
Abstract
Aim:
Widespread and repeated use of azoles, particularly fluconazole, has led to the rapid development of azole resistance in Candida albicans. We investigated the role of CaIPF14030 during the development of azole resistance in C albicans.
Methods:
The expression of CaIPF14030 was measured by quantitative RT-PCR, and CaIPF14030 was disrupted by the hisG-URA3-hisG (URA-blaster) method. The sensitivity of C albicans to azoles was examined using a spot assay, and the intracellular ATP concentrations were measured by a luminometer.
Results:
CaIPF14030 expression in C albicans was up-regulated by Ca2+ in a calcineurin-dependent manner, and the protein was overexpressed during the stepwise acquisition of azole resistance. However, disruption or ectopic overexpression of CaIPF14030 did not affect the sensitivity of C albicans to azoles. Finally, we demonstrated that disruption of CaIPF14030 significantly increased intracellular ATP levels, and overexpression significantly decreased intracellular ATP levels in C albicans.
Conclusion:
CaIPF14030 may negatively modulate intracellular ATP levels during the development of azole resistance in C albicans.
Similar content being viewed by others

Introduction
Candida albicans (C albicans), a major human fungal pathogen, causes disorders ranging from mild infections to life-threatening diseases1, 2. C albicans is often treated with fluconazole, which is a fungistatic drug. However, there is a rapid acquisition of resistance to azoles in C albicans. This is due to a limited number of multidrug resistance related genes, which include ERG11, CDR1, CDR2, and CaMDR13, 4, 5, 6. Recently, DNA microarray and proteomic analyses have revealed many differentially regulated genes in either laboratory7, 8 or clinical9, 10, 11, 12 azole-resistant C albicans isolates. However, few of these new observations have been confirmed by genetic studies because C albicans is difficult to study experimentally due to its asexual diploid nature and variant genetic code.
The calcineurin pathway is involved in the development of azole resistance in C albicans13, 14, 15. Calcineurin is a Ca2+ calmodulin-dependent serine/threonine phosphatase consisting of a catalytic subunit A (encoded by CNA1) and a regulatory subunit B (encoded by CNB1)16. The phosphatase activity of calcineurin is activated when calcineurin binds calmodulin in the presence of calcium ions. Activated calcineurin regulates downstream gene expression via transcription factors such as Crz1p17.
In our previous study, we found that RTA2, CaIPF14030, and MXR1 were involved in the calcineurin pathway. These three genes were overexpressed in the experimentally induced azole-resistant C albicans strain DSF2818. The rta2 and ipf14030 null mutants from C albicans were constructed along with deletions of CDR1, CDR2, and CaMDR118. The disruption of RTA2 increased the susceptibility of C albicans to azoles, whereas the disruption of CaIPF14030 did not influence the sensitivity of C albicans to azoles18. Bioinformatic analysis also revealed that the promoters of both the RTA2 and the CaIPF14030 genes contained the calcineurin-dependent responsive element (CDRE) sequence, which is controlled by either calcineurin or Crz1p17, 18. However, the function of CaIPF14030 was unknown. In the present study, we constructed the ipf14030 null mutant from the C albicans wild-type strain to investigate the role of CaIPF14030 during the development of azole resistance in C albicans.
Materials and methods
Antifungal agents
Fluconazole was from Pfizer Inc (New York, NY, USA). Ketoconazole and econazole were purchased from Sigma (St Louis, MO, USA). Itraconazole was a gift from Prof Chao-mei LIU, and voriconazole was a gift from Prof Wan-nian ZHANG from the Second Military Medical University, Shanghai, China.
Candida albicans strains and culture media
Candida albicans strains used in this study are listed in Table 1; they were cultured in YPD medium or SC medium lacking uridine, methionine and cysteine.
RNA isolation and quantitative RT-PCR
All primer sequences are listed in Table 2. RNA isolation, cDNA synthesis, and PCR amplification were performed as described previously18. Triplicate independent quantitative RT-PCRs were performed using the LightCycler System (Roche Diagnostics, Mannheim, Germany). Gene expression levels relative to the calibrator were expressed as 2−ΔΔCT.
