host populations can coexist stably in this model if migration is below a critical value. The critical migration rates were determined analytically for mainland–island models and it was shown that these solutions are lower-bound estimates for critical migration rates in the general model with two-way migration. In general, critical migration rates exist if either the bacteria cause fecundity reductions in infected female hosts or bacterial transmission is imperfect. In the case of symmetric two-way migration, infection polymorphism is most stable for intermediate levels of CI.
A similar analysis was carried out for two bacterial strains causing bidirectional CI (Telschow et al., 2005b). It was shown analytically for a two-population model that the stability could be fully described in terms of the critical migration rate, which is here defined as the highest migration rate below which a stable coexistence of the two bacterial strains is possible. The main finding of the study is that bidirectional CI can persist stably in the face of high migration rates. For example, CI in Culex and Nasonia is nearly complete (Perrot-Minnot et al., 1996; Rasgon and Scott, 2003), resulting in a critical migration rate of 19% per generation, whereas CI in Drosophila is more variable, ranging from 0.3 to 0.7 (Merçot and Charlat, 2004), which causes critical migration rates between 5 and 13%. These values are roughly one order of magnitude higher than for unidirectional CI, indicating that bidirectional CI may result in substantially more stable infection polymorphisms.
The studies of Flor et al. (2007) and Telschow et al. (2005b) do not take into account host nuclear background. We mention in this context that local adaptation in hosts significantly increases the stability of infection polymorphism and critical migration rates (Telschow et al., 2002a, 2002b, 2007). Moreover, it was shown that other reproductive parasites (male-killing bacteria) can prevent the spread of CI to a certain extent, thereby increasing the respective critical migration rates (Engelstädter et al., 2008).
Finally, Keeling et al. (2003) investigated the coexistence of two bacterial strains causing bidirectional CI in a (one- or two-dimensional) lattice model. The authors used a purely numerical approach, and in contrast to the above-discussed models, infection dynamics in each subpopulation were described by ordinary differential equations reflecting overlapping host generations. The main result of Keeling et al. (2003) is that stable coexistence of the two strains is possible if migration is below a threshold. This is in line with the above-discussed critical migration rates. Coexistence of strains comes about through the formation of different patches within the entire population that are occupied predominantly by one or the other strain. In addition, the study points out the role of founder control in determining the eventual infection status of a subpopulation.
Stochastic models of deme-structured populations
The final class of models that we discuss assumes that the host population is subdivided into a number of ‘demes’ of finite size, thus incorporating random genetic drift into the models. A first analysis of this type of model was presented by Wade and Stevens (1994). Here, it was assumed that mating takes place within demes, but all offspring disperse and compete at the level of the entire population before again forming demes for mating. Thus, this model assumes hard selection, that is, complete population mixture and global competition in each generation. It was shown that this scenario slows the spread of CI-inducing microbes. This result can be understood by examining the effect of the model setup on the frequency of incompatible matings, which determines the selective advantage of CI. In a panmictic population, incompatible matings occur with a probability equal to the variance in infection status (see also Box 1). In the model by Wade and Stevens (1994), the variance in infection status in the entire population is partitioned into the variance in infection frequency between demes and the average variance of infection status within demes. As the latter determines the overall mortality, it can be seen that the higher the variance in infection frequency across demes, the fewer the offspring that will be killed, so that selection for the CI microbes is reduced.
A recent, more explicit analysis has come to rather different conclusions (Reuter et al., 2008). In this model, competition occurs locally and only a fraction of the offspring disperses from their natal deme. Using a combination of analytical methods and simulations, the authors showed that the deme structure can substantially facilitate the spread of CI compared with a panmictic population. Specifically, it was shown that the spread of CI is most likely when migration rates are low and demes are of intermediate size. Under these conditions, the amount of local random drift maximizes the frequency of incompatible matings and is therefore most conducive to the spread of the CI microbes. The frequency of incompatible matings is lower (and conditions less favourable for CI) when demes are very small or large, because then random genetic drift is either so strong that it homogenizes infection within demes or so weak that it does not allow for stochastic increases in infection frequency within demes, leading to essentially deterministic dynamics. These results appear to hold qualitatively both with a migrant pool and a stepping-stone type of migration model. Furthermore, the investigation of sex-specific dispersal rates revealed that the likelihood of CI spread was mainly determined by the female dispersal rate, but that it did also decrease with increasing male dispersal rate.
The divergent predictions of the two models described—impeded or facilitated spread of CI through population subdivision in Wade and Stevens (1994) and Reuter et al. (2008), respectively—indicate that the selection regime applied is decisive for the impact of population structure on the spread of CI: global competition combined with complete mixing of the population disfavours the spread of CI-inducing microbes. On the other hand, local competition appears to generally favour the spread of CI, although this effect is strong only with low migration rates and intermediate deme size. This latter result may appear puzzling on first sight when compared with the results concerning the critical migration rate discussed in the previous section. Here, low migration rates impeded the spread of CI microbes into an adjacent population. It is important to realize that not only is the underlying model different (it assumes infinitely large subpopulations without random genetic drift), but also the questions addressed. Whilst the deme-structured models aim to study the likelihood of CI spread and its expected velocity in populations with a more ‘local’ structure, the deterministic models investigate under what conditions a stable polymorphism can be maintained in populations structured on a larger geographical scale. The deterministic models assume infinitely large subpopulations and, consequently, CI spread relies on high migration rates to overcome the invasion threshold. In the deme-structured models, by contrast, spread relies on stochastic increases of infection frequency above the invasion threshold, which requires low migration (little dilution) and genetic drift.
