- NEWS AND VIEWS
Rethinking WNT signalling
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 557, 495-496 (2018)
doi: https://doi.org/10.1038/d41586-018-04820-y
References
Szenker-Ravi, E. et al. Nature 557, 564–569 (2018).
Chen, P.-H., Chen, X., Lin, Z., Fang, D. & He, X. Genes Dev. 27, 1345–1350 (2013).
Gong, X. et al. PLoS One 7, e37137 (2012).
Hao, H.-X. et al. Nature 485, 195–200 (2012).
Carmon, K. S., Gong, X., Lin, Q., Thomas, A. & Liu, Q. Proc. Natl Acad. Sci. USA 108, 11452–11457 (2011).
de Lau, W. et al. Nature 476, 293–297 (2011).
Sato, T. et al. Nature 459, 262–265 (2009).
Jaks, V. et al. Nat. Genet. 40, 1291–1299 (2008).
Bell, S. M. et al. Development 135, 1049–1058 (2008).
Lebensohn, A. M. & Rohatgi, R. elife 7, e33126 (2018).
Kawakami, Y. et al. Cell 104, 891–900 (2001).
ten Berge, D., Brugmann, S. A., Helms, J. A. & Nusse, R. Development 135, 3247–3257 (2008).
Kengaku, M. et al. Science 280, 1274–1277 (1998).
Han, T. et al. Nat. Commun. 8, 15945 (2017).
 read the paper: RSPO2 inhibition of RNF43 and ZNRF3 governs limb development independently of LGR4/5/6
read the paper: RSPO2 inhibition of RNF43 and ZNRF3 governs limb development independently of LGR4/5/6
 Disarming Wnt
Disarming Wnt
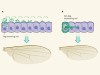 Tethered wings
Tethered wings