Construction of mutant strains
All primer sequences are listed in Table 2. Construction of plasmid pUC-IPF14030-URA3 was performed as described previously18. Briefly, the fragment containing the 5′ and 3′ ends of the CaIPF14030 gene for homologous recombination was obtained according to the Fusion PCR method. The 4-kb hisG-URA3-hisG fragment was from the p5921 plasmid19. The two fragments were subsequently cloned into plasmid pUCm-t (Sangon, Shanghai, China). The XhoI-digested fragment of pUC-IPF14030-URA3 was transformed into the ura3 mutant strain (RM1000) by standard methods20. The hybridization probe primers are listed in Table 2. Southern blot analysis was used to confirm the absence of the CaIPF14030 gene.
Overexpression of CaIPF14030 in ipf14030 mutants
The CaIPF14030 ORF was amplified by PCR with Pyrobest polymerase (TaKaRa, Dalian, China). The BamH I/Pst I-digested PCR fragment was ligated into pCaExp21 to obtain recombinant plasmid pEXP-IPF14030. DNA sequencing confirmed that the sequence of the insert was identical to the CaIPF14030 sequence reported in the Candida Genome Database (http://www.candidagenome.org/). The ipf14030 mutant (MZP101) was transformed with the Stu I-digested and linearized pEXP-IPF14030 plasmid and selected on SC medium lacking uridine, methionine and cysteine.
Susceptibility testing
The sensitivities of the mutant strains to azoles were determined by testing the strains on agar plates containing different concentrations of azoles (fluconazole, ketoconazole, econazole, itraconazole, and voriconazole). Five microliters of ten-fold serial dilutions of each yeast culture (OD600=1.0) were spotted on plates of the appropriate medium and then incubated at 30 °C for the indicated time.
Measurement of intracellular ATP levels
C albicans strains were diluted to 1×108 or 1×107 cells/mL in YPD broth. A total of 100 μL of cell suspension was mixed completely with the same volume of BacTiter-Glo reagent (Promega Corporation, Madison, WI, USA) and incubated for 10 min at room temperature. Luminescent signals were measured with a TD 20/20 luminometer (Turner Biosystem, Sunnyvale, CA, USA) with a 1 s integration time per sample. A control tube without cells was used to obtain a value for background luminescence. The signal-to-noise ratio (S/N) was calculated as follows: [mean of signal - mean of background]/standard deviation of background. A standard curve for ATP increments from 10 pmol/L to 1 μmol/L was constructed. Signals represented the mean of three separate experiments. The ATP content was calculated from the standard curve.
Statistical analysis
Experiments were performed at least three times. Data are presented as mean±standard deviations, and data were analyzed using Student's t test where indicated.
Results
CaIPF14030 up-regulation by Ca2+ in a calcineurin-dependent manner
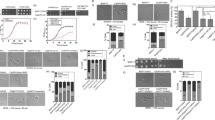
Bioinformatic analysis revealed that the CaIPF14030 promoter contains the CDRE sequence, which can be controlled by either calcineurin or Crz1p17. In our study, expression levels of CaIPF14030 were examined by quantitative RT-PCR in wild-type (CAF2-1), cnaΔ/Δ mutant (DSY2091), crz1Δ/Δ mutant (DSY2195), CNA revertant (DSY2115) and CRZ1 revertant (MKY268) strains after exposure to 200 mmol/L CaCl2 for 2 h (Figure 1A). CaIPF14030 was up-regulated by 6.51 fold in the CAF2-1 strain (Figure 1A). Depletion of CNA or CRZ1 inhibited the effects of CaCl2 on CaIPF14030 up-regulation by 1.53 fold in the DSY2091 strain and by 0.96 fold in the DSY2195 strain (Figure 1A). However, reintroduction of CNA or CRZ1 in the respective null mutant strains restored the effect of 200 mmol/L CaCl2 on CaIPF14030 up-regulation. There was a 3.62-fold change in the DSY2115 strain and a 3.19-fold change in the MKY268 strain (Figure 1A). Taken together, these data demonstrate that CaIPF14030 is up-regulated by Ca2+ in a calcineurin-dependent manner.