Implications of population structure
Host gene flow
It is a well-known result in population genetics that genetic influx into a population often does not correspond to the real migration rate (m) of individuals, owing to some class structure within the population. Examples of class structure include different sexes, allelic classes, age or infection status. The concept of effective migration rate (me) was developed to measure gene flow in such situations (Barton and Bengtsson, 1986; Kobayashi et al., 2008). The ratio of the effective migration rate to the real migration rate (me/m) is called gene flow factor and represents the degree of gene flow modification. In a class-structured population with mainland–island population structure, the gene flow factor is in good approximation equal to the mean reproductive value of immigrants (Kobayashi et al., 2008).
The concept of effective migration rate allows the determination of the impact of CI on host gene flow (Telschow et al., 2002a, 2002b, 2007; Engelstädter et al., 2008). It was shown that both uni- and bidirectional CI causes gene flow reduction between host populations that differ in their infection status. Using the methods described in Kobayashi and Telschow (2008), the gene flow factor can be calculated analytically as

In this formula, lA and lA are the mortality rates in incompatibility matings where males are infected with bacterial strains A and B, respectively, and we consider gene flow from a (mainland) population infected with strain A to an (island) population mainly infected with strain B. Formula (1) shows that CI can cause strong gene flow reduction between populations with different infection state. For example, if half of the offspring are killed through CI (lA=lB=0.5), gene flow is reduced by two-third (me/m≈1/3), and in the extreme case of complete CI (lA=lB=1), gene flow is reduced to zero, me/m≈0.
By setting lB=0, formula (1) can also be applied to unidirectional CI. An interesting implication of formula (1) then is that unidirectional CI causes an asymmetrical gene flow reduction: gene flow from an uninfected population to an infected population is reduced by 1/(1+lA),whereas gene flow in the opposite direction is reduced by 1−lA. As 1/(1+lA)>1−lA for lA>0, this shows that gene flow is less strongly reduced towards the CI-infected population than towards the uninfected population. Thus, CI converts infected populations into population genetic ‘sinks’ (that is, larger genetic influx than outflux), whereas their neighbouring uninfected populations are converted into genetic ‘sources’. This might have important implications for the evolutionary process of both host and symbiont.
Premating isolation and speciation
The idea that CI could facilitate speciation of hosts is nearly as old as CI's discovery (Laven, 1959), and has generated some controversy over the last few decades (Hurst and Schilthuizen, 1998; Werren, 1998; Bordenstein et al., 2001; Wade, 2001; Telschow et al., 2007). The role of CI would be to reduce gene flow between populations, allowing genetic divergence for locally adaptive traits, and to select for premating isolation. Above, we have summarized the effect of CI on host gene flow. Here, we discuss how population structure creates circumstances for CI to select for female mating preferences. We constrain ourselves to discussing theoretical results and refer to the review by Bordenstein (2003) for a detailed discussion of empirical patterns.
To date, three different models exist that investigate how CI could select for premating isolation: (1) unidirectional CI in a panmictic population (Champion de Crespigny et al., 2005, 2) unidirectional CI in a mainland–island model (Telschow et al., 2007) and (3) bidirectional CI in a two-population model (Telschow et al., 2005a). The basic idea is the same in all three scenarios: mating preference is adaptive when it reduces the risk of getting involved in an incompatibility mating.
Champion de Crespigny et al. (2005) considered a panmictic host population to investigate the evolution of female mating preference in the presence of unidirectional CI. Their model shows that during the spread of CI, a mutant allele that leads female carriers to preferentially mate with uninfected males can increase in frequency. However, this study may be criticized on two grounds. First, the results depend crucially on the assumption that females can choose between infected and uninfected males. This is, to our knowledge, not supported by any empirical data. Second, the increase in mutant allele frequency was observed only during the spread of CI, but not after CI has spread to fixation. Given that the spread of CI-inducing bacteria is potentially very rapid, this obviously limits the range under which CI can select for mating preference. Both critics raise serious doubts whether unidirectional CI can select for female mating preference in a panmictic host population.
A different picture emerges when population structure is considered. As reviewed above, stable infection polymorphisms are possible for both uni- and bidirectional CI if migration is below a critical rate (Telschow et al., 2005b; Flor et al., 2007). These infection polymorphisms create situations where females face an ongoing threat of being involved in incompatibility matings. Combining numerical and analytical approaches, it was shown that this allows mating preference to evolve under a broad range of circumstances (Telschow et al., 2005a, 2007). Both models for uni- and bidirectional CI differ from the panmictic model of Champion de Crespigny et al. (2005) in that they do not assume mating preference for uninfected males (or infection with a certain bacterial strain). Instead, mating preference is for locally adapted male traits, and mating preference alleles spread because the male traits tend to be associated with the respective infection states. In summary, population structure solves both criticisms that were raised against the panmictic model; it allows a stable infection polymorphism as well as a polymorphism of locally adapted male traits that females may use as a preference cue.