(A) Expression of CaIPF14030 was examined by quantitative RT-PCR in wild-type (CAF2-1), cnaΔ/Δ mutant (DSY2091), CNA revertant (DSY2115), crz1Δ/Δ mutant (DSY2195), and CRZ1 revertant (MKY268) strains in the presence of 200 mmol/L CaCl2. The corresponding strains in the absence of CaCl2 were used as controls. (B) Expression of CaIPF14030 was examined by quantitative RT-PCR in DSF7, DSF14, DSF21, and DSF28 strains obtained in our previous study18 and compared to DSY1024. (C) Expression of CaIPF14030 in the Exp-14030 strain (MZP101 carrying pCaEXP-IPF14030) was compared to the Exp-RM strain (RM1000 carrying pCaEXP). Strains were cultured in SC medium lacking methionine, cysteine, and uridine for 16 h before harvesting for quantitative RT-PCR analysis. Data are represented as mean±SD.
CaIPF14030 is overexpressed during the stepwise acquisition of azole resistance in DSY1024
It has been well documented that calcium can activate the calcineurin pathway and modulate azole activity in C albicans22, 23. Because the CaIPF14030 gene of C albicans is up-regulated by Ca2+ in a calcineurin-dependent manner, we investigated the dynamic expression of CaIPF14030 in the DSF7, DSF14, DSF21, and DSF28 strains obtained in our previous study18. As shown in Figure 1B, the CaIPF14030 gene was overexpressed in the DSF7, DSF14, DSF21, and DSF28 strains. These results are similar to those found for other known resistance-related genes such as ERG11, CDR1, and CaMDR14, 24. These data suggest that CaIPF14030 is involved in the development of azole resistance in C albicans.
Disruption and ectopic overexpression of the CaIPF14030 gene in C albicans
To investigate the role of CaIPF14030 during the development of azole resistance in C albicans, two alleles of CaIPF14030 were sequentially disrupted in the RM1000 strain using the URA-blaster method and 5-FOA selection. This yielded the following strains: Ura+ ipf14030/IPF14030 (MZP10), Ura− ipf14030/IPF14030 (MZP11), Ura+ ipf14030/ipf14030 (MZP100), and Ura− ipf14030/ipf 14030 (MZP101) (Table 1). The strategy to disrupt both copies of CaIPF14030 is depicted in Figure 2A and 2B. Southern blot analysis of DNA from the constructed strains revealed that the recombination patterns were as expected (Figure 2C). To further characterize the generated mutants, RM1000 and MZP101 growth curves were obtained. They showed identical growth rates.

Schematic representation of the plasmid construction and disruption of CaIPF14030 using the URA3 selection marker. (A) Diagrammatic representation of the plasmid construction of pUC-IPF14030-URA3 containing hisG-URA3-hisG with flanking portions on either side for homologous recombination. Ba: BamH I, X: Xho I, N: Nde I. (B) Sequential targeted disruption of the two CaIPF14030 alleles in C albicans with the disruption cassette. Strain designation is shown on the right. (C) Southern blot analysis of the genomic DNA digested with Kpn I and Sac I. The exact size and genotype of the expected hybridizing DNA fragment are indicated on the right. The hybridization probe was a 0.528-kb PCR fragment from −432 to 96 of the CaIPF14030 gene amplified by Pyrobest polymerase (TaKaRa, Dalian, China) from genomic DNA extracted from RM1000.