Evolutionary dynamics of CI
How new CI types evolve and how CI arose in the first place is currently not understood, in part because the mechanistic and genetic bases of CI remain elusive. Assuming that the modification (mod) and rescue (resc) aspects of CI are genetically independent (for empirical justification of this assumption, see Poinsot et al., 2003), a crucial feature of CI is that in panmictic populations, there is no selection on mod. This means that, provided they still retain the original resc function, mutant strains of bacteria that cannot induce modification in males or induce a different mod are selectively neutral relative to the ancestral strain (Prout, 1994; Charlat et al., 2001). Therefore, through random genetic drift, the bacteria may lose their ability to induce CI and become extinct (Turelli, 1994; Hurst and McVean, 1996), or evolve a new CI type that is bidirectionally incompatible to the ancestral CI type (Charlat et al., 2001, 2005a).
The neutrality of the mod function rests on the assumption of random mating and an unstructured host population. With inbreeding, mod or new mod variants are selected against when there is no resc against the new mod, whereas outbreeding (inbreeding avoidance) favours new variants of mod that cannot be rescued (Engelstädter and Charlat, 2006; Engelstädter et al., 2006). On the other hand, however, selection for mod is not influenced by in- or outbreeding if resc is always complete (Haygood and Turelli, 2009). Moreover, local relatedness between individuals is expected to also lead to positive selection for mod (Frank, 1997; Hurst, 1991). This is because bacteria that induce CI in males will relax local competition between hosts and thereby benefit their relatives in females. Given that the mating patterns, the scale of competition, local relatedness and random genetic drift are all expected to be important factors, it seems clear that host population structure—potentially having an impact on all of these factors—plays a key role in the evolutionary dynamics of CI.
A recent study investigating the impact of population structure on the evolutionary dynamics of CI found that selection for increased CI level was present with local density-regulation (as predicted by Hurst (1991) and Frank (1997)), but this effect was only weak and transient (Haygood and Turelli, 2009). This is because relatedness was not, as in Frank's (1997) treatment, a fixed parameter, but a variable quantity that converges quickly to zero as the population becomes homogenized through migration. The analysis by Haygood and Turelli thus indicates that the evolutionary maintenance of the mod function remains a conundrum even in subdivided populations, leading the authors to subscribe to the notion that CI persists in the long term only through clade selection (Hurst and McVean, 1996). According to this view, CI persists in the long term because clades of microorganisms that retain the ability to induce strong CI have a higher potential to spread in novel host species than do clades with low CI levels. In principle, this mechanism could also work with different populations of the same species (with migration playing the role of interspecies transmission), but migration rates need to be very low to prevent fast homogenization of the populations. Clade selection for CI as well as the evolution of new types of CI (that is, new mod/resc pairs) in structured host populations remains to be analysed.
CI and insect pest control
Many medically important insect species, including the disease-transmitting mosquitoes C. pipiens and A. aegypti, are infected with or can be artificially infected with CI-inducing Wolbachia. Therefore, CI has been investigated as a mechanism to control insect pest populations (Laven, 1967; Zabalou et al., 2004), or to drive transgenes into insect populations (Turelli and Hoffmann, 1999; Sinkins and Godfray, 2004). In what follows, we will give a brief overview of these strategies (for in-depth reviews, see Sinkins and Gould, 2006; Aksoy, 2008) and discuss how their applicability may be dependent on host population structure.
The most straightforward method to employ CI as a control of insect pests is to release large quantities of infected males into an uninfected population. These males will then result in increased mortality among offspring, potentially reducing the population size of the next generation. This strategy has been successfully applied in a field study of a mosquito (C. pipiens) population (Laven, 1967) and also in population cage experiments with the medfly Ceratitis capitata (Zabalou et al., 2004). As this method is largely independent from the infection dynamics of CI, we expect the host population structure to be important only insofar as it restricts dispersal of the released males.
The main disadvantage of this scheme of CI-based pest control is that large quantities of infected males need to be produced. These males need to be sexed accurately so that no infected females are accidentally released into the target population, in which case the infection might spread so that a further release of infected males has no effect. The second strategy for reducing the host population size by means of CI that circumvents these problems is the CI management (CIM) strategy (Dobson et al., 2002a). CIM consists of the repeated release of infected individuals (males and females), combined with a monitoring of the population for infection frequencies. During the spread of the infection, the host population may be reduced because of incompatible matings (Dobson et al., 2002a). In addition, the targeted release of other bidirectionally incompatible strains at appropriate times is expected to further suppress the host population (Dobson et al., 2002a). Finally, insect species that act as disease vectors may be controlled through CI-inducing bacteria that reduce the life span of their hosts, because older insects are often more likely to transmit the disease than younger ones. An example is the mosquito A. aegypti, which has recently been successfully transinfected with a life-span reducing strain of Wolbachia (McMeniman et al., 2009).
The theoretical results discussed in the previous sections suggest that the success of CIM may be strongly determined by the host population structure. For example, as population structure influences the speed of CI spread, this in turn determines how transient suppression of the host population is and in which time intervals new CI strains need to be released. More importantly, the release of bidirectionally incompatible CI strains into a structured host population may also lead to the establishment of stable infections in the various subpopulations, so that there may be only few incompatible matings and consequently little population suppression. Further modelling investigating the impact of CI on host population dynamics in structured populations is needed to assess the applicability of CIM in such populations.
As an alternative to reducing their population size, insect pests (in particular, disease vectors) may be controlled by driving desired genes into these populations (reviewed in Sinkins and Gould, 2006). For example, such genes could reduce the ability of mosquitoes to transmit Plasmodium parasites. CI-inducing Wolbachia have been proposed as a candidate drive mechanism by means of which genes can spread into the target population that either have been transgenically incorporated into the Wolbachia genome or that are linked to Wolbachia (Curtis, 1994; Sinkins et al., 1997; Turelli and Hoffmann, 1999). Again, we expect the applicability and efficiency of this strategy to depend on the host population structure, which may accelerate, slow or even prevent the spread of the transgenes along with the CI-inducing bacteria. It has also been suggested that actively influencing the population structure may be a promising strategy for increasing the probability of CI spread (Reuter et al., 2008): as moderately small deme sizes and low rates of migration favour the spread of CI, the release of few transgenic hosts combined with the restriction of migration can create an infection base from which further spread through the entire host population is facilitated.
Conclusions
Given the importance of population structure in the fields of epidemiology, population genetics and evolutionary biology, it is not surprising that population structure is expected to have a large impact on the dynamics of CI spread. Mathematical models with different assumptions and addressing different questions have produced a variety of predictions concerning the probability and velocity of CI spread as well as the stability of spatial polymorphisms. Depending on the scale of competition in the population, CI spread can be facilitated (with local competition) or impeded (with global dispersion and competition). Infection polymorphisms of two bidirectional CI-inducing strains can be very stable in structured host populations, whereas polymorphisms of infected and uninfected populations are stable only when migration rates are very low. Given the disparity and the sometimes diametrically opposed predictions of previous models, future models providing a unifying framework and producing all of the above outcomes with different parameters might be desirable.
On the empirical side, virtually all theoretical predictions concerning CI spread in structured populations remain untested. Data from surveys of natural populations indicate that both stable polymorphisms and spatial spread are common, but as of date, these data have rarely been linked quantitatively to model predictions. This is largely because of inherent difficulties in estimating demographic parameters such as migration rates and population sizes. Population cage experiments that emulate different kinds of population structure may be a more feasible alternative to get a better understanding of how much and what aspects of reality are captured by the various theoretical models. Such an understanding appears particularly important if one is to apply CI-inducing microbes to controlling insect pest populations.
Change history
24 November 2020
A Correction to this paper has been published: https://doi.org/10.1038/s41437-020-00391-9
References
Aksoy S (ed) (2008). Transgenesis and the Management of Vector-Borne Disease. Springer: Berlin.
Barton NH (1979). Dynamics of hybrid zones. Heredity 43: 341–359.
Barton NH, Bengtsson BO (1986). The barrier to genetic exchange between hybridising populations. Heredity 56: 357–376.
Baudry E, Bartos J, Emerson K, Whitworth T, Werren JH (2003). Wolbachia and genetic variability in the birdnest blowfly Protocalliphora sialia. Mol Ecol 12: 1843–1854.
Bordenstein SR (2003). Symbiosis and the origin of species. In: Bourtzis K, Miller TA (eds). Insect Symbiosis. CRC Press: Boca Raton. pp 283–304.
Bordenstein SR, O'Hara FP, Werren JH (2001). Wolbachia-induced incompatibility precedes other hybrid incompatibilities in Nasonia. Nature 409: 707–710.
Bordenstein SR, Werren JH (1998). Effects of A and B Wolbachia and host genotype on interspecies cytoplasmic incompatibility in Nasonia. Genetics 148: 1833–1844.
Bordenstein SR, Werren JH (2000). Do Wolbachia influence fecundity in Nasonia vitripennis? Heredity 84: 54–62.
Breeuwer JAJ (1997). Wolbachia and Cytoplasmic incompatibility in the spider mites Tetranychus urticae and T. turkestani. Heredity 79: 41–47.
Breeuwer JAJ, Werren JH (1990). Microorganisms associated with chromosome destruction and reproductive isolation between two insect species. Nature 346: 558–560.
Callaini G, Dallai R, Riparbelli MG (1997). Wolbachia-induced delay of paternal chromatin condensation does not prevent maternal chromosomes from entering anaphase in incompatible crosses of Drosophila simulans. J Cell Sci 110: 271–280.
Caspari E, Watson GS (1959). On the evolutionary importance of cytoplasmic sterility in mosquitoes. Evolution 13: 568–570.