To investigate whether other deletions were introduced into the C albicans genome, the CaIPF14030 ORF was placed under the control of the MET3 promoter in the pCaEXP vector, and it was successfully integrated into the ipf14030Δ/Δ mutant (MZP101) at the RP10 locus as determined by PCR (data not shown). Ectopic overexpression of CaIPF14030 in the Exp-14030 strain was confirmed by quantitative RT-PCR with CaIPF14030 expression in the parental strain (RM1000) carrying empty vector pCaEXP, which served as a control (Figure 1C). The growth rates of the RM1000 and Exp-14030 strains were similar, suggesting that there were no other gene disruptions that affected the growth of the strains.
Disruption or ectopic overexpression of CaIPF14030 does not affect the sensitivity of C albicans to azoles
Because the CaIPF14030 gene was overexpressed during the stepwise acquisition of azole resistance in DSY1024, a drug susceptibility assay was performed to determine whether CaIPF14030 is associated with enhanced azole resistance in C albicans. Spot assay results demonstrated that neither disruption nor ectopic overexpression of CaIPF14030 affected the sensitivity of C albicans to the azoles, including fluconazole, ketoconazole, econazole, itraconazole, and voriconazole (Figure 3A, 3B). Taken together, these data support the hypothesis that CaIPF14030 does not play an important role in the development of azole resistance in C albicans.

Drug susceptibility profiles of C albicans strains (Table 1) determined by spot assays. (A) The ipf14030 heterozygous and homozygous mutant strains from RM1000 were spotted on YPD agar plates supplemented with 0.0025% uridine with or without different antifungal agents at the indicated concentrations. Plates were incubated for 48 h at 30°C. (B) The ipf14030 mutant strains (MZP101) carrying pCaEXP or pCaEXP-IPF14030 and the parental strain (RM1000) carrying pCaEXP were spotted on SC medium plates lacking methionine, cysteine, and uridine with or without drugs. Plates were incubated for 72 h at 30 °C.
CaIPF14030 negatively controls the intracellular ATP content of C albicans
It has been well documented that lower intracellular ATP levels are found in C albicans strains that are azole resistant or have been treated by azoles25, 26. Because overexpression of CaIPF14030 occurred during the development of azole resistance in C albicans, intracellular ATP concentrations were measured in the following strains: wild-type (RM1000), ipf14030Δ/Δ mutant (MZP101) and ectopically over-expressed (Exp-14030). As shown in Figure 4, the level of intracellular ATP in the MZP101 strain was significantly higher than that in the RM1000 strain (P<0.01), whereas the level of intracellular ATP in the Exp-14030 strain was significantly lower than that in the RM1000 strain (P<0.01). These results suggest that CaIPF14030 plays a critical role in the generation of intracellular ATP during the development of azole resistance in C albicans.

Intracellular ATP content in the wild-type (RM1000), RM1000 carrying pCaEXP (Exp-RM), ipf14030 mutant (MZP101) and MZP101 carrying pCaEXP-IPF14030 (Exp-14030) strains. ATP levels represent the mean±standard deviation for three independent experiments. bP<0.05 when compared to the control strains (RM1000 or Exp-RM); cP<0.01 when compared to the control strains (RM1000 or Exp-RM).
Discussion
Bioinformatic analysis have revealed that the CaIPF14030 promoter contains a CDRE sequence, which can be controlled by either calcineurin or Crz1p17. The expression levels of CaIPF14030 were examined by quantitative RT-PCR in the wild-type (CAF2-1), cnaΔ/Δ mutant (DSY2091), crz1Δ/Δ mutant (DSY2195), CNA revertant (DSY2115) and CRZ1 revertant (MKY268) strains after exposure to 200 mmol/L CaCl2. CaIPF14030 was up-regulated by Ca2+ in a calcineurin-dependent manner. It is well documented that calcium can activate the calcineurin pathway and modulate azole activity in C albicans22, 23. In addition, the dynamic expression of CaIPF14030 was investigated in the DSF7, DSF14, DSF21, and DSF28 strains obtained in our previous study 18. The CaIPF14030 gene shows a steady overexpression state in the DSF7, DSF14, DSF21, and DSF28 strains. These results are similar to those involving other known resistance-related genes such as ERG11, CDR1, and CaMDR14, 24. Taken together, these data suggest that CaIPF14030 is involved in the development of azole resistance in C albicans.