Champion de Crespigny FE, Butlin R, Wedell N (2005). Can cytoplasmic incompatibility inducing Wolbachia promote the evolution of mate preferences? J Evol Biol 18: 967–977.
Champion de Crespigny FE, Hurst LD, Wedell N (2008). Do Wolbachia-associated incompatibilities promote polyandry? Evolution 62: 107–122.
Charlat S, Calmet C, Andrieu O, Merçot H (2005a). Exploring the evolution of Wolbchia compatibility types: a simulation approach. Genetics 170: 495–507.
Charlat S, Calmet C, Merçot H (2001). On the mod resc model and the evolution of Wolbachia compatibility types. Genetics 159: 1415–1422.
Charlat S, Engelstädter J, Dyson EA, Hornett EA, Duplouy AMR, Tortosa P et al. (2006). Competing selfish genetic elements in the butterfly Hypolimnas bolina. Curr Biol 16: 2453–2458.
Charlat S, Hornett EA, Dyson EA, Ho PPY, Loc NT, Schilthuizen M et al. (2005b). Is extreme male-killer prevalence a local or common event in the butterfly Hypolimnas bolina? A survey across Indo-Pacific populations. Mol Ecol 14: 3525–3530.
Clancy DJ, Hoffmann AA (1996). Cytoplasmic Incompatibility in Drosophila simulans: evolving complexity. Trends Ecol Evol 11: 145–146.
Clark ME, Veneti Z, Bourtzis K, Karr TL (2002). The distribution and proliferation of the intracellular bacteria Wolbachia during spermatogenesis in Drosophila. Mech Dev 111: 3–15.
Cosmides LM, Tooby J (1981). Cytoplasmic inheritance and intragenomic conflict. J Theor Biol 89: 83–129.
Curtis CF (1994). The case for malaria control by genetic manipulation of its vectors. Parasitol Today 10: 371–374.
Dobson SL, Fox CK, Jiggins FM (2002a). The effect of Wolbachia-induced cytoplasmic incompatibility on host population size in natural and manipulated systems. Proc R Soc London Ser B 269: 437–445.
Dobson SL, Marsland EJ, Rattanadechakul W (2002b). Mutualistic Wolbachia infection in Aedes albopictus: accelerating cytoplasmic drive. Genetics 160: 1087–1094.
Dobson SL, Rattanadechakul W, Marsland EJ (2004). Fitness advantage and cytoplasmic incompatibility in Wolbachia single- and superinfected Aedes albopictus. Heredity 93: 135–142.
Duron O, Bernard C, Unal S, Berthomieu A, Berticat C, Weill M (2006). Tracking factors modulating cytoplasmic incompatibilities in the mosquito Culex pipiens. Mol Ecol 15: 3061–3071.
Egas M, Vala F, Breeuwer JAJ (2002). On the Evolution of Cytoplasmic Incompatibility in Haplodiploid Species. Evolution 56: 1101–1109.
Engelstädter J, Charlat S (2006). Outbreeding selects for spiteful cytoplasmic elements. Proc R Soc London Ser B 273: 923–929.
Engelstädter J, Charlat S, Pomiankowski A, Hurst GDD (2006). The evolution of cytoplasmic incompatibility types: integrating segregation, inbreeding and outbreeding. Genetics 172: 2601–2611.
Engelstädter J, Telschow A, Hammerstein P (2004). Infection dynamics of different Wolbachia-types within one host population. J Theor Biol 231: 345–355.
Engelstädter J, Telschow A, Yamamura N (2008). Coexistence of cytoplasmic incompatibility and male-killing-inducing endosymbionts, and their impact on host gene flow. Theor Popul Biol 73: 125–133.
Fine PEM (1978). On the dynamics of symbiote-dependent cytoplasmic incompatibility in culicine mosquitoes. J Invertebr Pathol 30: 10–18.
Flor M, Hammerstein P, Telschow A (2007). Dynamics and stability of Wolbachia-induced unidirectional cytoplasmic incompatibility in parapatric host populations. J Evol Biol 20: 696–706.
Frank SA (1997). Cytoplasmic incompatibility and population structure. J Theor Biol 184: 327–330.
Frank SA (1998). Dynamics of cytoplasmic incompatability with multiple Wolbachia infections. J Theor Biol 192: 213–218.
Freeland SJ, McCabe BK (1997). Fitness compensation and the evolution of selfish cytoplasmic elements. Heredity 78: 391–402.
Ghelelovitch S (1952). Sur le déterminisme génétique de la sterilité dans le croisement entre differentes souches de Culex autogenicus Roubaud. CR Acad Sci Paris 24: 2386–2388.
Giordano R, O'Neill SL, Robertson HM (1995). Wolbachia infections and the expression of cytoplasmic incompatibility in Drosophila sechellia and D. mauritiana. Genetics 140: 1307–1317.
Gotoh T, Noda H, Ito S (2007a). Cardinium symbionts cause cytoplasmic incompatibility in spider mites. Heredity 98: 13–20.
Gotoh T, Sugasawa J, Noda H, Kitashima Y (2007b). Wolbachia-induced cytoplasmic incompatibility in Japanese populations of Tetranychus urticae (Acari: Tetranychidae). Exp Appl Acarol 42: 1–16.
Haygood R, Turelli M (2009). Evolution of incompatibility-inducing microbes in subdivided host populations. Evolution 63: 432–447.
Herre EA (1993). Population structure and the evolution of virulence in nematode parasites of fig wasps. Science 259: 1442–1445.
Hertig M, Wolbach SB (1924). Studies on Rickettsia-like organisms in insects. J Med Res 44: 329–374.
Hiroki M, Ishii Y, Kato Y (2005). Variation in the prevalence of cytoplasmic incompatibility-inducing Wolbachia in the butterfly Eurema hecabe across the Japanese archipelago. Evol Ecol Res 7: 931–942.
Hiroki M, Tagami Y, Miura K, Kato Y (2004). Multiple infection with Wolbachia inducing different reproductive manipulations in the butterfly Eurema hecabe. Proc R Soc London Ser B 271: 1751–1755.
Hoffmann AA, Turelli M (1988). Unidirectional incompatibility in Drosophila simulans: inheritance, geographic variation and fitness effects. Genetics 119: 435–444.
Hoffmann AA, Turelli M, Harshman LG (1990). Factors affecting the distribution of cytoplasmic incompatibility in Drosophila simulans. Genetics 126: 933–948.
Hoffmann AA, Turelli M (1997). Cytoplasmic incompatibility in insects. In: O'Neill SL, Hoffmann AA, Werren JH (eds). Influential Passengers: Inherited Microorganisms and Arthropod Reproduction. Oxford University Press: New York. pp 41–80.
Hoffmann AA, Turelli M, Simmons GM (1986). Unidirectional incompatibility between populations of Drosophila simulans. Evolution 40: 692–701.
Hornett EA, Duplouy AMR, Davies N, Roderick GK, Wedell N, Hurst GDD et al. (2008). You can't keep a good parasite down: evolution of a male-killer suppressor uncovers cytoplasmic incompatibility. Evolution 62: 1258–1263.
Hunter MS, Perlman SJ, Kelly SE (2003). A bacterial symbiont in the Bacteroidetes induces cytoplasmic incompatibility in the parasitoid wasp Encarsia pergandiella. Proc R Soc London Ser B 270: 2185–2190.
Hurst GDD, Schilthuizen M (1998). Selfish genetic elements and speciation. Heredity 80: 2–8.
Hurst LD (1991). The evolution of cytoplasmic incompatibility or when spite can be successful. J Theor Biol 148: 269–277.
Hurst LD, McVean GT (1996). Clade selection, reversible evolution and the persistence of selfish elements: the evolutionary dynamics of cytoplasmic incompatibility. Proc R Soc London Ser B 263: 97–104.
Jaenike J, Dyer KA, Cornish C, Minhas MS (2006). Asymmetrical reinforcement and Wolbachia infection in Drosophila. PLoS Biol 4: e325.
Jansen VAA, Turelli M, Godfray HCJ (2008). Stochastic spread of Wolbachia. Proc R Soc London Ser B 275: 2769–2776.
Keeling MJ (1999). The effects of local spatial structure on epidemiological invasions. Proc R Soc London Ser B 266: 859–867.
Keeling MJ, Jiggins FM, Read JM (2003). The invasion and coexistence of competing Wolbachia strains. Heredity 91: 382–388.
Keller GP, Windsor DM, Saucedo JM, Werren JH (2004). Reproductive effects and geographical distributions of two Wolbachia strains infecting the Neotropical beetle, Chelymorpha alternans Boh. (Chrysomelidae, Cassidinae). Mol Ecol 13: 2405–2420.
Kobayashi Y, Hammerstein P, Telschow A (2008). The neutral effective migration rate in a mainland-island context. Theor Popul Biol 74: 84–92.
Kobayashi Y, Telschow A (2008). The concept of effective migration rate and its application in speciation models. In: Koven VT (ed). Population Genetics Research Progress. Nova Science Publishers: Hauppauge, NY. pp 151–175.
Koehnke A, Telschow A, Werren JH, Hammerstein P (2009). Life and death of an influential passenger: Wolbachia and the evolution of CI-modifiers by their hosts. PLoS One 4: e4425.
Laven H (1951). Crossing experiments with Culex strains. Evolution 5: 370–375.
Laven H (1956). Cytoplasmic inheritance in Culex. Nature 177: 141–142.
Laven H (1957). Vererbung Durch Kerngene Und Das Problem Der Ausserkaryotischen Vererbung Bei Culex pipiens. 2.Ausserkaryotische Vererbung. Zeitschrift Fur Induktive Abstammungs Und Vererbungslehre 88: 478–516.
Laven H (1959). Speciation by cytoplasmic isolation in the Culex pipiens-complex. Cold Spring Harb Symp Quant Biol 24: 166–173.
Laven H (1967). Eradication of Culex pipiens fatigans through cytoplasmic incompatibility. Nature 216: 383–384.