Both copies of CaIPF14030 were successfully disrupted, and the role of CaIPF14030 during the development of azole resistance was investigated. A drug susceptibility assay demonstrated that the disruption or ectopic overexpression of CaIPF14030 did not affect the sensitivity of the tested C albicans strains to azoles. Therefore, these data support the hypothesis that CaIPF14030 does not play an important role in the development of azole resistance in C albicans. Recent studies have shown that lower intracellular ATP levels were observed in C albicans strains that were azole-resistant or had been treated with azoles25, 26. Our data showed that disruption of CaIPF14030 significantly increased intracellular ATP content, whereas ectopic overexpression of CaIPF14030 significantly decreased intracellular ATP content. Therefore, CaIPF14030 may play a critical role in regulating intracellular ATP levels during the development of azole resistance in C albicans. Further analysis is required to elucidate the precise relationship between the generation of intracellular ATP that is negatively controlled by CaIPF14030 and the lower intracellular ATP levels in C albicans strains that are azole resistant or have been treated with azoles.
It has been previously reported that there was a strong correlation between azole susceptibility and intracellular ATP levels in C albicans27. It has also been documented that several peptides have synergistic activity against C albicans when combined with fluconazole due to inhibition of production and release of intracellular ATP28, 29, 30. Because CaIPF14030 plays a critical role in regulating intracellular ATP levels, the protein encoded by CaIPF14030 may be targeted by the inhibitors of intracellular ATP. Further studies must be performed to unravel the function of CaIPF14030 in C albicans.
A BLAST search of the C albicans genome (available at http://www.genolist.pasteur.fr/CandidaDB/) revealed that the CaIPF14030 gene (also known as orf19.851) encoded a putative polypeptide of 839 amino acids with a calculated molecular mass of 97.2 kDa. The SOSUI31 program predicted that the protein encoded by CaIPF14030 was an integral membrane protein with one membrane-spanning segment. There are two CaIPF14030 homologs (Mnn4p and YJR061Wp) in the S cerevisiae genome database (available at http://www.yeastgenome.org/) that have 30%–37% identity and 44%–55% similarity with the C albicans protein. Mnn4p has been shown to be a positive regulator of mannosylphosphate transferase, and it is involved in mannosylphosphorylation of N-linked oligosaccharides. Expression of MNN4 increases in late-logarithmic and stationary growth phases32, 33. It has been shown that YJR061W is a putative membrane protein involved in glycosylation, and it has been shown to be repressed by Rim101p34, 35. Further studies are needed to verify whether CaIPF14030 is involved in mannosylphosphorylation or glycosylation.
Our data show that CaIPF14030 is up-regulated by Ca2+ in a calcineurin-dependent manner as well as during the stepwise acquisition of azole resistance in C albicans. Disruption or ectopic overexpression of CaIPF14030 did not affect the sensitivity of C albicans to azoles, although disruption of CaIPF14030 significantly increased intracellular ATP content, and ectopic overexpression significantly decreased intracellular ATP content. In conclusion, our findings suggest that CaIPF14030 plays a critical role in the generation of intracellular ATP during the development of azole resistance in C albicans.
Author contribution
Xin-ming JIA and Jun GU designed the experiments; Xin-ming JIA performed the experiments and wrote the paper; and Jun-dong ZHANG, Ying WANG, Yuan-ying JIANG, and Hong-yue TAN critically read the manuscript.
References
Richardson MD . Changing patterns and trends in systemic fungal infections. J Antimicrob Chemother 2005; 56: i5–i11.
Garber G . An overview of fungal infections. Drugs 2001; 61: 1–12.
Prasad R, De Wergifosse P, Goffeau A, Balzi E . Molecular cloning and characterization of a novel gene of Candida albicans, CDR1, conferring multiple resistance to drugs and antifungals. Curr Genet 1995; 27: 320–9.