Marshall JF (1938). The British Mosquitoes. Trustees of the British Museum: London. pp 253–254.
McMeniman CJ, Lane RV, Cass BN, Fong AWC, Sidhu M, Wang YF et al. (2009). Stable introduction of a life-shortening Wolbachia infection into the mosquito Aedes aegypti. Science 323: 141–144.
Merçot H, Charlat S (2004). Wolbachia infections in Drosophila melanogaster and D. simulans: polymorphism and levels of cytoplasmic incompatibility. Genetica 120: 51–59.
Merçot H, Llorente B, Micheline J, Atlan A, Montchamp-Moreau C (1995). Variability within the Seychelles cytoplasmic incompatibility system in Drosophila simulans. Genetics 141: 1015–1023.
Mouton L, Henri H, Boulétreau M, Vavre F (2005). Multiple infections and diversity of cytoplasmic incompatibility in a haplodiploid species. Heredity 94: 187–192.
Narita S, Nomura M, Kageyama D (2007). Naturally occurring single and double infection with Wolbachia strains in the butterfly Eurema hecabe: transmission efficiencies and population density dynamics of each Wolbachia strain. FEMS Microbiol Ecol 61: 235–245.
Nigro L, Prout T (1990). Is There selection on RFLP differences in mitochondrial DNA? Genetics 125: 551–555.
Perlman SJ, Kelly SE, Hunter MS (2008). Population biology of cytoplasmic incompatibility: maintenance and spread of Cardinium symbionts in a parasitic wasp. Genetics 178: 1003–1011.
Perrot-Minnot MJ, Guo LR, Werren JH (1996). Single and double infections with Wolbachia in the parasitic wasp Nasonia vitripennis: effects on compatibility. Genetics 143: 961–972.
Poinsot D, Charlat S, Merçot H (2003). On the mechanism of Wolbachia-induced cytoplasmic incompatibility: confronting the models with the facts. Bioessays 25: 259–265.
Poinsot D, Montchamp-Moreau C, Merçot H (2000). Wolbachia segregation rate in Drosophila simulans naturally bi-infected cytoplasmic lineages. Heredity 82: 191–198.
Prout T (1994). Some evolutionary possibilities for a microbe that causes incompatibility in its host. Evolution 48: 909–911.
Rasgon JL, Scott TW (2003). Wolbachia and cytoplasmic incompatibility in the California Culex pipiens mosquito species complex: parameter estimates and infection dynamics in natural populations. Genetics 165: 2029–2038.
Rasgon JL, Scott TW (2004). Impact of population age structure on Wolbachia transgene driver efficacy: ecologically complex factors and release of genetically modified mosquitoes. Insect Biochem Mol Biol 34: 707–713.
Reuter M, Lehmann L, Guillaume F (2008). The spread of incompatibility-inducing parasites in sub-divided host populations. BMC Evol Biol 8: 134.
Reynolds KT, Hoffmann AA (2002). Male age, host effects and the weak expression or nonexpression of cytoplasmic incompatibility in Drosophila strains infected by maternally transmitted Wolbachia. Genet Res 80: 79–87.
Riegler M, Stauffer C (2002). Wolbachia infections and superinfections in cytoplasmically incompatible populations of the European cherry fruit fly Rhagoletis cerasi (Diptera, Tephritidae). Mol Ecol 11: 2425–2434.
Rigaud T, Rousset F (1996). What generates the diversity of Wolbachia-arthropod interactions? Biodivers Conserv 5: 999–1013.
Roubaud E (1941). Phenomena of amixia in the intercrossings of Culicids of the pipiens group. CR Hebd Seances Acad Sci 212: 257–259.
Rousset F, Raymond CS, Kjellberg F (1991). Cytoplasmic incompatibilities in the mosquito Culex pipiens: how to explain a cytotype polymorphism? J Evol Biol 4: 69–81.
Rousset F, Raymond M (1991). Cytoplasmic incompatibility in insects—why sterilize females. Trends Ecol Evol 6: 54–57.
Schofield P (2002). Spatially explicit models of Turelli-Hoffmann Wolbachia invasive wave fronts. J Theor Biol 215: 121–131.
Sinkins SP, Curtis CF, O'Neill SL (1997). The potential application of inherited symbiont systems to pest control. In: O'Neill SL, Hoffmann AA, Werren JH (eds). Influential Passengers. Oxford University Press: Oxford. pp 155–177.
Sinkins SP, Godfray HCJ (2004). Use of Wolbachia to drive nuclear transgenes through insect populations. Proc R Soc London Ser B 271: 1421–1426.
Sinkins SP, Gould F (2006). Gene drive systems for insect disease vectors. Nat Rev Genet 7: 427–435.
Telschow A, Flor M, Kobayashi Y, Hammerstein P, Werren JH (2007). Wolbachia-induced unidirectional cytoplasmic incompatibility and speciation: mainland-island model. PLoS One 2: e701.
Telschow A, Hammerstein P, Werren JH (2002a). Effects of Wolbachia on genetic divergence between populations: mainland-island model. Integr Comp Biol 42: 340–351.
Telschow A, Hammerstein P, Werren JH (2002b). The effect of Wolbachia on genetic divergence between populations: models with two-way migration. Am Nat 160: S54–S66.
Telschow A, Hammerstein P, Werren JH (2005a). The effect of Wolbachia versus genetic incompatibilities on reinforcement and speciation. Evolution 59: 1607–1619.
Telschow A, Yamamura N, Werren JH (2005b). Bidirectional cytoplasmic incompatibility and the stable coexistence of two Wolbachia strains in parapatric host populations. J Theor Biol 235: 265–274.
Tram U, Ferree PA, Sullivan W (2003). Identification of Wolbachia--host interacting factors through cytological analysis. Microbes Infect 5: 999–1011.
Tram U, Sullivan W (2002). Role of delayed nuclear envelope breakdown and mitosis in Wolbachia-induced cytoplasmic incompatibility. Science 296: 1124–1126.
Turelli M (1994). Evolution of incompatibility-inducing microbes and their hosts. Evolution 48: 1500–1513.
Turelli M, Hoffmann AA (1991). Rapid spread of an inherited incompatibility factor in California Drosophila. Nature 353: 440–442.
Turelli M, Hoffmann AA (1999). Microbe-induced cytoplasmic incompatibility as a mechanism for introducing transgenes into arthropod populations. Insect Mol Biol 8: 243–255.
Turelli M, Hoffmann AA, McKechnie SW (1992). Dynamics of cytoplasmic incompatibility and mtDNA variation in natural Drosophila simulans populations. Genetics 132: 713–723.
Vavre F, Dedeine F, Quillon M, Fouillet P, Fleury F, Boulétreau M (2001). Within-species diversity of Wolbachia-induced cytoplasmic incompatibility in haplodiploid insects. Evolution 55: 1710–1714.
Vavre F, Fleury F, Varaldi J, Fouillet P, Boulétreau M (2000). Evidence for female mortality in Wolbachia-Mediated cytoplasmic incompatibility in haplodiploid insects: epidemiologic and evolutionary consequences. Evolution 54: 191–200.
Wade MJ (2001). Infectious speciation. Nature 409: 675–677.
Wade MJ, Stevens L (1994). The effect of population subdivision on the rate of spread of parasite-mediated cytoplasmic incompatibility. J Theor Biol 167: 81–87.
Weeks AR, Turelli M, Harcombe WR, Reynolds KT, Hoffmann AA (2007). From parasite to mutualist: rapid evolution of Wolbachia in natural populations of Drosophila. PLoS Biol 5: e114.
Werren JH (1997). Biology of Wolbachia. Annu Rev Entomol 42: 587–609.
Werren JH (1998). Wolbachia and speciation. In: Howard DJ, Berlocher SH (eds). Endless Forms: Species and Speciation. Oxford University Press: New York. pp 245–260.
Xi ZY, Khoo CCH, Dobson SL (2005). Wolbachia establishment and invasion in an Aedes aegypti laboratory population. Science 310: 326–328.
Yen JH, Barr AR (1971). New hypothesis of the cause of cytoplasmic incompatibility in Culex pipiens L. Nature 232: 657–658.
Yen JH, Barr AR (1973). The etiological agent of cytoplasmic incompatibility in Culex pipiens. J Invertebr Pathol 22: 242–250.
Zabalou S, Riegler M, Theodorakopoulou M, Stauffer C, Savakis C, Bourtzis K (2004). Wolbachia-induced cytoplasmic incompatibility as a means for insect pest population control. Proc Natl Acad Sci USA 101: 15042–15045.
Acknowledgements
We thank Sylvain Charlat, Matthias Flor, Yutaka Kobayashi, Max Reuter and two anonymous reviewers for helpful comments on the article. AT acknowledges support from a postdoctoral fellowship awarded by the Volkswagen Foundation.
Author information
Authors and Affiliations
Corresponding author
Additional information
We dedicate this article to Peter Hammerstein whose enthusiasm for Wolbachia proved infectious for both of us and who is celebrating his 60th birthday this year.
Rights and permissions
About this article
Cite this article
Engelstädter, J., Telschow, A. Cytoplasmic incompatibility and host population structure. Heredity 103, 196–207 (2009). https://doi.org/10.1038/hdy.2009.53
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/hdy.2009.53
Keywords
This article is cited by
-
Environmental Temperature, but Not Male Age, Affects Wolbachia and Prophage WO Thereby Modulating Cytoplasmic Incompatibility in the Parasitoid Wasp, Habrobracon Hebetor
Microbial Ecology (2022)
-
Influence of reproductive biology on establishment capacity in introduced Hymenoptera species
Biological Invasions (2021)
-
Dual effects of insect fecundity overdispersion on the Wolbachia establishment and the implications for epidemic biocontrol
Journal of Pest Science (2021)
-
Endosymbiont diversity in natural populations of Tetranychus mites is rapidly lost under laboratory conditions
Heredity (2020)
-
Endosymbiotic Bacteria Are Prevalent and Diverse in Agricultural Spiders
Microbial Ecology (2020)