Sanglard D, Kuchler K, Ischer F, Pagani JL, Monod M, Bille J . Mechanisms of resistance to azole antifungal agents in Candida albicans isolates from AIDS patients involve specific multidrug transporters. Antimicrob Agents Chemother 1995; 39: 2378–86.
White TC . Increased mRNA levels of ERG16, CDR, and MDR1 correlate with increases in azole resistance in Candida albicans isolates from a patient infected with human immunodeficiency virus. Antimicrob Agents Chemother 1997; 41: 1482–7.
Prasad R, Kapoor K . Multidrug resistance in yeast Candida. Int Rev Cytol 2005; 242: 215–48.
Barker KS, Crisp S, Wiederhold N, Lewis RE, Bareither B, Eckstein J, et al. Genome-wide expression profiling reveals genes associated with amphotericin B and fluconazole resistance in experimentally induced antifungal resistant isolates of Candida albicans. J Antimicrob Chemother 2004; 54: 376–85.
Cowen LE, Nantel A, Whiteway MS, Thomas DY, Tessier DC, Kohn LM, et al. Population genomics of drug resistance in Candida albicans. Proc Natl Acad Sci U S A 2002; 99: 9284–9.
Rogers PD, Barker KS . Genome-wide expression profile analysis reveals coordinately regulated genes associated with stepwise acquisition of azole resistance in Candida albicans clinical isolates. Antimicrob Agents Chemother 2003; 47: 1220–7.
Xu Z, Zhang LX, Zhang JD, Cao YB, Yu YY, Wang DJ, et al. cDNA microarray analysis of differential gene expression and regulation in clinically drug-resistant isolates of Candida albicans from bone marrow transplanted patients. Int J Med Microbiol 2006; 296: 421–34.
Hooshdaran MZ, Barker KS, Hilliard GM, Kusch H, Morschhauser J, Rogers PD . Proteomic analysis of azole resistance in Candida albicans clinical isolates. Antimicrob Agents Chemother 2004; 48: 2733–5.
Kusch H, Biswas K, Schwanfelder S, Engelmann S, Rogers PD, Hecker M, et al. A proteomic approach to understanding the development of multidrug-resistant Candida albicans strains. Mol Genet Genomics 2004; 271: 554–65.
Sanglard D, Ischer F, Marchetti O, Entenza J, Bille J . Calcineurin A of Candida albicans: involvement in antifungal tolerance, cell morphogenesis and virulence. Mol Microbiol 2003; 48: 959–76.
Cannon RD, Lamping E, Holmes AR, Niimi K, Tanabe K, Niimi M, et al. Candida albicans drug resistance another way to cope with stress. Microbiology 2007; 153: 3211–7.
Akins RA . An update on antifungal targets and mechanisms of resistance in Candida albicans. Med Mycol 2005; 43: 285–318.
Aramburu J, Rao A, Klee CB . Calcineurin: from structure to function. Curr Top Cell Regul 2000; 36: 237–95.
Karababa M, Valentino E, Pardini G, Coste AT, Bille J, Sanglard D . CRZ1, a target of the calcineurin pathway in Candida albicans. Mol Microbiol 2006; 59: 1429–51.
Jia XM, Ma ZP, Jia Y, Gao PH, Zhang JD, Wang Y, et al. RTA2, a novel gene involved in azole resistance in Candida albicans. Biochem Biophys Res Commun 2008; 373: 631–6.
Fonzi WA, Irwin MY . Isogenic strain construction and gene mapping in Candida albicans. Genetics 1993; 134: 717–28.
Sanglard D, Ischer F, Monod M, Bille J . Susceptibilities of Candida albicans multidrug transporter mutants to various antifungal agents and other metabolic inhibitors. Antimicrob Agents Chemother 1996; 40: 2300–5.
Care RS, Trevethick J, Binley KM, Sudbery PE . The MET3 promoter: a new tool for Candida albicans molecular genetics. Mol Microbiol 1999; 34: 792–8.
Cruz MC, Goldstein AL, Blankenship JR, Del Poeta M, Davis D, Cardenas ME, et al. Calcineurin is essential for survival during membrane stress in Candida albicans. Embo J 2002; 21: 546–59.
Steinbach WJ, Reedy JL, Cramer RA Jr, Perfect JR, Heitman J . Harnessing calcineurin as a novel anti-infective agent against invasive fungal infections. Nat Rev Microbiol 2007; 5: 418–30.
Calvet HM, Yeaman MR, Filler SG . Reversible fluconazole resistance in Candida albicans: a potential in vitro model. Antimicrob Agents Chemother 1997; 41: 535–9.
Xu Y, Wang Y, Yan L, Liang RM, Tang RJ, Gao PH, et al. Proteomic analysis reveals a synergistic mechanism of fluconazole and berberine against fluconazole-resistant Candida albicans: endogenous ros augmentation. J Proteome Res 2009; 8: 5296–304.
Zheng C, Yan Z, Liu W, Jiang L . Identification and characterization of a functional Candida albicans homolog of the Saccharomyces cerevisiae TCO89 gene. FEMS Yeast Res 2007; 7: 558–68.
Kretschmar M, Nichterlein T, Kuntz P, Hof H . Rapid detection of susceptibility to fluconazole in Candida species by a bioluminescence assay of intracellular ATP. Diagn Microbiol Infect Dis 1996; 25: 117–21.
Lupetti A, Paulusma-Annema A, Welling MM, Dogterom-Ballering H, Brouwer C, Senesi S, et al. Synergistic activity of the N-terminal peptide of human lactoferrin and fluconazole against Candida species. Antimicrob Agents Chemother 2003; 47: 262–7.
Kuipers ME, De Vries HG, Eikelboom MC, Meijer DKF, Swart PJ . Synergistic fungistatic effects of lactoferrin in combination with antifungal drugs against clinical Candida isolates. Antimicrob Agents Chemother 1999; 43: 2635–41.
Kavanagh K, Dowd S . Histatins: antimicrobial peptides with therapeutic potential. J Pharm Pharmacol 2004; 56: 285–89.
Hirokawa T, Boon-Chieng S, Mitaku S . SOSUI: classification and secondary structure prediction system for membrane proteins. Bioinformatics 1998; 14: 378–9.
Odani T, Shimma Y, Tanaka A, Jigami Y . Cloning and analysis of the MNN4 gene required for phosphorylation of N-linked oligosaccharides in Saccharomyces cerevisiae. Glycobiology 1996; 6: 805–10.
Odani T, Shimma Y, Wang XH, Jigami Y . Mannosylphosphate transfer to cell wall mannan is regulated by the transcriptional level of the MNN4 gene in Saccharomyces cerevisiae. FEBS Lett 1997; 420: 186–90.
Lamb TM, Mitchell AP . The transcription factor Rim101p governs ion tolerance and cell differentiation by direct repression of the regulatory genes NRG1 and SMP1 in Saccharomyces cerevisiae. Mol Cell Biol 2003; 23: 677–86.
Conde R, Pablo G, Cueva R, Larriba G . Screening for new yeast mutants affected in mannosylphosphorylation of cell wall mannoproteins. Yeast 2003; 20: 1189–211.
Acknowledgements
We are grateful to Prof D SANGLARD for the gift of the CAF2-1, DSY2091, DSY2115, DSY2195, MKY268, and DSY1024 strains and to W A FONZI for plasmid p5921. We also thank J PLA for kindly providing the C albicans strain RM1000 and P E SUDBERY for plasmid pCaExp. This work was supported by the National Natural Science Foundation of China (No 30900055, 30872276, and 30972650).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Jia, Xm., Wang, Y., Zhang, Jd. et al. CaIPF14030 negatively modulates intracellular ATP levels during the development of azole resistance in Candida albicans. Acta Pharmacol Sin 32, 512–518 (2011). https://doi.org/10.1038/aps.2010.232
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/aps.2010.232
